
Dynamic Changes in Plant Secondary Metabolites Induced by Botrytis cinerea Infection


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plants and Pathogens
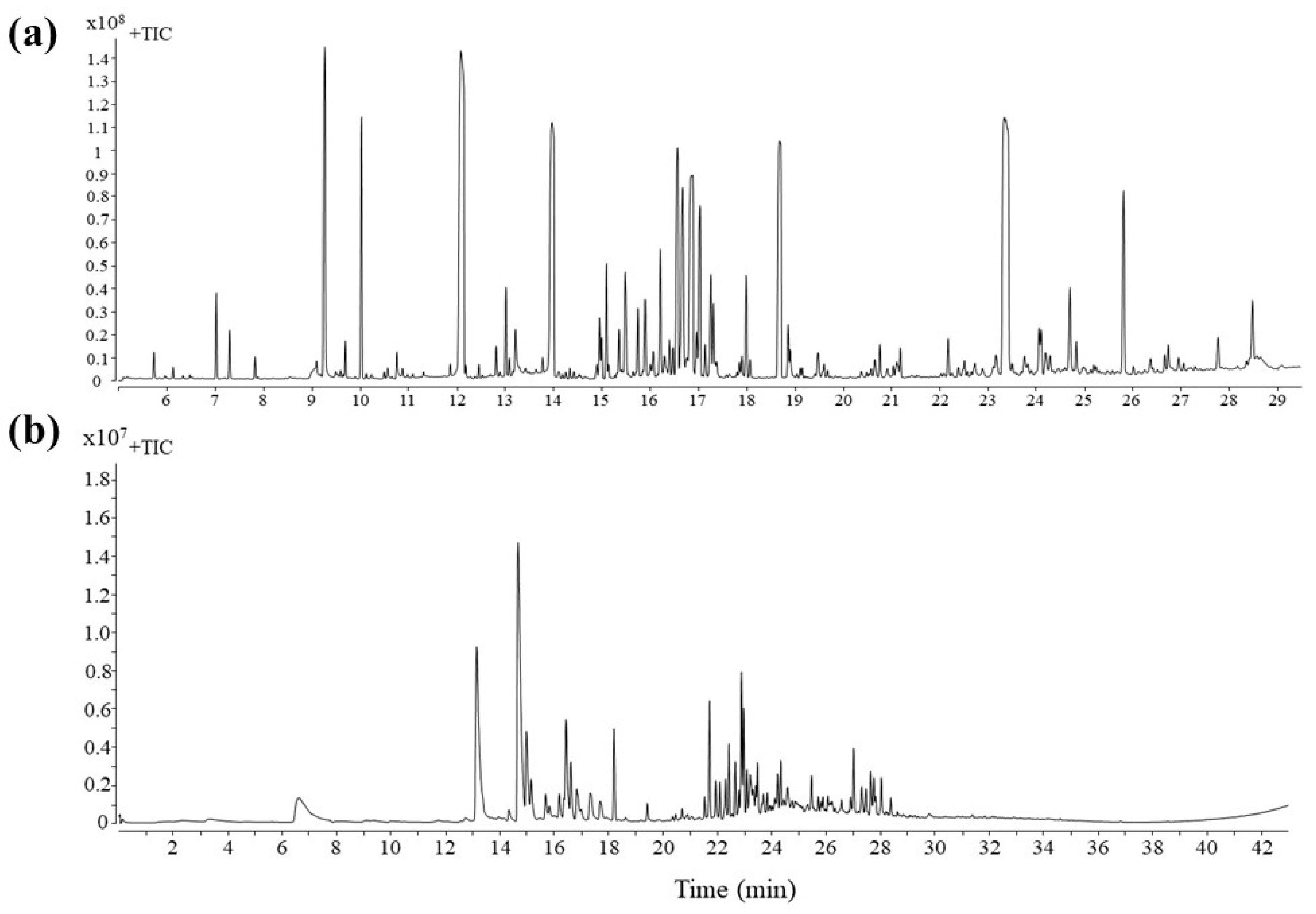
2.3. Nonvolatile Metabolites of Grape Leaves
2.4. Volatile Metabolites of Grape Leaves
2.5. Effects of Secondary Metabolites on Botrytis cinerea Growth
2.6. Statistical Analysis
3. Results
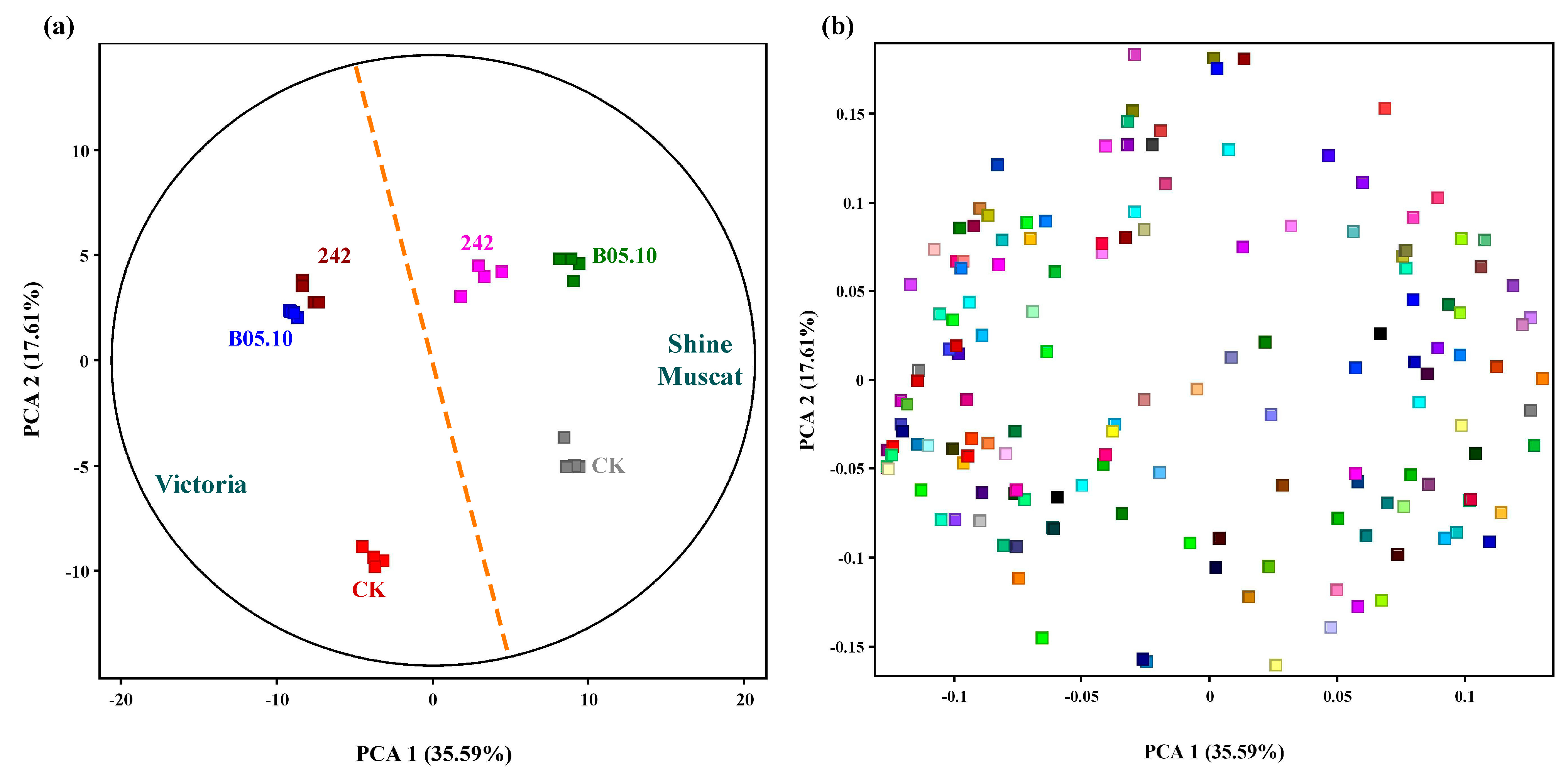
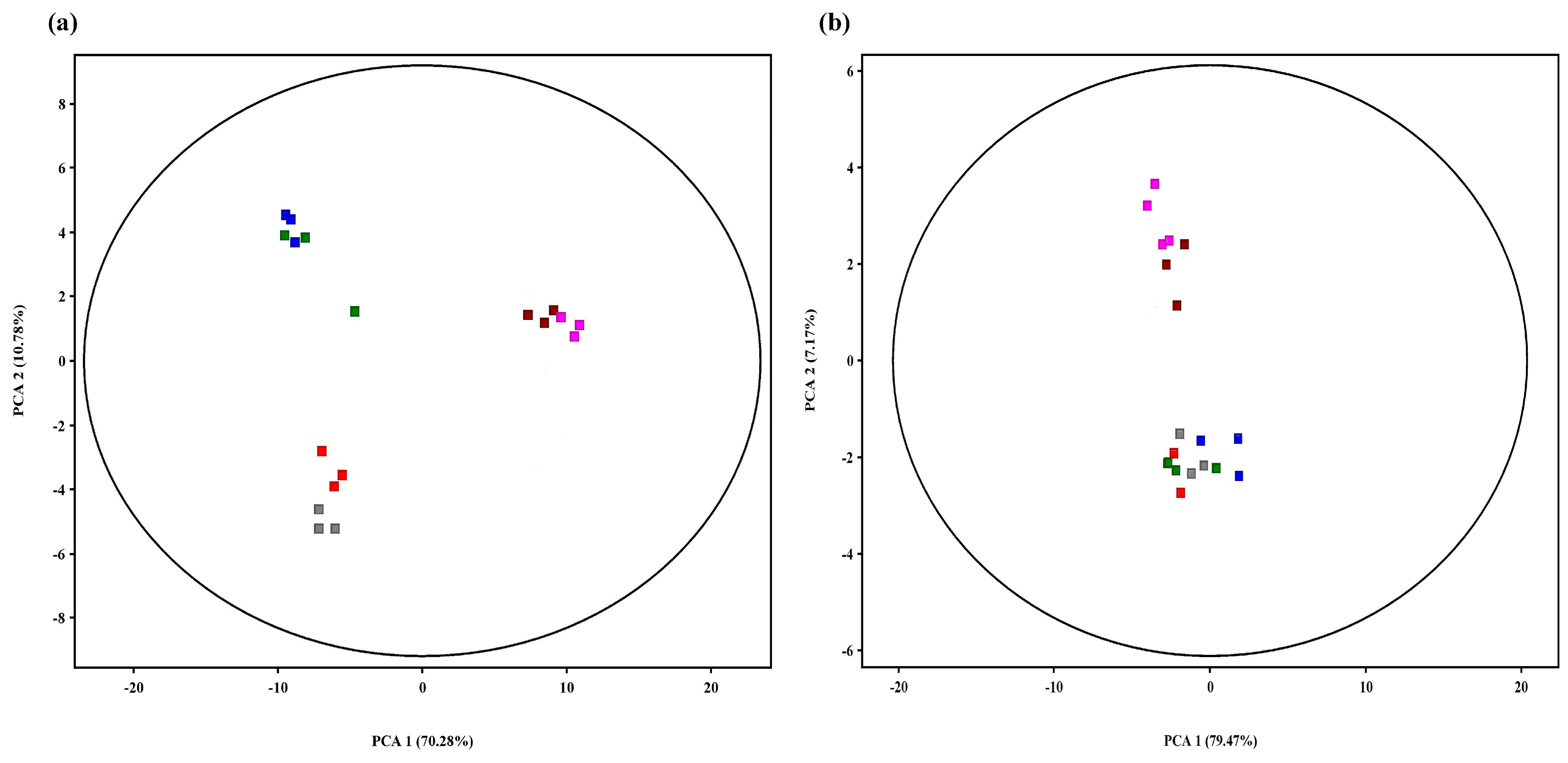
3.1. Nonvolatile Metabolites
3.2. Characterization of Significantly Changed Nonvolatile Differential Metabolites
3.3. Identification of Volatile Metabolites
3.4. Crucially Changed Volatile Metabolites
3.5. Effect of Botrytis cinerea on Metabolic Pathways of Grapes
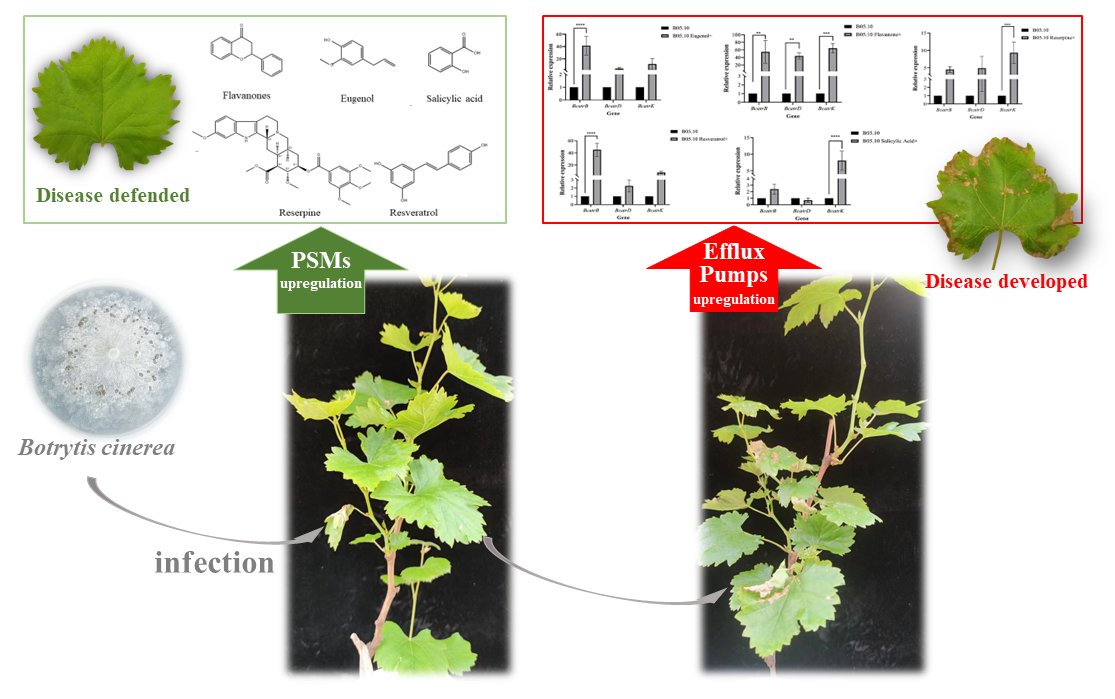
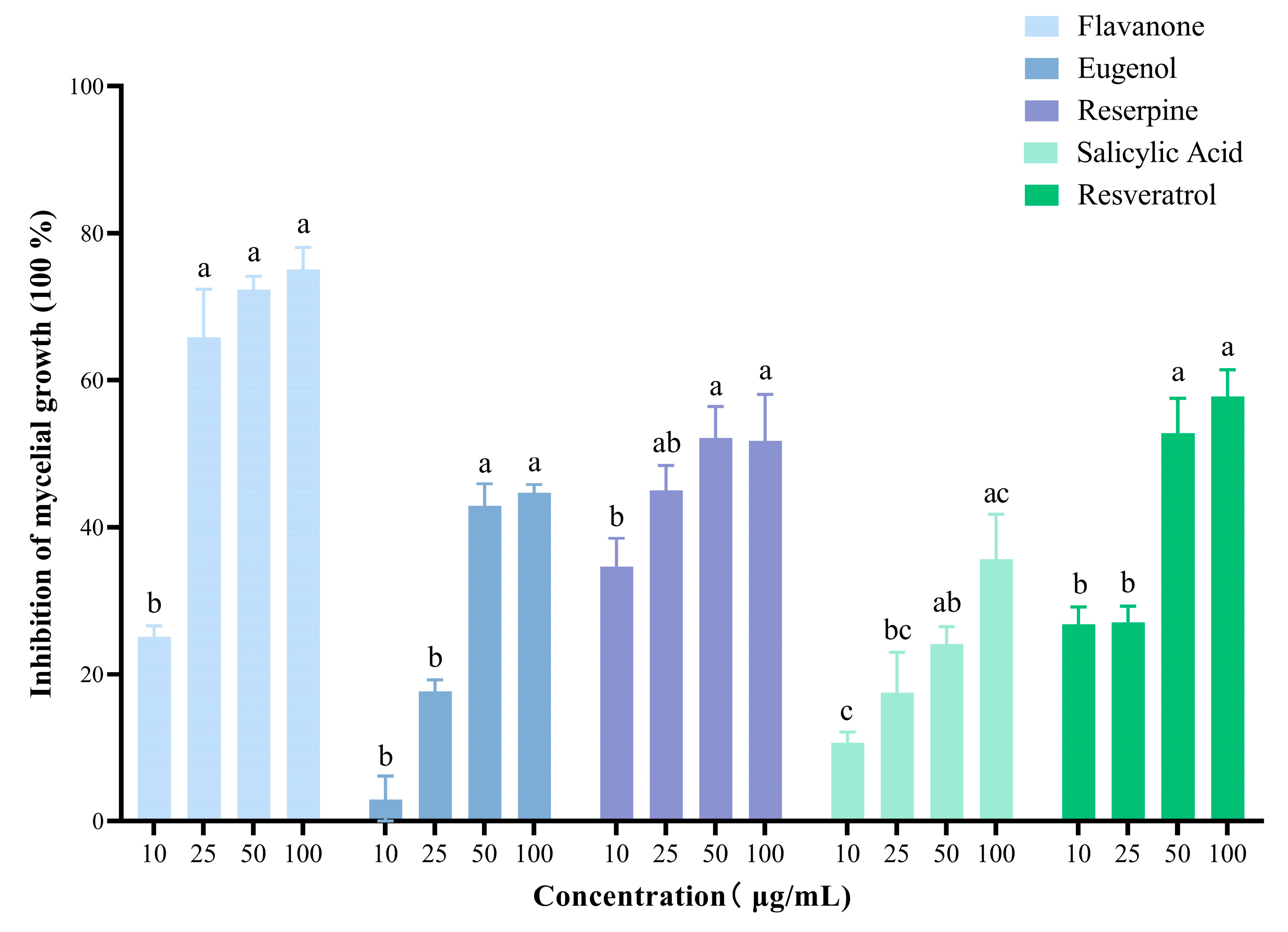
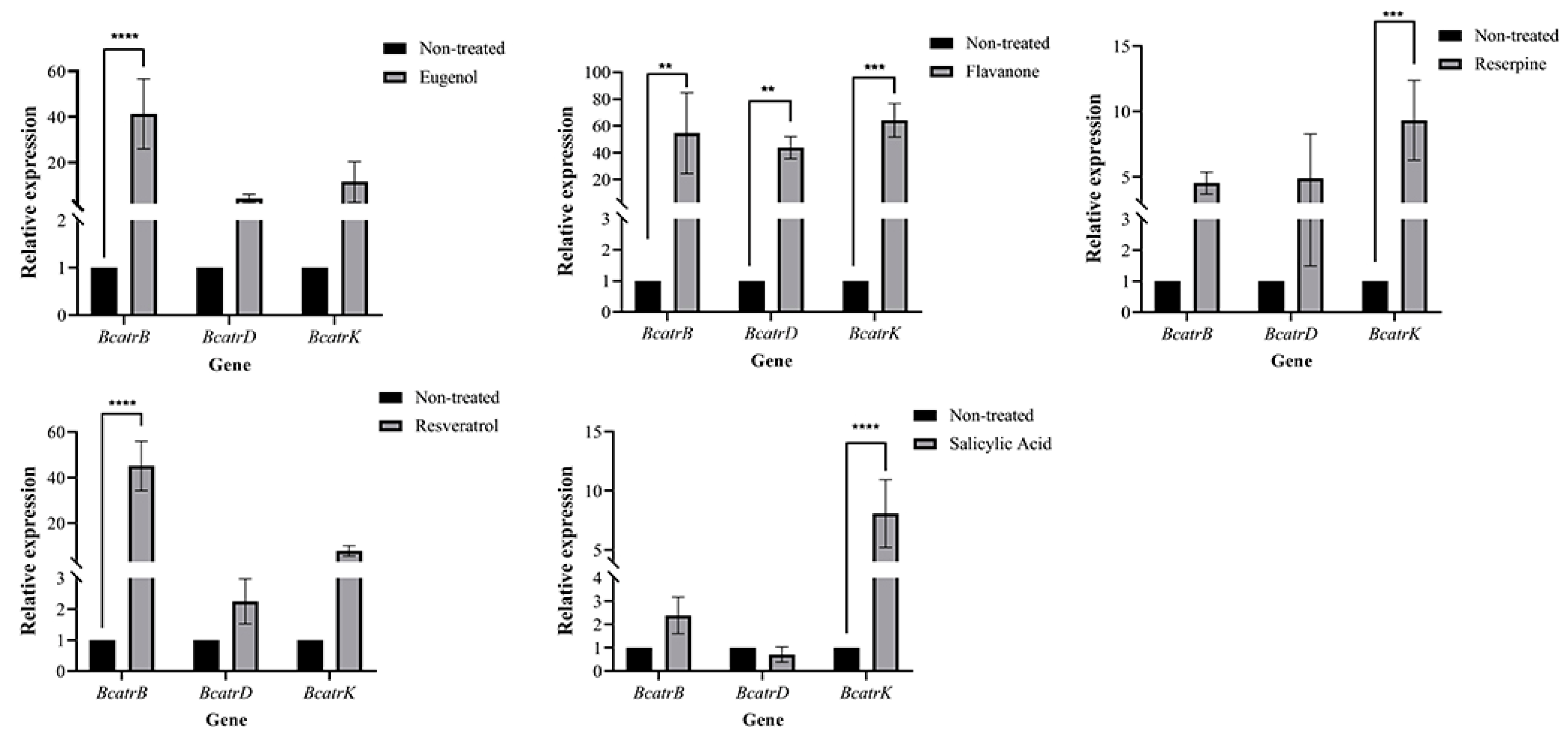
3.6. Bioactivity of Major Metabolites on Botrytis cinerea
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhan, C.; Shen, S.; Yang, C.; Liu, Z.; Fernie, A.R.; Graham, I.A.; Luo, J. Plant metabolic gene clusters in the multi-omics era. Trends Plant Sci. 2022, 27, 981–1001. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, J.P.; Osbourn, A.E. Fungal resistance to plant antibiotics as a mechanism of pathogenesis. Microbiol. Mol. Biol. Rev. 1999, 63, 708–724. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, K.; Matsui, K.; Ozawa, R.; Takabayashi, J. Volatile C6-aldehydes and allo-ocimene activate defense genes and induce resistance against Botrytis cinerea in Arabidopsis thaliana. Plant Cell Physiol. 2005, 46, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, K.; Matsui, K.; Ozawa, R.; Takabayashi, J. Analysis of defensive responses activated by volatile allo-ocimene treatment in Arabidopsis thaliana. Phytochemistry 2006, 67, 1520–1529. [Google Scholar] [CrossRef]
- Vio-Michaelis, S.; Apablaza-Hidalgo, G.; Gómez, M.; Peña-Vera, R.; Montenegro, G. Antifungal activity of three Chilean plant extracts on Botrytis cinerea. Bot. Sci. 2012, 90, 179–183. [Google Scholar] [CrossRef]
- Soylu, E.M.; Kurt, Ş.; Soylu, S. In vitro and in vivo antifungal activities of the essential oils of various plants against tomato grey mould disease agent Botrytis cinerea. Int. J. Food. Microbiol. 2010, 143, 183–189. [Google Scholar] [CrossRef]
- Wilson, C.L.; Solar, J.M.; El Ghaouth, A.; Wisniewski, M.E. Rapid evaluation of plant extracts and essential oils for antifungal activity against Botrytis cinerea. Plant Dis. 1997, 81, 204–210. [Google Scholar] [CrossRef]
- Monteiro, S.; Barakat, M.; Piçarra-Pereira, M.A.; Teixeira, A.R.; Ferreira, R.B. Osmotin and thaumatin from grape: A putative general defense mechanism against pathogenic fungi. Phytopathology 2003, 93, 1505–1512. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Nosek, M.; Kornaś, A.; Kuźniak, E.; Miszalski, Z. Plastoquinone redox state modifies plant response to pathogen. Plant Physiol. Biochem. 2015, 96, 163–170. [Google Scholar] [CrossRef]
- Hu, Z.; Shao, S.; Zheng, C.; Sun, Z.; Shi, J.; Yu, J.; Qi, Z.; Shi, K. Induction of systemic resistance in tomato against Botrytis cinerea by N-decanoyl-homoserine lactone via jasmonic acid signaling. Planta 2018, 247, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yang, F.; Tong, H.; Hu, Y.; Zhang, X.; Tian, T.; Zhang, Y.; Su, Q. Plant flavonoids enhance the tolerance to thiamethoxam and flupyradifurone in whitefly Bemisia tabaci (Hemiptera: Aleyrodidae). Pestic. Biochem. Physiol. 2021, 171, 104744. [Google Scholar] [CrossRef]
- Chen, C.; Liu, Y.; Shi, X.; Desneux, N.; Han, P.; Gao, X. Elevated carboxylesterase activity contributes to the lambda-cyhalothrin insensitivity in quercetin fed Helicoverpa armigera (Hübner). PLoS ONE 2017, 12, e0183111. [Google Scholar] [CrossRef]
- Chen, C.; Han, P.; Yan, W.; Wang, S.; Shi, X.; Zhou, X.; Desneux, N.; Gao, X. Uptake of quercetin reduces larval sensitivity to lambda-cyhalothrin in Helicoverpa armigera. J. Pest. Sci. 2018, 91, 919–926. [Google Scholar] [CrossRef]
- Meirelles, L.A.; Perry, E.K.; Bergkessel, M.; Newman, D.K. Bacterial defenses against a natural antibiotic promote collateral resilience to clinical antibiotics. PloS. Biol. 2021, 19, e3001093. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Xie, W.; Yang, X.; Wang, S.L.; Wu, Q.J.; Zhang, Y.J. Identification of glutathione S-transferases in Bemisia tabaci (Hemiptera: Aleyrodidae) and evidence that GSTd7 helps explain the difference in insecticide susceptibility between B. tabaci Middle East-Minor Asia 1 and Mediterranean. Insect Mol. Biol. 2018, 27, 22–35. [Google Scholar] [CrossRef]
- Amezian, D.; Nauen, R.; Le Goff, G. Transcriptional regulation of xenobiotic detoxification genes in insects-An overview. Pestic. Biochem. Physiol. 2021, 174, 104822. [Google Scholar] [CrossRef]
- Amezian, D.; Nauen, R.; Le Goff, G. Comparative analysis of the detoxification gene inventory of four major Spodoptera pest species in response to xenobiotics. Insect Biochem. Mol. Biol. 2021, 138, 103646. [Google Scholar] [CrossRef]
- Frías, M.; Brito, N.; González, C. The Botrytis cinerea cerato-platanin BcSpl1 is a potent inducer of systemic acquired resistance (SAR) in tobacco and generates a wave of salicylic acid expanding from the site of application. Mol. Plant. Pathol. 2013, 14, 191–196. [Google Scholar] [CrossRef]
- Jeandet, P.; Sbaghi, M.; Bessis, R. The significance of stilbene-type phytoalexin degradation by culture filtrates of Botrytis cinerea in the vine-Botrytis interaction. Mech. Plant Def. Responses 1993, 1993, 84. [Google Scholar] [CrossRef]
- Wang, W.; Fang, Y.; Imran, M.; Hu, Z.; Zhang, S.; Huang, Z.; Liu, X. Characterization of the field fludioxonil resistance and its molecular basis in Botrytis cinerea from Shanghai Province in China. Microorganisms 2021, 9, 266. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.C.; Wang, G.Z.; Zhang, B.R.; Dai, T.; Gu, A.Y.; Li, X.L.; Cheng, X.K.; Liu, P.F.; Hao, J.J.; Liu, X.L. Metabolic mechanism of plant defense against rice blast induced by probenazole. Metabolites 2021, 11, 246. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhu, Y.; Ma, W.; Shi, J.; Peng, Q.; Lin, Z.; Lv, H. Comprehensive investigation on non-volatile and volatile metabolites in four types of green teas obtained from the same tea cultivar of Longjing 43 (Camellia sinensis var. sinensis) using the widely targeted metabolomics. Food Chem. 2022, 394, 133501. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhang, Z.; Chen, L.; Chen, L.; He, L.; Lu, H.; Ren, Y.; Mu, W.; Liu, F.; Pests, I. Baseline sensitivity of Botrytis cinerea to the succinate dehydrogenase inhibitor isopyrazam and efficacy of this fungicide. Plant Dis. 2016, 100, 1314–1320. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Peluffo, L.; Lia, V.; Troglia, C.; Maringolo, C.; Norma, P.; Escande, A.; Esteban Hopp, H.; Lytovchenko, A.; Fernie, A.R.; Heinz, R.; et al. Metabolic profiles of sunflower genotypes with contrasting response to Sclerotinia sclerotiorum infection. Phytochemistry 2010, 71, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Wang, Y.; Ma, Y.; Su, W.; Bai, Y.; Zhao, P. A rapid LC/MS/MS quantitation assay for naringin and its two metabolites in rats plasma. J. Pharm. Biomed. Anal. 2006, 40, 454–459. [Google Scholar] [CrossRef]
- Chen, F.C.; Lin, Y.M. Rhusflavanone, a new biflavanone from the seeds of wax-tree. J. Chem. Soc. Perkin Trans. 1976, 1, 98–101. [Google Scholar] [CrossRef]
- Careri, M.; Corradini, C.; Elviri, L.; Nicoletti, I.; Zagnoni, I. Liquid chromatography−electrospray tandem mass spectrometry of cis-resveratrol and trans-resveratrol: Development, validation, and application of the method to red wine, grape, and winemaking byproducts. J. Agric. Food Chem. 2004, 52, 6868–6874. [Google Scholar] [CrossRef]
- Radko, Y.; Christensen, K.B.; Christensen, L.P. Semi-preparative isolation of dihydroresveratrol-3-O-β-d-glucuronide and four resveratrol conjugates from human urine after oral intake of a resveratrol-containing dietary supplement. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2013, 930, 54–61. [Google Scholar] [CrossRef]
- Ramesh, S.A.; Tyerman, S.D.; Gilliham, M.; Xu, B. γ-Aminobutyric acid (GABA) signalling in plants. Cell Mol. Life Sci. 2017, 74, 1577–1603. [Google Scholar] [CrossRef] [PubMed]
- Romero-Pérez, A.I.; Ibern-Gómez, M.; Lamuela-Raventós, R.M.; Torre-Boronat, M.C.D. Piceid, the major resveratrol derivative in grape juices. J. Agric. Food Chem. 1999, 47, 1533–1536. [Google Scholar] [CrossRef] [PubMed]
- Galvez, C.E.; Jimenez, C.M.; Gomez, A.A.; Lizarraga, E.F.; Sampietro, D.A. Chemical composition and antifungal activity of essential oils from Senecio nutans, Senecio viridis, Tagetes terniflora and Aloysia gratissima against toxigenic Aspergillus and Fusarium species. Nat. Prod. Res. 2020, 34, 1442–1445. [Google Scholar] [CrossRef] [PubMed]
- Souleyre, E.J.F.; Bowen, J.K.; Matich, A.J.; Tomes, S.; Chen, X.Y.; Hunt, M.B.; Wang, M.Y.; Ileperuma, N.R.; Richards, K.; Rowan, D.D.; et al. Genetic control of α-farnesene production in apple fruit and its role in fungal pathogenesis. Plant J. 2019, 100, 1148–1162. [Google Scholar] [CrossRef] [PubMed]
- Abou El-Soud, N.H.; Deabes, M.; Abou El-Kassem, L.; Khalil, M. Chemical composition and antifungal activity of Ocimum basilicum L. essential oil. Maced. J. Med. Sci. 2015, 3, 374. [Google Scholar] [CrossRef]
- Lawson, S.K.; Sharp, L.G.; Satyal, P.; Setzer, W.N. Volatile components of the aerial parts of Prunella vulgaris L. (Lamiaceae). Am. J. Essent. Oils Nat. Prod. 2020, 8, 17–19. [Google Scholar]
- Rudramurthy, S.M.; Paul, R.A.; Chakrabarti, A.; Mouton, J.W.; Meis, J.F. Invasive aspergillosis by Aspergillus flavus: Epidemiology, diagnosis, antifungal resistance, and management. J. Fungi 2019, 5, 55. [Google Scholar] [CrossRef]
- Rafiei, V.; Ruffino, A.; Persson Hodén, K.; Tornkvist, A.; Mozuraitis, R.; Dubey, M.; Tzelepis, G. A Verticillium longisporum pleiotropic drug transporter determines tolerance to the plant host β-pinene monoterpene. Mol. Plant Pathol. 2022, 23, 291–303. [Google Scholar] [CrossRef]
- Elhawary, E.A.; Mostafa, N.M.; Labib, R.M.; Singab, A.N. Metabolomic profiles of essential oils from selected rosa varieties and their antimicrobial activities. Plants 2021, 10, 1721. [Google Scholar] [CrossRef]
- Kapoor, I.P.S.; Singh, B.; Singh, G.; Isidorov, V.; Szczepaniak, L. Chemistry, antifungal and antioxidant activities of cardamom (Amomum subulatum) essential oil and oleoresins. Int. J. Essent. Oil. Ther. 2008, 2, 29–40. [Google Scholar]
- Scala, A.; Allmann, S.; Mirabella, R.; Haring, M.A.; Schuurink, R.C. Green leaf volatiles: A plant’s multifunctional weapon against herbivores and pathogens. Int. J. Mol. Sci. 2013, 14, 17781–17811. [Google Scholar] [CrossRef] [PubMed]
- Kang, Z.W.; Liu, F.H.; Zhang, Z.F.; Tian, H.G.; Liu, T.X. Volatile β-ocimene can regulate developmental performance of peach aphid Myzus persicae through activation of defense responses in Chinese cabbage Brassica pekinensis. Front. Plant Sci. 2018, 9, 708. [Google Scholar] [CrossRef] [PubMed]
- Pechous, S.W.; Whitaker, B.D. Cloning and functional expression of an (E, E)-α-farnesene synthase cDNA from peel tissue of apple fruit. Planta 2004, 219, 84–94. [Google Scholar] [CrossRef]
- Rodrigues de Queiroz, A.; Hines, C.; Brown, J.; Sahay, S.; Vijayan, J.; Stone, J.M.; Bickford, N.; Wuellner, M.; Glowacka, K.; Buan, N.R.; et al. The effects of exogenously applied antioxidants on plant growth and resilience. Phytochem. Rev. 2023, 1–41. [Google Scholar] [CrossRef]
- Návarová, H.; Bernsdorff, F.; Döring, A.C.; Zeier, J. Pipecolic acid, an endogenous mediator of defense amplification and priming, is a critical regulator of inducible plant immunity. Plant Cell 2012, 24, 5123–5141. [Google Scholar] [CrossRef]
- Mohnike, L.; Huang, W.; Worbs, B.; Feussner, K.; Zhang, Y.; Feussner, I. N-Hydroxy pipecolic acid methyl ester is involved in Arabidopsis immunity. J. Exp. Bot. 2023, 74, 458–471. [Google Scholar] [CrossRef]
- Libik-Konieczny, M.; Kuźniak, E.; Surówka, E.; Ślesak, I.; Nosek, M.; Miszalski, Z. Crassulacean acid metabolism and its role in plant acclimatization to abiotic stresses and defence against pathogens. Prog. Bot. 2020, 81, 277–306. [Google Scholar] [CrossRef]
- Barlass, M.; Miller, R.M.; Antcliff, A.J. Development of methods for screening grapevines for resistance to infection by downy mildew. I. Dual culture in vitro. Am. J. Enol. Vitic. 1986, 37, 61–66. [Google Scholar] [CrossRef]
- Barlass, M.; Miller, R.M.; Douglas, T.J. Development of methods for screening grapevines for resistance to infection by downy mildew. II. Resveratrol production. Am. J. Enol. Vitic. 1987, 38, 65–68. [Google Scholar] [CrossRef]
- Jeandet, P.; Douillet-Breuil, A.C.; Bessis, R.; Debord, S.; Sbaghi, M.; Adrian, M. Phytoalexins from the Vitaceae: Biosynthesis, phytoalexin gene expression in transgenic plants, antifungal activity, and metabolism. J. Agric. Food Chem. 2002, 50, 2731–2741. [Google Scholar] [CrossRef]
- Swaminathan, S.; Reem, N.T.; Lionetti, V.; Zabotina, O.A. Coexpression of fungal cell wall-modifying enzymes reveals their additive impact on Arabidopsis resistance to the fungal pathogen, Botrytis cinerea. Biology 2021, 10, 1070. [Google Scholar] [CrossRef] [PubMed]
- Billard, A.; Fillinger, S.; Leroux, P.; Lachaise, H.; Beffa, R.; Debieu, D. Strong resistance to the fungicide fenhexamid entails a fitness cost in Botrytis cinerea, as shown by comparisons of isogenic strains. Pest Manag. Sci. 2012, 68, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Gao, L.; Chang, P.; Chen, Z.; Zhang, X.; Yin, W.; Fan, Y.; Wang, X. The impact of Elsinoë ampelina infection on key metabolic properties in Vitis vinifera ‘Red Globe’Berries via Multiomics approaches. Mol. Plant-Microbe. Interact. 2022, 35, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Deighton, N.; Muckenschnabel, I.; Colmenares, A.J.; Collado, I.G.; Williamson, B. Botrydial is produced in plant tissues infected by Botrytis cinerea. Phytochemistry 2001, 57, 689–692. [Google Scholar] [CrossRef] [PubMed]
- Schoonbeek, H.; Del Sorbo, G.; De Waard, M.A. The ABC transporter BcatrB affects the sensitivity of Botrytis cinerea to the phytoalexin resveratrol and the fungicide fenpiclonil. Mol. Plant-Microbe Interact. 2001, 14, 562–571. [Google Scholar] [CrossRef]
- Nakaune, R.; Hamamoto, H.; Imada, J.; Akutsu, K.; Hibi, T. A novel ABC transporter gene, PMR5, is involved in multidrug resistance in the phytopathogenic fungus Penicillium digitatum. Mol. Genet. Genomics 2002, 267, 179–185. [Google Scholar] [CrossRef]
- Hayashi, K.; Schoonbeek, H.; De Waard, M.A. Bcmfs1, a novel major facilitator superfamily transporter from Botrytis cinerea, provides tolerance towards the natural toxic compounds camptothecin and cercosporin and towards fungicides. Appl. Environ. Microbiol. 2002, 68, 4996–5004. [Google Scholar] [CrossRef]
- De Waard, M.A.; Andrade, A.C.; Hayashi, K.; Schoonbeek, H.J.; Stergiopoulos, I.; Zwiers, L.H. Impact of fungal drug transporters on fungicide sensitivity, multidrug resistance and virulence. Pest Manage. Sci. 2006, 62, 195–207. [Google Scholar] [CrossRef]
- Shitan, N. Secondary metabolites in plants: Transport and self-tolerance mechanisms. Biosci. Biotechnol. Biochem. 2016, 80, 1283–1293. [Google Scholar] [CrossRef]
- Wang, Z.; Ma, T.; Huang, Y.; Wang, J.; Chen, Y.; Kistler, H.C.; Ma, Z.; Yin, Y. A fungal ABC transporter FgAtm1 regulates iron homeostasis via the transcription factor cascade FgAreA-HapX. PloS Pathog. 2019, 15, e1007791. [Google Scholar] [CrossRef]
- Zwiers, L.H.; De Waard, M.A. Characterization of the ABC transporter genes MgAtr1 and MgAtr2 from the wheat pathogen Mycosphaerella graminicola. Fungal Genet. Biol. 2000, 30, 115–125. [Google Scholar] [CrossRef] [PubMed]



 ), noninoculation at 6 dpi (B
), noninoculation at 6 dpi (B  ), noninoculation at 9 dpi (C
), noninoculation at 9 dpi (C  ), inoculation at 3 dpi (D
), inoculation at 3 dpi (D  ), inoculation at 6 dpi (E
), inoculation at 6 dpi (E  ), and inoculation at 9 dpi (F
), and inoculation at 9 dpi (F  ).
), noninoculation at 6 dpi (B ), noninoculation at 9 dpi (C ), inoculation at 3 dpi (D ), inoculation at 6 dpi (E ), and inoculation at 9 dpi (F ).
).
), noninoculation at 6 dpi (B ), noninoculation at 9 dpi (C ), inoculation at 3 dpi (D ), inoculation at 6 dpi (E ), and inoculation at 9 dpi (F ).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence |
|---|---|
| BcActin | Forward (F): TCCAAGCGTGGTATTCTTACCC |
| Reverse (R): TGGTGCTACACGAAGTTCGTTG | |
| BcatrB | F: TCTAACCCCGCTGAACACAT |
| R: AGAGAGGGGTTGCGAATTCA | |
| BcatrD | F: TCCAGGAGCCAGCAATACAA |
| R: AACCCTGCGGCAAATGAATT | |
| BcatrK | F: CCGCTTTGATGGAGAACGAG |
| R: GTGATGTAGTCGCCACCAAC |
| Compound Name | Retention Time | Formula | Adduct | m/z | Mass Error | MS/MS Spectrum | FC (VB5 vs. VNI) * | FC (VB2 vs. VNI) | FC (SB5 vs. SNI) | FC (SB2 vs. SNI) | FC (SNI vs. VNI) | MS/MS Reference | Concentration (µg/mL) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Flavonoids | |||||||||||||
| Kaempferol-7-O-glucoside | 4.97 | C21H20O11 | [M + H]− | 447.0967 | 8.53 | 447, 121, 149, 315 | 0.75 | 0.70 | 2.82 | 3.03 | 0.29 | Database | |
| Catechin | 5.69 | C15H14O6 | [M + H]+ | 291.8177 | −2.19 | 139, 207, 123, 147 | 1.88 | 1.87 | 1.53 | 4.89 | 1.25 | Database | |
| Flavanone | 6.08 | C15H12O8 | [M + H]+ | 321.0603 | −1.09 | 153, 275, 149, 139 | 2.68 | 3.43 | 1.16 | 7.28 | 2.23 | Standards | 2.488–6.123 |
| Quercetin-3β-D-glucoside | 7.53 | C21H20O12 | [M + H]− | 463.0891 | 1.81 | 300, 301, 463, 271 | 1.83 | 1.33 | 0.74 | 2.00 | 0.59 | Database | |
| Naringenin | 7.54 | C15H12O5 | [M + H]+ | 273.0681 | −1.33 | - | 3.94 | 1.29 | 0.53 | 0.65 | 5.97 | [27] | |
| Kaempferol-3-glucuronide | 8.66 | C21H18O12 | [M + H]− | 461.2493 | 1.34 | 285, 461, 113, 229 | 0.88 | 0.88 | 3.65 | 5.49 | 0.28 | Database | |
| Afzelin | 9.01 | C21H20O10 | [M + H]− | 431.1926 | 1.1 | 269, 285, 431, 59 | 2.07 | 2.06 | 2.12 | 2.50 | 4.61 | Database | |
| Amino Acids | |||||||||||||
| Asparagine | 0.87 | C4H8N2O3 | [M + H]− | 131.0454 | −5.68 | 114, 113, 131, 95 | 0.75 | 0.71 | 0.87 | 0.89 | 0.48 | Database | |
| Aspartic Acid | 0.89 | C4H7NO4 | [M + H]− | 132.0294 | −5.86 | 88, 132, 115, 71 | 0.31 | 0.27 | 1.59 | 1.43 | 0.73 | Database | |
| Phenylalanine | 4.35 | C9H11NO2 | [M + H]+ | 166.0861 | −0.83 | 166, 120, 103, 149 | 0.13 | 0.18 | 1.07 | 3.91 | 0.60 | Database | |
| Phenolic Acids and Derivatives | |||||||||||||
| Rhusflavanone | 1.00 | C30H22O10 | [M + H]+ | 543.1326 | 5.34 | - | 1.91 | 1.83 | 1.06 | 1.11 | 1.45 | [28] | |
| Salicylic Acid | 5.63 | C7H6O3 | [M + H]− | 137.0257 | −5.39 | - | 1.39 | 1.64 | 1.98 | 0.91 | 1.48 | Standards | 0.026–0.0451 |
| Trans-Piceid | 5.81 | C20H22O8 | [M + H]+ | 391.1317 | −1.67 | - | 1.62 | 1.63 | 1.27 | 1.44 | 1.51 | [29] | |
| Psoralidin | 6.58 | C20H16O5 | [M + H]+ | 337.1067 | −0.83 | 337, 275, 235, 245 | 1.00 | 1.11 | 1.43 | 2.29 | 1.83 | Database | |
| Reaveratroloside | 7.60 | C20H22O9 | [M + H]− | 405.213 | 1.03 | 243, 405, 361, 375 | 3.35 | 2.04 | 1.95 | 1.52 | 1.65 | Database | |
| Dihydroresveratrol | 7.61 | C14H14O3 | [M + H]− | 229.094 | 0.01 | - | 0.62 | 0.65 | 1.07 | 1.03 | 1.10 | [30] | |
| Resveratrol | 8.23 | C14H12O3 | [M + H]− | 227.0712 | −0.45 | 227, 185, 202, 164 | 13.72 | 2.68 | 8.33 | 3.63 | 0.56 | Standards | 0.005–0.045 |
| Cis-Resveratrol | 8.23 | C14H12O3 | [M + H]+ | 229.086 | −1.26 | - | 5.31 | 2.96 | 2.06 | 3.98 | 1.26 | [29] | |
| Astragalin | 8.57 | C21H20O11 | [M + H]− | 447.1879 | 0.82 | 447, 284, 255, 300 | 0.74 | 0.68 | 2.39 | 2.80 | 0.34 | Database | |
| Isorhamnetin | 8.65 | C16H12O7 | [M + H]+ | 317.0651 | −1.37 | 317, 285, 274, 257 | 0.90 | 0.64 | 0.88 | 4.39 | 0.77 | Database | |
| Eugenol | 9.09 | C10H12O2 | [M + H]− | 163.0243 | −3.29 | 148, 119, 163, 59 | 1.21 | 1.46 | 1.69 | 2.11 | 1.32 | Standards | 0.301–0.396 |
| Organic Acids | |||||||||||||
| Malic Acid | 1.23 | C4H6O5 | [M + H]− | 133.039 | −6.27 | 115, 133, 71, 72 | 1.15 | 1.15 | 1.12 | 1.20 | 0.92 | Database | |
| Ascorbic Acid | 1.34 | C6H8O6 | [M + H]− | 175.0623 | −3.51 | 115, 175, 87, 71 | 1.08 | 1.04 | 1.35 | 1.66 | 0.64 | Database | |
| Pipecolic Acid | 1.47 | C6H11NO2 | [M + H]+ | 130.0863 | 0.14 | 130, 84, 85, 67 | 0.55 | 0.45 | 1.29 | 2.16 | 0.71 | Database | |
| 2-Isopropylmalic Acid | 5.91 | C7H12O5 | [M + H]− | 175.0206 | −2.96 | 175, 115, 85, 113 | 0.63 | 0.67 | 2.49 | 1.52 | 0.46 | Database | |
| Epicatechin | 6.50 | C15H14O6 | [M + H]+ | 291.1752 | −2.18 | 139, 123, 147, 165 | 2.28 | 1.64 | 1.65 | 6.34 | 0.98 | Database | |
| Ellagic Acid | 8.43 | C14H6O8 | [M + H]− | 300.8998 | 0.11 | 219, 208, 200, 126 | 0.87 | 0.90 | 1.65 | 1.17 | 1.06 | Database | |
| Jasmonic Acid | 9.25 | C12H18O3 | [M + H]− | 209.1182 | −0.74 | 59, 209, 133, 173 | 0.63 | 1.27 | 1.52 | 1.45 | 1.26 | Database | |
| Oleanolic Acid | 12.62 | C30H48O3 | [M + H]− | 455.3537 | 1.28 | 455, 122, 130, 148 | 0.91 | 0.64 | 0.80 | 0.72 | 1.42 | Database | |
| Abietic Acid | 12.64 | C20H30O2 | [M + H]− | 301.0357 | 1.76 | 301, 151, 229, 271 | 0.96 | 1.11 | 0.92 | 1.00 | 1.25 | Database | |
| Sugars | |||||||||||||
| Glucose 1-phosphate | 0.92 | C6H13O9P | [M + H]+ | 261.0369 | −0.71 | 98, 85, 97, 127 | 1.15 | 1.29 | 1.11 | 5.27 | 1.05 | Database | |
| Alcohols | |||||||||||||
| Piceatannol | 7.62 | C14H12O4 | [M + H]− | 243.0664 | 0.99 | 243, 109, 198, 215 | 1.04 | 1.21 | 1.38 | 1.00 | 1.06 | Database | |
| Alkaloids | |||||||||||||
| Berberine | 8.03 | C20H17NO4 | [M + H]+ | 336.2181 | −1.06 | - | 1.74 | 1.39 | 0.44 | 0.43 | 6.28 | Standards | |
| Reserpine | 9.28 | C33H40N2O9 | [M + H]+ | 609.2731 | −0.43 | - | 1.13 | 0.94 | 0.93 | 0.16 | 5.74 | Standards | 1.125–1.248 |
| Others | |||||||||||||
| Stearamide | 12.98 | C18H37NO | [M + H]+ | 284.2943 | −1.99 | 284, 60, 88, 102 | 1.15 | 1.06 | 0.86 | 1.11 | 1.17 | Database | |
| Days After Inoculation | Compound | Time | M/Z | Log2FC | Regulation |
|---|---|---|---|---|---|
| 3 | Heptane | 12.5 | 57.0703 | 2.2 | Up |
| 2,2,4,4-Tetramethyloctane | 13.8 | 57.0701 | 1.9 | Up | |
| β-Ocimene | 14.6 | 145.9681 | 1.4 | Up | |
| Octadecane | 17.7 | 57.0701 | 17.9 | Up | |
| 2,5-Dihydroxybenzaldehyde | 18.1 | 266.9993 | −0.6 | Down | |
| Cyclohexane | 18.3 | 69.0701 | 18.4 | Up | |
| Decane | 18.5 | 57.0699 | 17.9 | Up | |
| Tetradecane | 19.3 | 57.0703 | 0.3 | Up | |
| 6,7-Dimethyl-1,2,3,5,8,8a-hexahydronaphthalene | 20.6 | 91.0543 | 19.5 | Up | |
| Eicosane | 20.6 | 57.0703 | 0.6 | Up | |
| Tricosanol | 21.0 | 55.0547 | 0.5 | Up | |
| 6 | 6,7-Dimethyl-1,2,3,5,8,8a-hexahydronaphthalene | 20.5 | 91.0540 | −0.1 | Down |
| Ylangene | 24.1 | 105.0692 | 1.7 | Up | |
| 7-epi-Silphiperfol-5-ene | 24.2 | 175.1478 | 20.8 | Up | |
| 2-Piperidinone | 24.5 | 57.7010 | 1.6 | Up | |
| Germacrene D | 26.9 | 161.1326 | 1.2 | Up | |
| Butylated Hydroxytoluene | 27.7 | 205.1583 | 0.9 | Up | |
| 9 | Trans-β-Ocimene | 14.3 | 145.9686 | 0.9 | Up |
| β-Ocimene | 14.6 | 145.9681 | 0.9 | Up | |
| 2,4,6-Octatriene | 17.2 | 121.1007 | 1.5 | Up | |
| 1,3-Cyclohexadiene | 17.6 | 121.1022 | 1.0 | Up | |
| Sulfurous Acid | 21.5 | 57.0697 | 0.2 | Up | |
| Nonadecane | 22.2 | 57.0697 | −0.3 | Down | |
| n-Tridecan-1-ol | 22.4 | 69.0697 | −0.1 | Down | |
| 1-Iodo-2-methylundecane | 22.7 | 57.0697 | −0.1 | Down | |
| 1-Octanol | 22.8 | 69.0700 | 0.9 | Up | |
| Hexadecane | 22.9 | 57.0697 | −0.1 | Down | |
| β-Guaiene | 23.1 | 119.0856 | 0.3 | Up | |
| Ylangene | 24.2 | 105.0697 | 22.5 | Up | |
| 7-epi-Silphiperfol-5-ene | 24.3 | 175.1478 | 22.5 | Up | |
| 2-Piperidinone | 24.5 | 57.0700 | 2.2 | Up | |
| Cis-13-Eicosenoic Acid | 24.7 | 57.0697 | 21.2 | Up | |
| Caryophyllene | 25.4 | 105.0697 | 22.5 | Up | |
| γ-Elemene | 25.8 | 121.1004 | 20.8 | Up | |
| isoledene | 26.0 | 105.0698 | 21.5 | Up | |
| Cedrene | 26.1 | 57.0700 | 21.0 | Up | |
| cis-Muurola-4 | 26.5 | 161.1326 | 21.6 | Up | |
| γ-Muurolene | 26.8 | 161.1326 | 21.8 | Up | |
| Germacrene D | 27.0 | 161.1326 | 23.9 | Up | |
| Octadecane | 27.2 | 57.0700 | 1.1 | Up | |
| α-Farnesene | 27.6 | 57.0709 | 0.6 | Up | |
| Butylated Hydroxytoluene | 27.7 | 205.1583 | 0.6 | Up | |
| β-copaene | 27.8 | 159.1168 | 21.7 | Up | |
| 1-Hexadecanol | 27.9 | 69.0697 | −19.9 | Down | |
| Naphthalene | 28.0 | 159.1162 | 2.3 | Up | |
| 1H-3a,7-Methanoazulene | 29.7 | 57.0599 | 1.1 | Up |
| Pathway Name | Match Status a | P b | Holm p c | Impact d |
|---|---|---|---|---|
| Galactose Metabolism | 5/27 | 0.003 | 0.263 | 0.272 |
| Aminoacyl-tRNA Biosynthesis | 6/46 | 0.006 | 0.610 | 0.000 |
| Butanoate Metabolism | 3/17 | 0.024 | 1.000 | 0.136 |
| Pentose Phosphate Pathway | 3/19 | 0.032 | 1.000 | 0.116 |
| Valine, Leucine, and Isoleucine Biosynthesis | 3/22 | 0.047 | 1.000 | 0.000 |
| Alanine, Aspartate, and Glutamate Metabolism | 3/22 | 0.047 | 1.000 | 0.255 |
| Starch and Sucrose Metabolism | 2/22 | 0.200 | 1.000 | 0.099 |
| Isoquinoline Alkaloid Biosynthesis | 1/6 | 0.207 | 1.000 | 0.000 |
| Phenylpropanoid Biosynthesis | 3/46 | 0.250 | 1.000 | 0.100 |
| Monobactam Biosynthesis | 1/8 | 0.266 | 1.000 | 0.000 |
| Tropane, Piperidine, and Pyridine Alkaloid Biosynthesis | 1/8 | 0.266 | 1.000 | 0.000 |
| Lysine Biosynthesis | 1/9 | 0.293 | 1.000 | 0.000 |
| Cyanoamino Acid Metabolism | 2/29 | 0.300 | 1.000 | 0.000 |
| Phenylalanine Metabolism | 1/11 | 0.346 | 1.000 | 0.471 |
| Glycine, Serine, and Threonine Metabolism | 2/33 | 0.357 | 1.000 | 0.120 |
| Arginine and Proline Metabolism | 2/34 | 0.370 | 1.000 | 0.066 |
| Nicotinate and Nicotinamide Metabolism | 1/13 | 0.395 | 1.000 | 0.000 |
| Valine, Leucine, and Isoleucine Degradation | 2/37 | 0.411 | 1.000 | 0.000 |
| Sulfur Metabolism | 1/15 | 0.440 | 1.000 | 0.033 |
| Glucosinolate Biosynthesis | 3/65 | 0.449 | 1.000 | 0.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Gao, T.; Liang, Z.; Hao, J.; Liu, P.; Liu, X. Dynamic Changes in Plant Secondary Metabolites Induced by Botrytis cinerea Infection. Metabolites 2023, 13, 654. https://doi.org/10.3390/metabo13050654
Wu Z, Gao T, Liang Z, Hao J, Liu P, Liu X. Dynamic Changes in Plant Secondary Metabolites Induced by Botrytis cinerea Infection. Metabolites. 2023; 13(5):654. https://doi.org/10.3390/metabo13050654
Chicago/Turabian StyleWu, Zhaochen, Tuqiang Gao, Zhengya Liang, Jianjun Hao, Pengfei Liu, and Xili Liu. 2023. "Dynamic Changes in Plant Secondary Metabolites Induced by Botrytis cinerea Infection" Metabolites 13, no. 5: 654. https://doi.org/10.3390/metabo13050654
APA StyleWu, Z., Gao, T., Liang, Z., Hao, J., Liu, P., & Liu, X. (2023). Dynamic Changes in Plant Secondary Metabolites Induced by Botrytis cinerea Infection. Metabolites, 13(5), 654. https://doi.org/10.3390/metabo13050654