

Anti-Allergic Effect of Dietary Polyphenols Curcumin and Epigallocatechin Gallate via Anti-Degranulation in IgE/Antigen-Stimulated Mast Cell Model: A Lipidomics Perspective

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
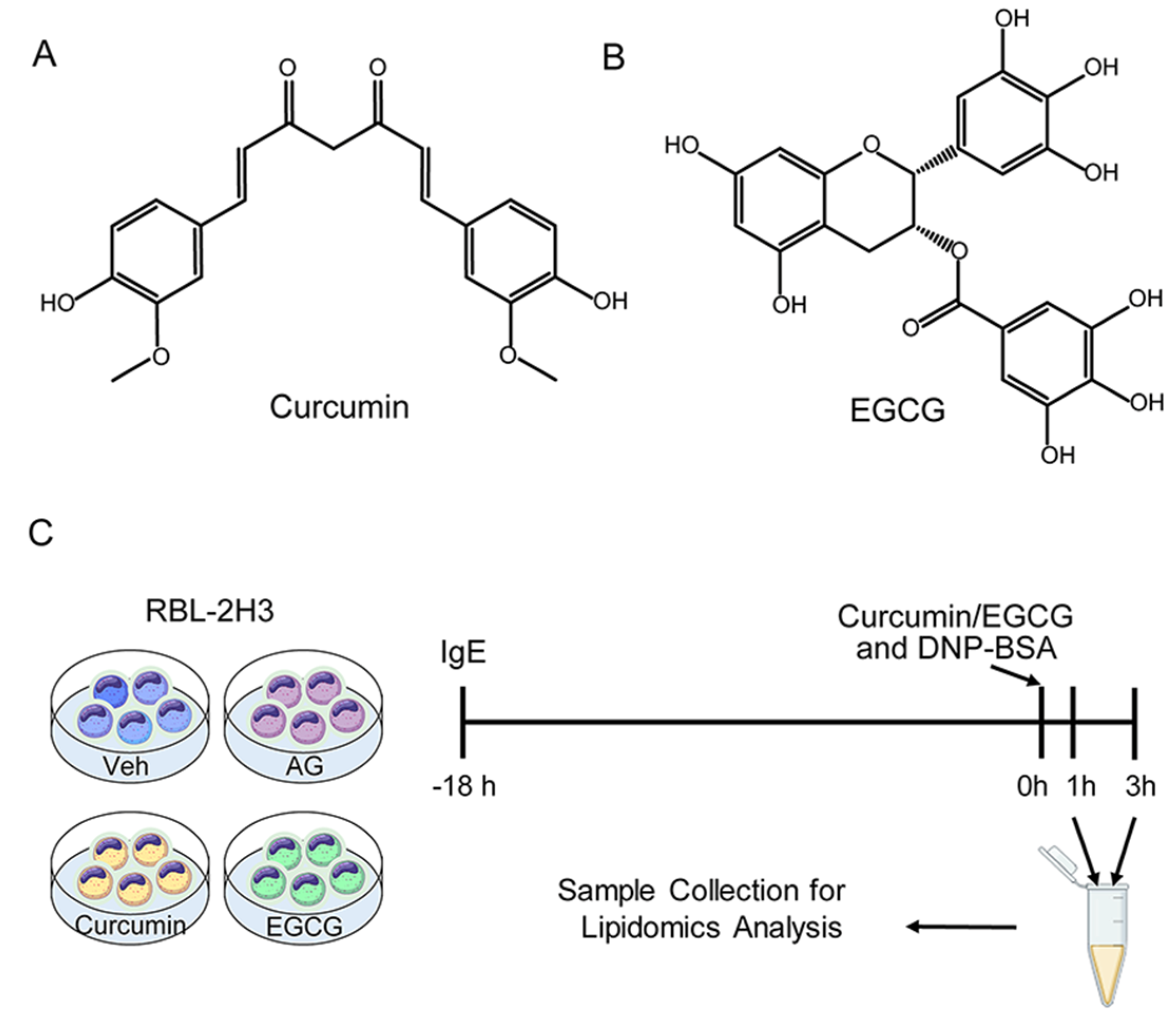
2.1. Cell Culture and Cell Viability
2.2. Sample Collection
2.3. Nontargeted Lipidomics Study
2.4. Data Processing and Statistics
2.5. β-Hexosaminidase Release Assay
2.6. TNF-α and IL-4 Release Assay
3. Results
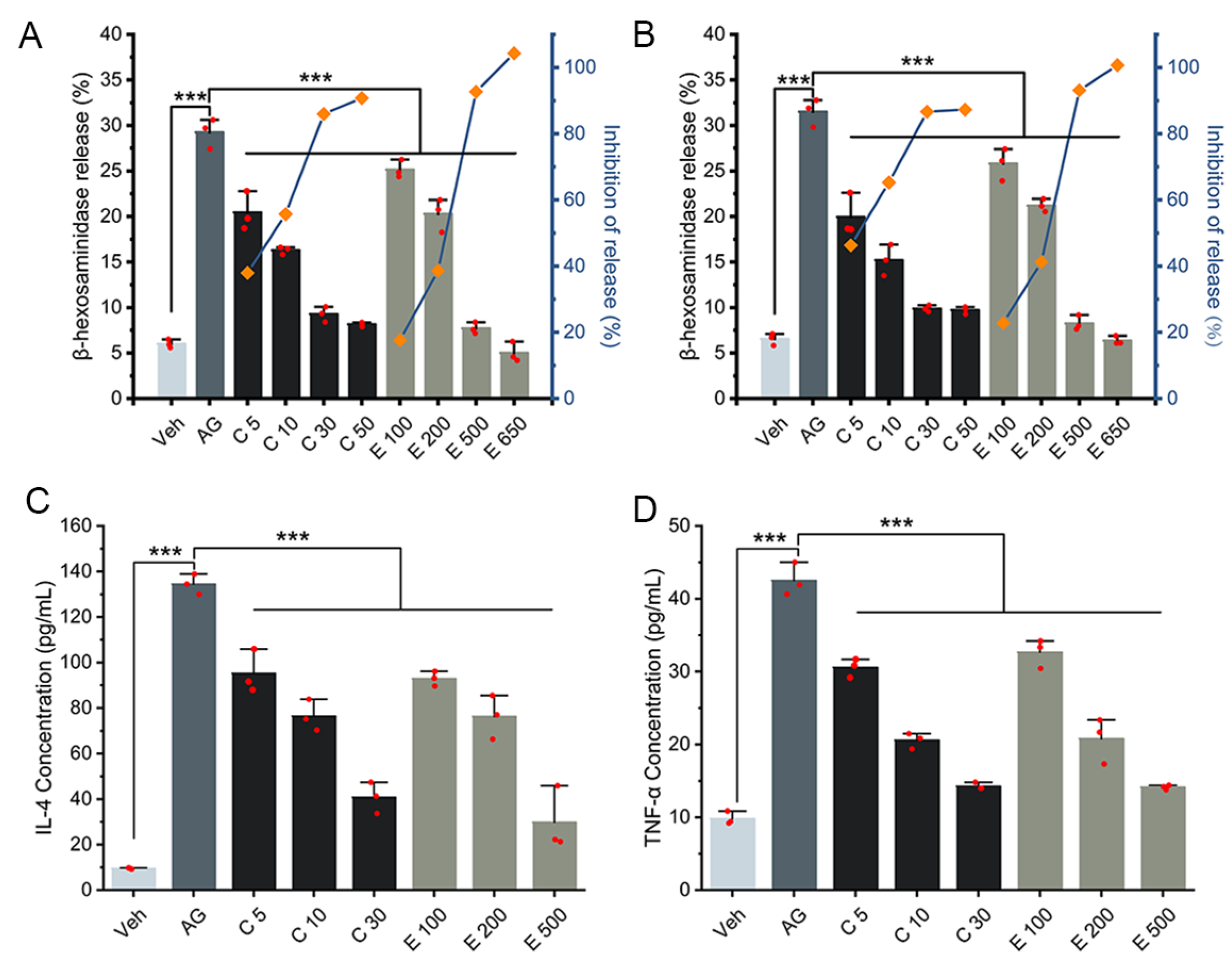
3.1. Inhibitory Effect of Curcumin/EGCG on IgE-Mediated Degranulation
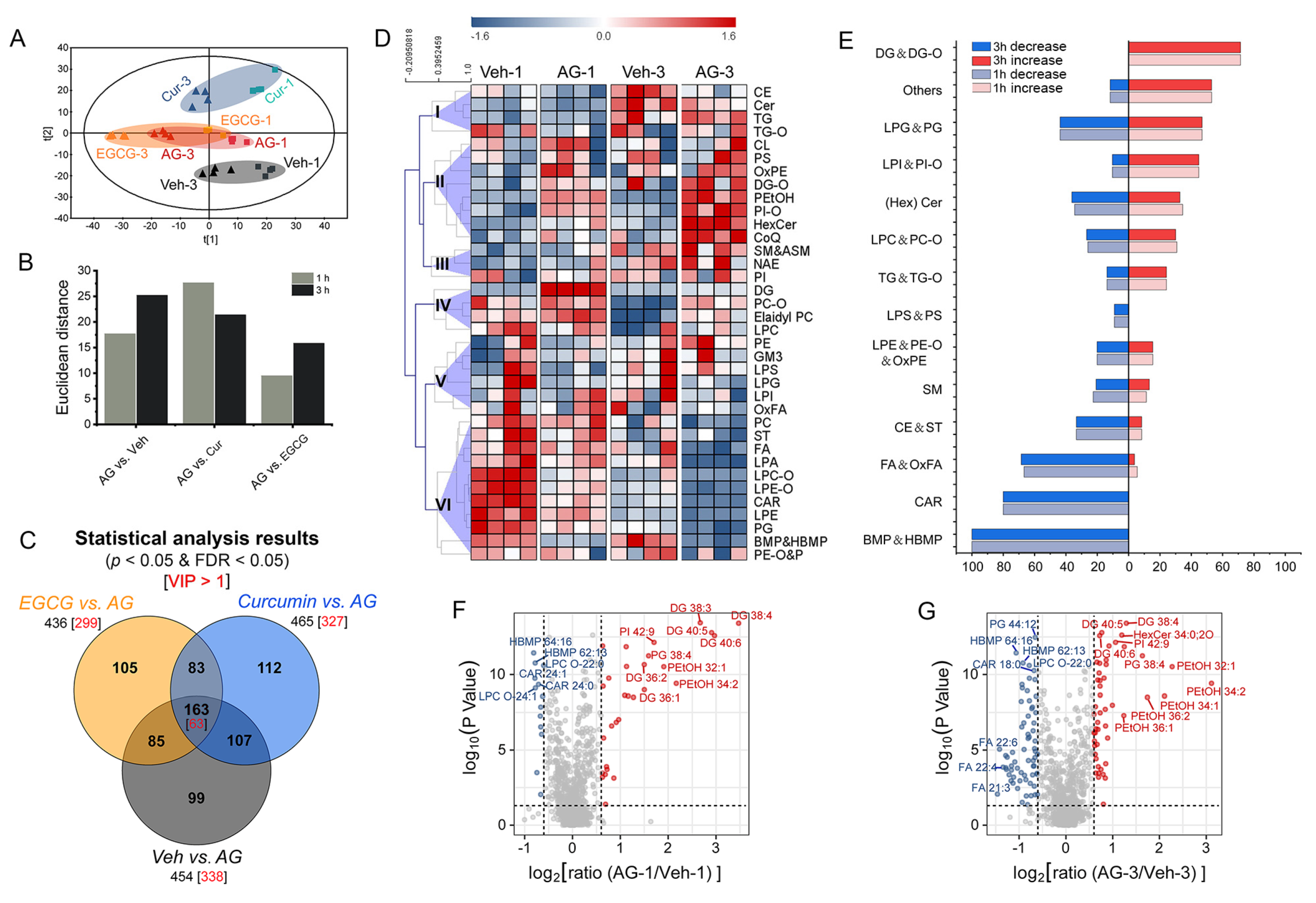
3.2. Lipidome of RBL-2H3 Cells
3.3. Global Profiling of Lipidome Disturbance
3.4. Lipidome Changes Associated with IgE-Mediated Degranulation
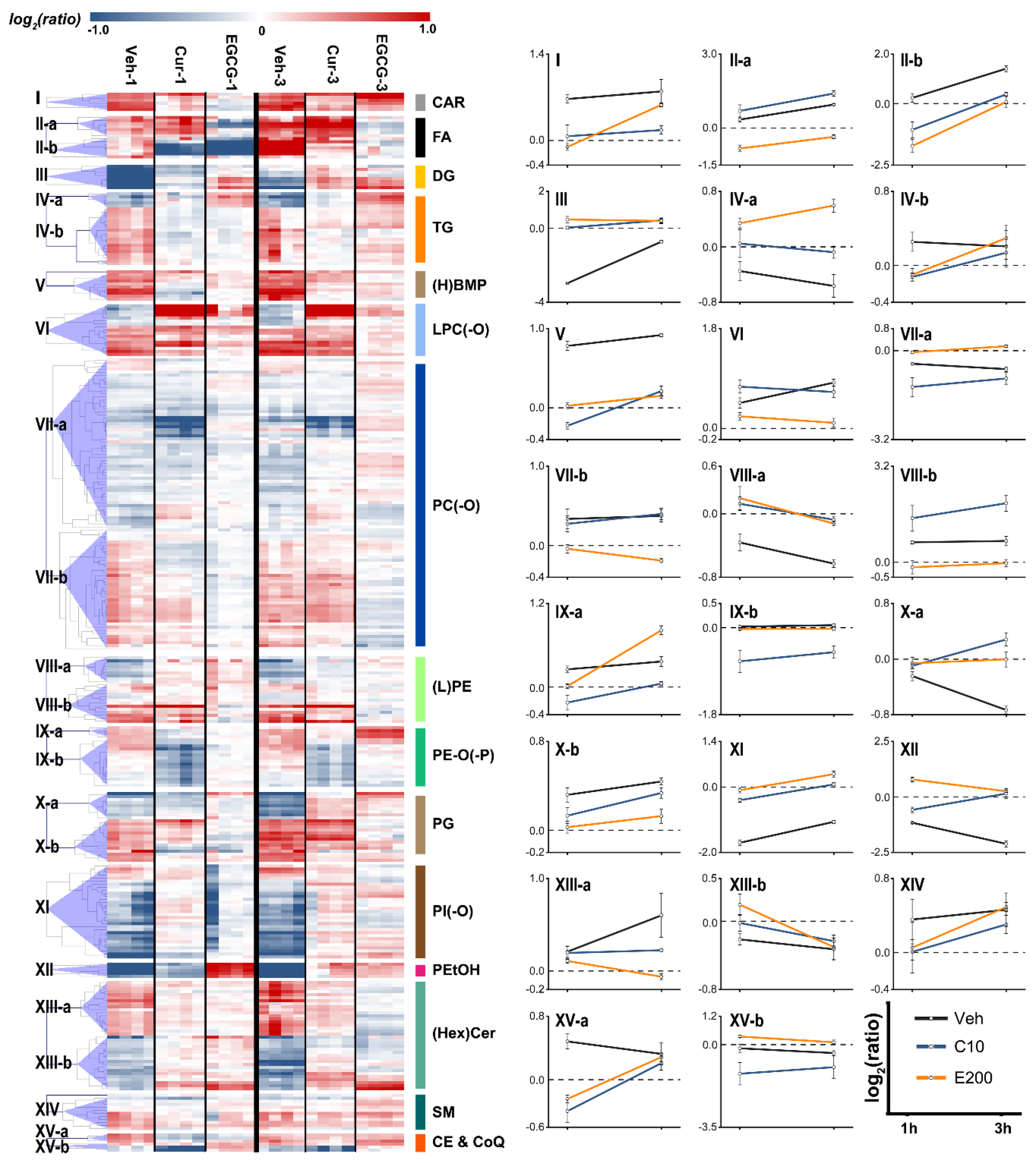
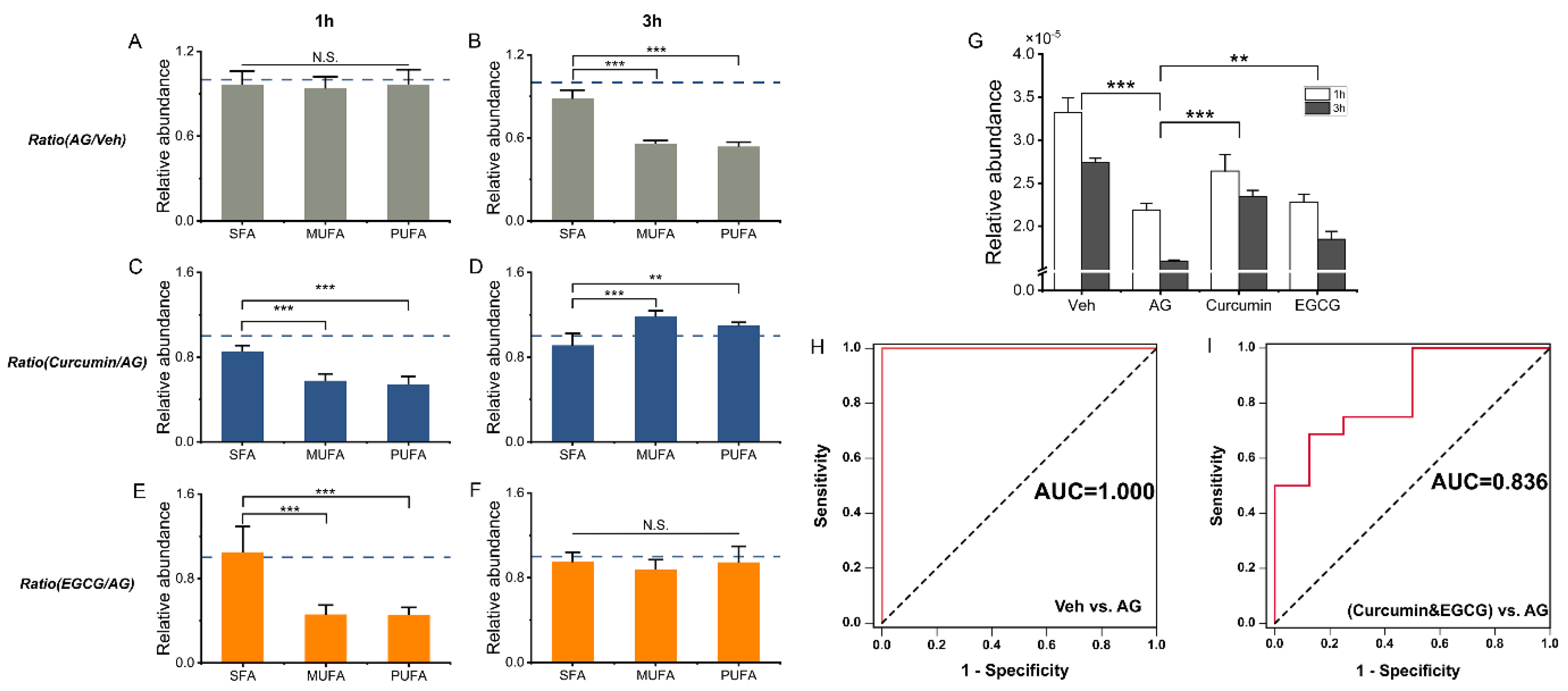
3.5. Comparison between Curcumin and EGCG Intervention
3.6. Defining Potential Biomarkers
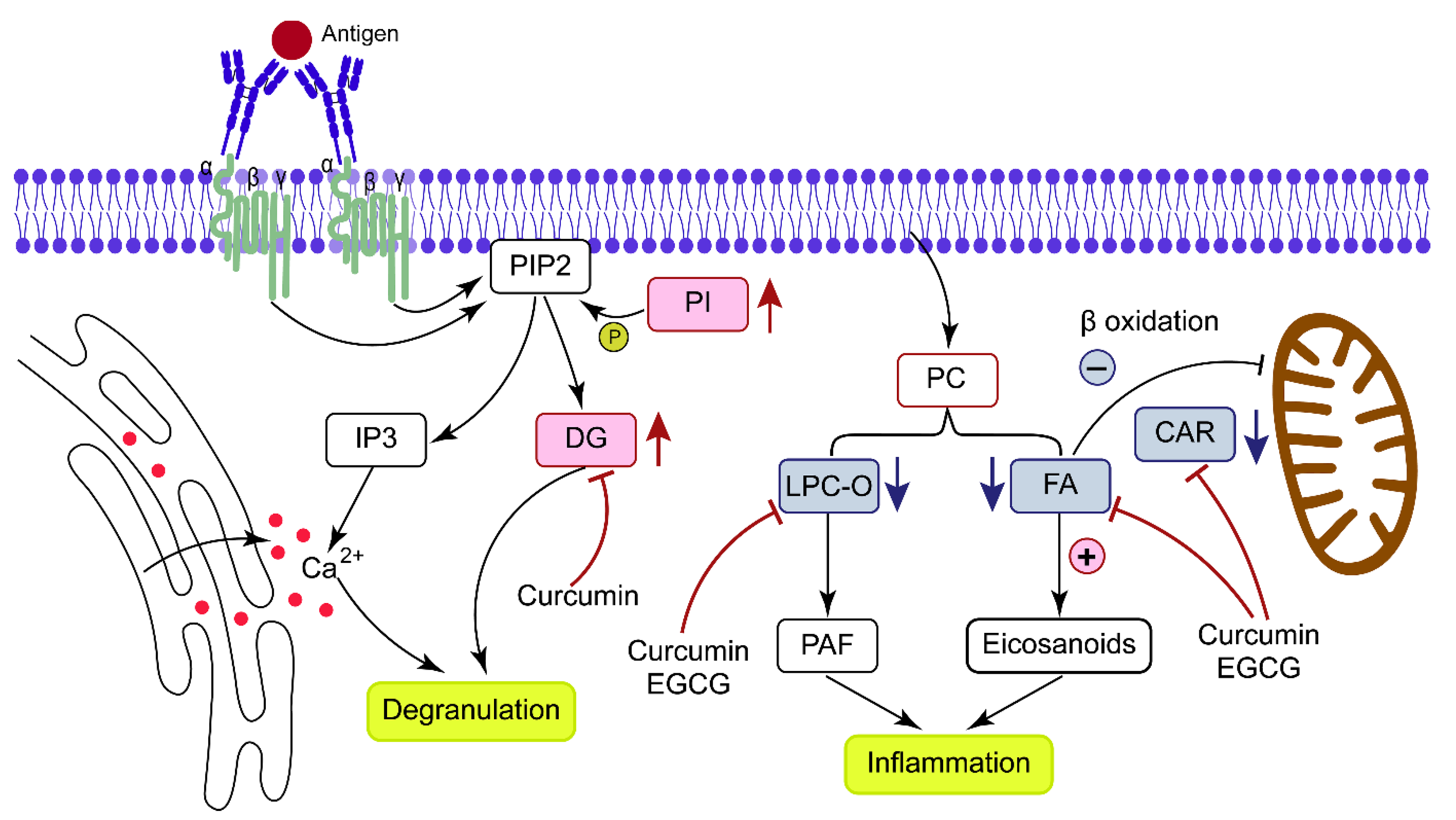
4. Discussion
4.1. DG Metabolism
4.2. FA Metabolism
4.3. BMP Metabolism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Qu, Y.Z.; Gao, Y.R.; Sun, S.Y.; Ding, R.X.; Cang, W.H.; Wu, R.N.; Wu, J.R. Role of the dietary components in food allergy: A comprehensive review. Food Chem. 2022, 386, 12. [Google Scholar] [CrossRef]
- Bessa, C.; Francisco, T.; Dias, R.; Mateus, N.; de Freitas, V.; Perez-Gregorio, R. Use of Polyphenols as Modulators of Food Allergies. From Chemistry to Biological Implications. Front. Sustain. Food Syst. 2021, 5, 18. [Google Scholar] [CrossRef]
- Ohishi, T.; Hayakawa, S.; Miyoshi, N. Involvement of microRNA modifications in anticancer effects of major polyphenols from green tea, coffee, wine, and curry. Crit. Rev. Food Sci. Nutr. 2022, 1–32. [Google Scholar] [CrossRef]
- Plundrich, N.J.; Bansode, R.R.; Foegeding, E.A.; Williams, L.L.; Lila, M.A. Protein-bound Vaccinium fruit polyphenols decrease IgE binding to peanut allergens and RBL-2H3 mast cell degranulation in vitro. Food Funct. 2017, 8, 1611–1621. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Mattison, C.P. Polyphenol-rich pomegranate juice reduces IgE binding to cashew nut allergens. J. Sci. Food Agric. 2018, 98, 1632–1638. [Google Scholar] [CrossRef]
- Sun, S.F.; Jiang, T.Y.; Gu, Y.J.; Yao, L.; Du, H.; Luo, J.Z.; Che, H.L. Contribution of five major apple polyphenols in reducing peanut protein sensitization and alleviating allergencitiy of peanut by changing allergen structure. Food Res. Int. 2023, 164, 10. [Google Scholar] [CrossRef]
- Yan, X.H.; Zeng, Z.L.; McClements, D.J.; Gong, X.F.; Yu, P.; Xia, J.H.; Gong, D.M. A review of the structure, function, and application of plant-based protein-phenolic conjugates and complexes. Compr. Rev. Food Sci. Food Saf. 2023, 22, 1312–1336. [Google Scholar] [CrossRef]
- Hagemann, P.M.; Nsiah-Dosu, S.; Hundt, J.E.; Hartmann, K.; Orinska, Z. Modulation of Mast Cell Reactivity by Lipids: The Neglected Side of Allergic Diseases. Front. Immunol. 2019, 10, 1174. [Google Scholar] [CrossRef]
- Gilfillan, A.M.; Tkaczyk, C. Integrated signalling pathways for mast-cell activation. Nat. Rev. Immunol. 2006, 6, 218–230. [Google Scholar] [CrossRef]
- Bankova, L.G.; Lai, J.Y.; Yoshimoto, E.; Boyce, J.A.; Austen, K.F.; Kanaoka, Y.; Barrett, N.A. Leukotriene E-4 elicits respiratory epithelial cell mucin release through the G-protein-coupled receptor, GPR99. Proc. Natl. Acad. Sci. USA 2016, 113, 6242–6247. [Google Scholar] [CrossRef] [PubMed]
- Schauberger, E.; Peinhaupt, M.; Cazares, T.; Lindsley, A.W. Lipid Mediators of Allergic Disease: Pathways, Treatments, and Emerging Therapeutic Targets. Curr. Allergy Asthma Rep. 2016, 16, 48. [Google Scholar] [CrossRef]
- Han, X.L.; Gross, R.W. Global analyses of cellular lipidomes directly from crude extracts of biological samples by ESI mass spectrometry: A bridge to lipidomics. J. Lipid Res. 2003, 44, 1071–1079. [Google Scholar] [CrossRef]
- Abu-Hijleh, H.M.; Al-Zoubi, R.M.; Zarour, A.; Ansari, A.A.; Bawadi, H. The Therapeutic Role of Curcumin in Inflammation and Post-Surgical Outcomes. Food Rev. Int. 2023, 16. [Google Scholar] [CrossRef]
- Mokra, D.; Joskova, M.; Mokry, J. Therapeutic Effects of Green Tea Polyphenol (‒)-Epigallocatechin-3-Gallate (EGCG) in Relation to Molecular Pathways Controlling Inflammation, Oxidative Stress, and Apoptosis. Int. J. Mol. Sci. 2023, 24, 340. [Google Scholar] [CrossRef]
- Matsui, T.; Ito, C.; Itoigawa, M.; Shibata, T. Three phlorotannins from Sargassum carpophyllum are effective against the secretion of allergic mediators from antigen-stimulated rat basophilic leukemia cells. Food Chem. 2022, 377, 131992. [Google Scholar] [CrossRef]
- Tang, F.; Chen, F.; Ling, X.; Huang, Y.; Zheng, X.; Tang, Q.; Tan, X. Inhibitory effect of methyleugenol on IgE-mediated allergic inflammation in RBL-2H3 cells. Mediat. Inflamm. 2015, 2015, 463530. [Google Scholar] [CrossRef]
- Nagai, H.; Abe, T.; Yamaguchi, I.; Mito, K.; Tsunematsu, M.; Kimata, M.; Inagaki, N. Role of mast cells in the onset of IgE-mediated late-phase cutaneous response in mice. J. Allergy Clin. Immunol. 2000, 106, S91–S98. [Google Scholar] [CrossRef]
- Zeng, J.; Li, J.L.; Liu, S.S.Y.; Yang, Z.Q.; Zhong, Y.; Chen, X.M.; Li, G.L.; Li, J. Lipidome disturbances in preadipocyte differentiation associated with bisphenol A and replacement bisphenol S exposure. Sci. Total Environ. 2021, 753, 10. [Google Scholar] [CrossRef]
- Zeng, J.; Liu, S.; Cai, W.; Jiang, H.; Lu, X.; Li, G.; Li, J.; Liu, J. Emerging lipidome patterns associated with marine Emiliania huxleyi-virus model system. Sci. Total Environ. 2019, 688, 521–528. [Google Scholar] [CrossRef]
- Li, J.; Yuan, H.B.; Rong, Y.T.; Qian, M.C.; Liu, F.Q.; Hua, J.J.; Zhou, Q.H.; Deng, Y.L.; Zeng, J.; Jiang, Y.W. Lipid metabolic characteristics and marker compounds of ripened Pu-erh tea during pile fermentation revealed by LC-MS-based lipidomics. Food Chem. 2023, 404, 12. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Ito, C.; Masubuchi, S.; Itoigawa, M. Licarin A is a candidate compound for the treatment of immediate hypersensitivity via inhibition of rat mast cell line RBL-2H3 cells. J. Pharm. Pharmacol. 2015, 67, 1723–1732. [Google Scholar] [CrossRef]
- Singh, B.K.; Kambayashi, T. The Immunomodulatory Functions of Diacylglycerol Kinase zeta. Front. Cell Dev. Biol. 2016, 4, 96. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, M.; Shirai, Y.; Ueyama, T.; Saito, N. Diacylglycerol kinase gamma regulates antigen-induced mast cell degranulation by mediating Ca(2+) influxes. Biochem. Biophys. Res. Commun. 2014, 445, 340–345. [Google Scholar] [CrossRef]
- Arita, M. Eosinophil polyunsaturated fatty acid metabolism and its potential control of inflammation and allergy. Allergol. Int. 2016, 65, S2–S5. [Google Scholar] [CrossRef]
- Wang, X.; Kulka, M. n-3 Polyunsaturated fatty acids and mast cell activation. J. Leukoc. Biol. 2015, 97, 859–871. [Google Scholar] [CrossRef]
- Sugiyama, H.; Nonaka, T.; Kishimoto, T.; Komoriya, K.; Tsuji, K.; Nakahata, T. Peroxisome proliferator-activated receptors are expressed in human cultured mast cells: A possible role of these receptors in negative regulation of mast cell activation. Eur. J. Immunol. 2000, 30, 3363–3370. [Google Scholar] [CrossRef]
- Tachibana, M.; Wada, K.; Katayama, K.; Kamisaki, Y.; Maeyama, K.; Kadowaki, T.; Blumberg, R.S.; Nakajima, A. Activation of peroxisome proliferator-activated receptor gamma suppresses mast cell maturation involved in allergic diseases. Allergy 2008, 63, 1136–1147. [Google Scholar] [CrossRef]
- Field, K.A.; Holowka, D.; Baird, B. Compartmentalized activation of the high affinity immunoglobulin E receptor within membrane domains. J. Biol. Chem. 1997, 272, 4276–4280. [Google Scholar] [CrossRef]
- Simons, K.; Sampaio, J.L. Membrane Organization and Lipid Rafts. Cold Spring Harb. Perspect. Biol. 2011, 3, 17. [Google Scholar] [CrossRef]
- Trinchese, G.; Paparo, L.; Aitoro, R.; Fierro, C.; Varchetta, M.; Nocerino, R.; Mollica, M.P.; Berni Canani, R. Hepatic Mitochondrial Dysfunction and Immune Response in a Murine Model of Peanut Allergy. Nutrients 2018, 10, 744. [Google Scholar] [CrossRef]
- Showalter, M.R.; Berg, A.L.; Nagourney, A.; Heil, H.; Carraway, K.L., 3rd; Fiehn, O. The Emerging and Diverse Roles of Bis(monoacylglycero) Phosphate Lipids in Cellular Physiology and Disease. Int. J. Mol. Sci. 2020, 21, 8067. [Google Scholar] [CrossRef]
- Akgoc, Z.; Sena-Esteves, M.; Martin, D.R.; Han, X.; d’Azzo, A.; Seyfried, T.N. Bis(monoacylglycero)phosphate: A secondary storage lipid in the gangliosidoses. J. Lipid Res. 2015, 56, 1006–1013. [Google Scholar] [CrossRef]
- Gallala, H.D.; Sandhoff, K. Biological function of the cellular lipid BMP-BMP as a key activator for cholesterol sorting and membrane digestion. Neurochem. Res. 2011, 36, 1594–1600. [Google Scholar] [CrossRef]
- Grabner, G.F.; Fawzy, N.; Pribasnig, M.A.; Trieb, M.; Taschler, U.; Holzer, M.; Schweiger, M.; Wolinski, H.; Kolb, D.; Horvath, A.; et al. Metabolic disease and ABHD6 alter the circulating bis(monoacylglycerol)phosphate profile in mice and humans. J. Lipid Res. 2019, 60, 1020–1031. [Google Scholar] [CrossRef]
- Groot Kormelink, T.; Arkesteijn, G.J.; van de Lest, C.H.; Geerts, W.J.; Goerdayal, S.S.; Altelaar, M.A.; Redegeld, F.A.; Nolte-’t Hoen, E.N.; Wauben, M.H. Mast Cell Degranulation Is Accompanied by the Release of a Selective Subset of Extracellular Vesicles That Contain Mast Cell-Specific Proteases. J. Immunol. 2016, 197, 3382–3392. [Google Scholar] [CrossRef]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, U654–U672. [Google Scholar] [CrossRef]
- Skokos, D.; Le Panse, S.; Villa, I.; Rousselle, J.C.; Peronet, R.; David, B.; Namane, A.; Mecheri, S. Mast cell-dependent B and T lymphocyte activation is mediated by the secretion of immunologically active exosomes. J. Immunol. 2001, 166, 868–876. [Google Scholar] [CrossRef]
- Skokos, D.; Botros, H.G.; Demeure, C.; Morin, J.; Peronet, R.; Birkenmeier, G.; Boudaly, S.; Mecheri, S. Mast cell-derived exosomes induce phenotypic and functional maturation of dendritic cells and elicit specific immune responses in vivo. J. Immunol. 2003, 170, 3037–3045. [Google Scholar] [CrossRef]
- Snyder, F. Platelet-Activating-Factor—The Biosynthetic and Catabolic Enzymes. Biochem. J. 1995, 305, 689–705. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Fatty Acids | Veh vs. AG | Curcumin vs. AG | EGCG vs. AG | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| p | FDR | AG/Veh | p | FDR | Cur/AG | p | FDR | EGCG/AG | |||||
| Ratio-1 h | Ratio-3 h | Ratio-1 h | Ratio-3 h | Ratio-1 h | Ratio-3 h | ||||||||
| CAR 24:0 | 24:0 | 4.51 × 10−10 | 1.27 × 10−8 | 0.611 | 0.522 | 9.38 × 10−4 | 2.64 × 10−3 | 1.159 | 1.240 | 3.52 × 10−1 | 4.63 × 10−1 | 0.849 | 1.241 |
| CAR 24:1 | 24:1 | 1.77 × 10−10 | 6.10 × 10−9 | 0.580 | 0.582 | 1.48 × 10−4 | 5.12 × 10−4 | 1.103 | 1.350 | 6.81 × 10−2 | 1.27 × 10−1 | 0.861 | 1.220 |
| DG 38:4 | 18:0_20:4 | 3.90 × 10−14 | 1.82 × 10−11 | 11.091 | 2.447 | 5.23 × 10−1 | 6.16 × 10−1 | 0.913 | 1.229 | 1.44 × 10−5 | 1.11 × 10−4 | 1.495 | 1.743 |
| DG 40:5 | 18:0_22:5 | 1.64 × 10−13 | 4.88 × 10−11 | 7.501 | 1.702 | 1.56 × 10−1 | 2.37 × 10−1 | 0.969 | 1.320 | 1.16 × 10−4 | 6.10 × 10−4 | 1.372 | 1.397 |
| DG 40:6 | 18:0_22:6 | 2.62 × 10−13 | 4.88 × 10−11 | 7.822 | 1.649 | 1.54 × 10−2 | 3.22 × 10−2 | 1.025 | 1.351 | 1.08 × 10−4 | 5.81 × 10−4 | 1.399 | 1.324 |
| HBMP 58:8 | 18:1_18:1_22:6 | 2.76 × 10−9 | 5.47 × 10−8 | 0.648 | 0.643 | 3.19 × 10−4 | 1.00 × 10−3 | 1.081 | 1.278 | 2.01 × 10−1 | 3.00 × 10−1 | 1.080 | 0.992 |
| HBMP 62:13 | 18:1_22:6_22:6 | 1.72 × 10−11 | 1.11 × 10−9 | 0.582 | 0.529 | 7.29 × 10−1 | 7.93 × 10−1 | 0.855 | 1.158 | 6.06 × 10−2 | 1.14 × 10−1 | 1.018 | 1.111 |
| HBMP 64:16 | 20:4_22:6_22:6 | 3.66 × 10−12 | 3.41 × 10−10 | 0.569 | 0.478 | 2.99 × 10−3 | 7.68 × 10−3 | 0.725 | 1.041 | 5.32 × 10−2 | 1.02 × 10−1 | 0.999 | 1.140 |
| LPC-O 22:0 | O-22:0 | 2.52 × 10−11 | 1.30 × 10−9 | 0.658 | 0.581 | 1.35 × 10−7 | 2.42 × 10−6 | 1.209 | 1.477 | 5.04 × 10−4 | 1.90 × 10−3 | 1.045 | 1.163 |
| LPC-O 24:1 | O-24:1 | 7.71 × 10−10 | 1.99 × 10−8 | 0.584 | 0.524 | 8.44 × 10−9 | 7.14 × 10−7 | 1.287 | 1.517 | 2.89 × 10−2 | 6.04 × 10−2 | 1.060 | 1.052 |
| PEtOH 34:1 | 16:0_18:1 | 2.64 × 10−9 | 5.34 × 10−8 | 2.239 | 4.310 | 1.86 × 10−1 | 2.71 × 10−1 | 0.670 | 1.121 | 1.98 × 10−6 | 2.49 × 10−5 | 1.728 | 1.197 |
| PEtOH 36:1 | 18:0_18:1 | 5.14 × 10−8 | 6.47 × 10−7 | 1.511 | 2.358 | 1.09 × 10−1 | 1.77 × 10−1 | 0.696 | 1.047 | 8.64 × 10−5 | 4.82 × 10−4 | 1.524 | 1.156 |
| PG 36:4 | 16:0_20:4 | 1.79 × 10−11 | 1.11 × 10−9 | 1.505 | 1.650 | 3.32 × 10−4 | 1.03 × 10−3 | 0.795 | 1.007 | 3.62 × 10−7 | 6.02 × 10−6 | 1.088 | 1.328 |
| PG 38:4 | 18:0_20:4 | 5.71 × 10−12 | 4.83 × 10−10 | 3.010 | 3.106 | 9.49 × 10−4 | 2.66 × 10−3 | 0.723 | 0.993 | 3.01 × 10−7 | 5.22 × 10−6 | 1.341 | 1.634 |
| PI 36:5 | 16:0_20:5 | 1.42 × 10−12 | 1.47 × 10−10 | 2.175 | 2.374 | 2.15 × 10−5 | 1.02 × 10−4 | 1.043 | 1.197 | 9.42 × 10−1 | 9.68 × 10−1 | 0.864 | 1.135 |
| PI 42:9 | 20:4_22:5 | 7.28 × 10−13 | 9.68 × 10−11 | 3.257 | 2.087 | 9.68 × 10−5 | 3.62 × 10−4 | 0.759 | 1.061 | 1.86 × 10−2 | 4.14 × 10−2 | 0.939 | 1.321 |
| PI-O 36:4 | O-16:0_20:4 | 1.72 × 10−10 | 6.10 × 10−9 | 1.691 | 1.759 | 9.41 × 10−2 | 1.57 × 10−1 | 0.905 | 1.012 | 3.72 × 10−1 | 4.83 × 10−1 | 0.859 | 1.225 |
| PI-O 38:6 | O-16:0_22:6 | 5.88 × 10−10 | 1.61 × 10−8 | 1.556 | 1.658 | 2.83 × 10−3 | 7.32 × 10−3 | 1.035 | 1.127 | 9.71 × 10−2 | 1.69 × 10−1 | 0.868 | 0.999 |
| PI-O 40:7 | O-18:1_22:6 | 1.26 × 10−12 | 1.47 × 10−10 | 1.546 | 1.894 | 4.01 × 10−1 | 5.09 × 10−1 | 1.013 | 1.010 | 3.18 × 10−2 | 6.56 × 10−2 | 0.945 | 0.924 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, J.; Hao, J.; Yang, Z.; Ma, C.; Gao, L.; Chen, Y.; Li, G.; Li, J. Anti-Allergic Effect of Dietary Polyphenols Curcumin and Epigallocatechin Gallate via Anti-Degranulation in IgE/Antigen-Stimulated Mast Cell Model: A Lipidomics Perspective. Metabolites 2023, 13, 628. https://doi.org/10.3390/metabo13050628
Zeng J, Hao J, Yang Z, Ma C, Gao L, Chen Y, Li G, Li J. Anti-Allergic Effect of Dietary Polyphenols Curcumin and Epigallocatechin Gallate via Anti-Degranulation in IgE/Antigen-Stimulated Mast Cell Model: A Lipidomics Perspective. Metabolites. 2023; 13(5):628. https://doi.org/10.3390/metabo13050628
Chicago/Turabian StyleZeng, Jun, Jingwen Hao, Zhiqiang Yang, Chunyu Ma, Longhua Gao, Yue Chen, Guiling Li, and Jia Li. 2023. "Anti-Allergic Effect of Dietary Polyphenols Curcumin and Epigallocatechin Gallate via Anti-Degranulation in IgE/Antigen-Stimulated Mast Cell Model: A Lipidomics Perspective" Metabolites 13, no. 5: 628. https://doi.org/10.3390/metabo13050628
APA StyleZeng, J., Hao, J., Yang, Z., Ma, C., Gao, L., Chen, Y., Li, G., & Li, J. (2023). Anti-Allergic Effect of Dietary Polyphenols Curcumin and Epigallocatechin Gallate via Anti-Degranulation in IgE/Antigen-Stimulated Mast Cell Model: A Lipidomics Perspective. Metabolites, 13(5), 628. https://doi.org/10.3390/metabo13050628