

The Relationship between Maternal Antibodies to Fetal Brain and Prenatal Stress Exposure in Autism Spectrum Disorder

 , , and
, , and
Abstract
:1. Introduction
2. Methods
3. Statistical Analyses
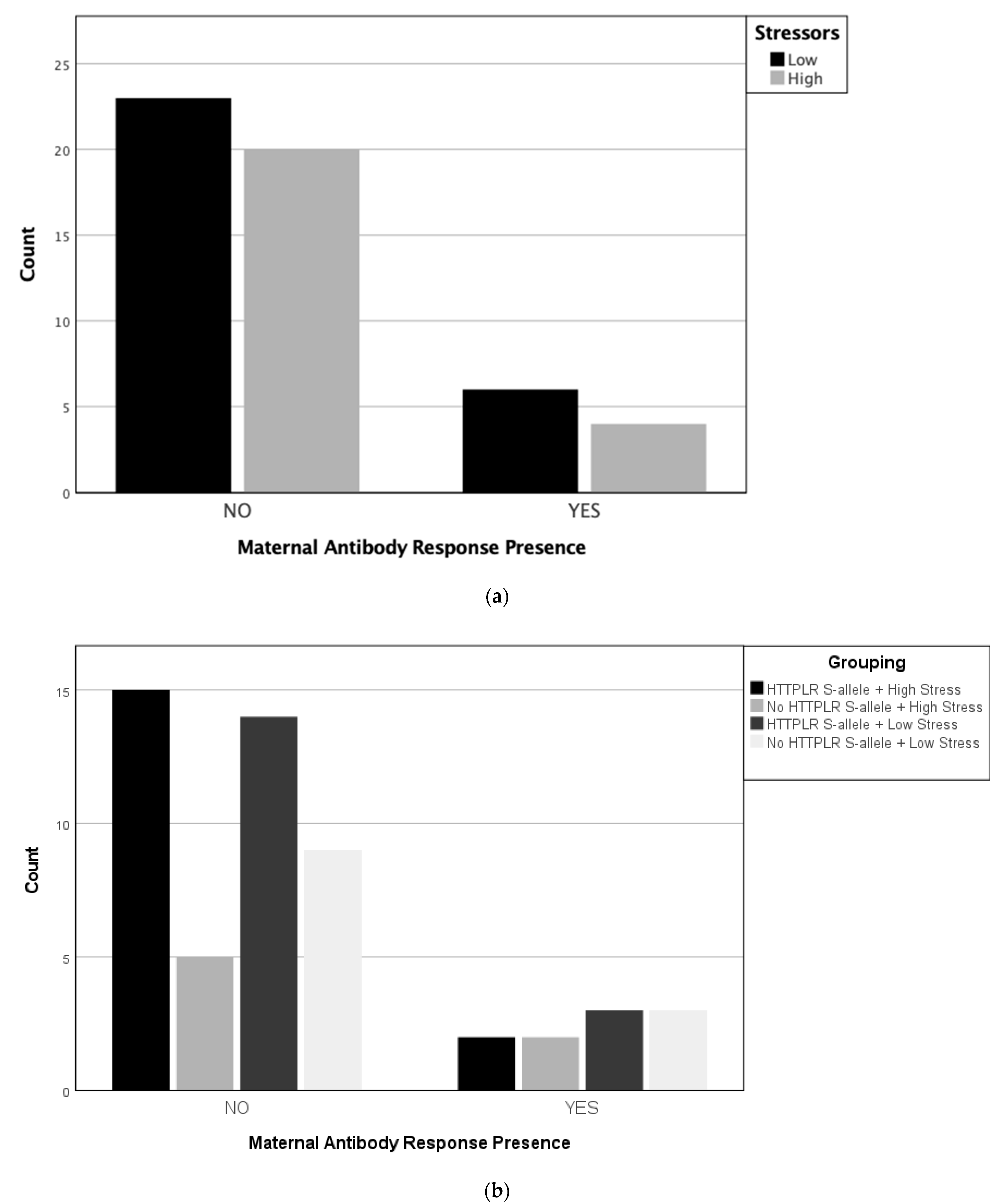
4. Results
Participant Characteristics
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gage, S.H.; Munafò, M.R.; Davey Smith, G. Causal inference in developmental origins of health and disease (DOHaD) research. Annu. Rev. Psychol. 2016, 67, 567–585. [Google Scholar] [CrossRef] [PubMed]
- Hallmayer, J.; Cleveland, S.; Torres, A.; Phillips, J.; Cohen, B.; Torigoe, T.; Miller, J.; Fedele, A.; Collins, J.; Smith, K.; et al. Genetic heritability and shared environmental factors among twin pairs with autism. Arch. Gen. Psychiatry 2011, 68, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Folstein, S.; Rutter, M. Infantile autism: A genetic study of 21 twin pairs. J. Child Psychol. Psychiatry 1977, 18, 297–321. [Google Scholar] [CrossRef]
- Trottier, G.; Srivastava, L.; Walker, C.-D. Etiology of infantile autism: A review of recent advances in genetic and neurobiological research. J. Psychiatry Neurosci. 1999, 24, 103. [Google Scholar] [PubMed]
- Abbott, P.W.; Gumusoglu, S.B.; Bittle, J.; Beversdorf, D.Q.; Stevens, H.E. Prenatal stress and genetic risk: How prenatal stress interacts with genetics to alter risk for psychiatric illness. Psychoneuroendocrinology 2018, 90, 9–21. [Google Scholar] [CrossRef]
- Niederhofer, H.; Reiter, A. Maternal stress during pregnancy, its objectivation by ultrasound observation of fetal intrauterine movements and child’s temperament at 6 months and 6 years of age: A pilot study. Psychol. Rep. 2000, 86, 526–528. [Google Scholar] [CrossRef]
- Van Os, J.; Selten, J.-P. Prenatal exposure to maternal stress and subsequent schizophrenia: The May 1940 invasion of the Netherlands. Br. J. Psychiatry 1998, 172, 324–326. [Google Scholar] [CrossRef]
- Beversdorf, D.Q.; Manning, S.; Hillier, A.; Anderson, S.; Nordgren, R.; Walters, S.; Nagaraja, H.N.; Cooley, W.C.; Gaelic, S.E.; Bauman, M. Timing of prenatal stressors and autism. J. Autism Dev. Disord. 2005, 35, 471–478. [Google Scholar] [CrossRef]
- Roberts, A.L.; Lyall, K.; Rich-Edwards, J.W.; Ascherio, A.; Weisskopf, M.G. Maternal exposure to childhood abuse is associated with elevated risk of autism. JAMA Psychiatry 2013, 70, 508. [Google Scholar] [CrossRef]
- Beversdorf, D.Q.; Stevens, H.E.; Jones, K.L. Prenatal stress, maternal immune dysregulation, and their association with autism spectrum disorders. Curr. Psychiatry Rep. 2018, 20, 76. [Google Scholar] [CrossRef]
- Beversdorf, D.Q.; Stevens, H.E.; Margolis, K.G.; Van de Water, J. Prenatal stress and maternal immune dysregulation in autism spectrum disorders: Potential points for intervention. Curr. Pharm. Des. 2019, 25, 4331–4343. [Google Scholar] [CrossRef]
- Li, J.; Vestergaard, M.; Obel, C.; Christensen, J.; Precht, D.H.; Lue, M.; Olsen, J. A nationwide study on the risk of autism after prenatal stress exposure to maternal bereavement. Pediatrics 2009, 123, 1102–1107. [Google Scholar] [CrossRef] [PubMed]
- Larsson, J.H.; Eaton, W.W.; Madsen, K.M.; Vestergaard, M.; Olesen, A.V.; Agerbo, E.; Schendel, D.; Thorsen, P.; Mortensen, P.B. Risk factors for autism: Perinatal factors, parental psychiatric history, and socioeconomic status. Am. J. Epidemiol. 2005, 161, 916–925. [Google Scholar] [CrossRef] [PubMed]
- Class, Q.A.; Abel, K.M.; Khashan, A.S.; Ricket, M.E.; Larsson, H.; Hultman, C.M.; Långström, N.; Lichtenstein, P.; D’Onofrio, B.M. Offspring psychopathology following preconception, prenatal and postnatal maternal bereavement stress. Psychol. Med. 2014, 44, 71–84. [Google Scholar] [CrossRef]
- Manzari, N.; Matvienko-Sikar, K.; Baldoni, F.; O’Keefe, G.W.; Khashan, A.S. Prenatal maternal stress and risk of neurodevelopmental disorders in the offspring: A systematic review and meta-analysis. Soc. Psychiatry Psychiatr. Epidemiol. 2019, 54, 1299–1309. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-J.; Strodl, E.; Wu, C.-A.; Chen, J.-Y.; Huang, L.-H.; Yin, X.-N.; Wen, G.-M.; Sun, D.-K.; Xian, D.-X.; Li, C.-G.; et al. Prenatal maternal stress and autistic-like behaviors in Chinese preschoolers. Stress Health 2021, 37, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Kinney, D.K.; Miller, A.M.; Crowley, D.J.; Huang, E.; Gerber, E. Autism prevalence following prenatal exposure to hurricanes and tropical storms in Louisiana. J. Autism Dev. Disord. 2008, 38, 481–488. [Google Scholar] [CrossRef]
- Murphy, D.L.; Lerner, A.; Rudnick, G.; Lesch, K.-P. Serotonin transporter: Gene, genetic disorders, and pharmacogenetics. Mol. Interv. 2004, 4, 109. [Google Scholar] [CrossRef] [PubMed]
- Bondy, B.; Buettner, A.; Zill, P. Genetics of suicide. Mol. Psychiatry 2006, 11, 336–351. [Google Scholar] [CrossRef]
- Feinn, R.; Nellissery, M.; Kranzler, H.R. Meta-analysis of the association of a functional serotonin transporter promoter polymorphism with alcohol dependence. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2005, 133, 79–84. [Google Scholar] [CrossRef]
- Miozzo, R.; Eaton, W.W.; Bienvenu, O.J., 3rd; Samuels, J.; Nestadt, G. The serotonin transporter gene polymporhism (SLC6A4) and risk for psychiatric morbidity and comorbidity in the Baltimore ECA follow-up study. Compr. Psychiatry 2020, 102, 152199. [Google Scholar] [CrossRef] [PubMed]
- Hecht, P.M.; Hudson, M.; Connors, S.L.; Tilley, M.R.; Liu, X.; Beversdorf, D.Q. Maternal serotonin transporter genotype affects risk for ASD with exposure to prenatal stress. Autism Res. 2016, 9, 1151–1160. [Google Scholar] [CrossRef]
- Kiecolt-Glaser, J.K.; Belury, M.A.; Porter, K.; Beversdorf, D.Q.; Lemeshow, S.; Glaser, R. Depressive Symptoms, omega-6:omega-3 Fatty Acids, and Inflammation in Older Adults. Psychosom. Med. 2007, 69, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Morey, J.N.; Boggero, I.A.; Scott, A.B.; Segerstrom, S.C. Current directions in stress and human immune function. Curr. Opin. Psychol. 2015, 5, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.L.; Pride, M.C.; Edmiston, E.; Yang, M.; Silverman, J.L.; Crawley, J.N.; Van de Water, J. Autism-specific maternal autoantibodies produce behavioral abnormalities in an endogenous antigen-driven mouse model of autism. Mol. Psychiatry 2020, 25, 2994–3009. [Google Scholar] [CrossRef]
- Lee, B.K.; Magnusson, C.; Gardner, R.M.; Blomström, Å.; Newschaffer, C.J.; Burstyn, I.; Karlsson, H.; Dalman, C. Maternal hospitalization with infection during pregnancy and risk of autism spectrum disorders. Brain Behav. Immun. 2015, 44, 100–105. [Google Scholar] [CrossRef]
- Chaouat, G. The Th1/Th2 paradigm: Still important in pregnancy? Semin. Immunopathol. 2007, 29, 95–113. [Google Scholar] [CrossRef]
- Wegmann, T.G.; Lin, H.; Guilbert, L.; Mosmann, T.R. Bidirectional cytokine interactions in the maternal-fetal relationship: Is successful pregnancy a TH2 phenomenon? Immunol. Today 1993, 14, 353–356. [Google Scholar] [CrossRef]
- Antoun, S.; Ellul, P.; Peyre, H.; Rosenzwajg, M.; Gressens, P.; Klatzmann, D.; Delorme, R. Fever during pregnancy as a risk factor for neurodevelopmental disorders: Results from a systematic review and meta-analysis. Mol. Autism 2021, 12, 60. [Google Scholar] [CrossRef]
- Atladóttir, H.Ó.; Henriksen, T.B.; Schendel, D.E.; Parner, E.T. Autism after infection, febrile episodes, and antibiotic use during pregnancy: An exploratory study. Pediatrics 2012, 130, e1447–e1454. [Google Scholar] [CrossRef]
- Zerbo, O.; Iosif, A.-M.; Walker, C.; Ozonoff, S.; Hansen, R.L.; Hertz-Picciotto, I. Is maternal influenza or fever during pregnancy associated with autism or developmental delays? Results from the CHARGE (CHildhood Autism Risks from Genetics and Environment) study. J. Autism Dev. Disord. 2013, 43, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Careaga, M.; Murai, T.; Bauman, M.D. Maternal Immune Activation and Autism Spectrum Disorder: From Rodents to Nonhuman and Human Primates. Biol. Psychiatry 2017, 81, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Goines, P.E.; Croen, L.A.; Braunschweig, D.; Yoshida, C.K.; Grether, J.; Hansen, R.; Kharrazi, M.; Ashwood, P.; Van De Water, J. Increased midgestational IFN-γ, IL-4 and IL-5 in women bearing a child with autism: A case-control study. Mol. Autism 2011, 2, 13. [Google Scholar] [CrossRef] [PubMed]
- Goines, P.; Van de Water, J. The immune system’s role in the biology of autism. Curr. Opin. Neurol. 2010, 23, 111. [Google Scholar] [CrossRef]
- Jones, K.L.; Croen, L.A.; Yoshida, C.K.; Heuer, L.; Hansen, R.; Zerbo, O.; DeLorenze, G.N.; Kharrazi, M.; Yolken, R.; Ashwood, P.; et al. Autism with intellectual disability is associated with increased levels of maternal cytokines and chemokines during gestation. Mol. Psychiatry 2017, 22, 273–279. [Google Scholar] [CrossRef]
- Currenti, S.A. Understanding and Determining the Etiology of Autism. Cell. Mol. Neurobiol. 2010, 30, 161–171. [Google Scholar] [CrossRef]
- Braunschweig, D.; Van de Water, J. Maternal autoantibodies in autism. Arch. Neurol. 2012, 69, 693–699. [Google Scholar] [CrossRef]
- Jones, K.L.; Van de Water, J. Maternal autoantibody related autism: Mechanisms and pathways. Mol. Psychiatry 2019, 24, 252–265. [Google Scholar] [CrossRef]
- Braunschweig, D.; Ashwood, P.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Croen, L.A.; Pessah, I.N.; Van de Water, J. Autism: Maternally derived antibodies specific for fetal brain proteins. Neutotoxicology 2008, 29, 226–231. [Google Scholar] [CrossRef]
- Brimberg, L.; Sadiq, A.; Gregersen, P.; Diamond, B. Brain-reactive IgG correlates with autoimmunity in mothers of a child with an autism spectrum disorder. Mol. Psychiatry 2013, 18, 1171–1177. [Google Scholar] [CrossRef]
- Singer, H.S.; Morris, C.M.; Gause, C.D.; Gillin, P.K.; Crawford, S.; Zimmerman, A.W. Antibodies against fetal brain in sera of mothers with autistic children. J. Neuroimmunol. 2008, 194, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Braunschweig, D.; Krakowiak, P.; Duncanson, P.; Boyce, R.; Hansen, R.L.; Ashwood, P.; Hertz-Picciotto, I.; Pessah, I.N.; Van de Water, J. Autism-specific maternal autoantibodies recognize critical proteins in developing brain. Transl. Psychiatry 2013, 3, e277. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Celis, A.; Croen, L.A.; Yoshida, C.K.; Alexeeff, S.E.; Schauer, J.; Yolken, R.H.; Ashwood, P.; Van de Water, J. Maternal autoantibody profiles as biomarkers for ASD and ASD with co-occurring intellectual disability. Mol. Psychiatry 2022, 27, 3760–3767. [Google Scholar] [CrossRef]
- Ramirez-Celis, A.; Edmiston, E.; Schauer, J.; Vu, T.; Van De Water, J. Peptides of neuron specific enolase as potential ASD biomarkers: From discovery to epitope mapping. Brain Behav. Immun. 2020, 84, 200–208. [Google Scholar] [CrossRef]
- Lord, C.; Rutter, M.; Goode, S.; Heemsbergen, J.; Jordan, H.; Mawhood, L.; Schopler, E. Austism diagnostic observation schedule: A standardized observation of communicative and social behavior. J. Autism Dev. Disord. 1989, 19, 185–212. [Google Scholar] [CrossRef]
- Lord, C.; Rutter, M.; Le Couteur, A. Autism Diagnostic Interview-Revised: A revised version of a diagnostic interview for caregivers of individuals with possible pervasive developmental disorders. J. Autism Dev. Disord. 1994, 24, 659–685. [Google Scholar] [CrossRef] [PubMed]
- Holmes, T.H.; Rahe, R.H. The social readjustment rating scale. J. Psychosom. Res. 1967, 11, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Beversdorf, D.Q.; Shah, A.; Jhin, A.; Noel-MacDonnell, J.; Hecht, P.; Ferguson, B.J.; Bruce, D.; Tilley, M.; Talebizadeh, Z. microRNAs and Gene–Environment Interactions in Autism: Effects of Prenatal Maternal Stress and the SERT Gene on Maternal microRNA Expression [Brief Research Report]. Front. Psychiatry 2021, 12, 668577. [Google Scholar] [CrossRef]
- Baganz, N.L.; Blakely, R.D. A Dialogue between the Immune System and Brain, Spoken in the Language of Serotonin. ACS Chem. Neurosci. 2013, 4, 48–63. [Google Scholar] [CrossRef]
- Bruce, M.R.; Couch, A.C.M.; Grant, S.; McLellan, J.; Ku, K.; Change, C.; Bachman, A.; Matson, M.; Berman, R.F.; Maddock, R.J.; et al. Altered behavior, brain structure, and neurometabolites in a rat model of autism-specific maternal autoantibody exposure. Mol. Psychiatry 2023. [Google Scholar] [CrossRef]
- Mazón-Cabrera, R.; Liesenborgs, J.; Brône, B.; Vandormael, P.; Somers, V. Novel maternal autoantibodies in autism spectrum disorder: Implications for screening and diagnosis. Front. Neurosci. 2023, 17, 1067833. [Google Scholar] [CrossRef]
- Noskova, T.; Pivac, N.; Nedic, G.; Kazantseva, A.; Gaysina, D.; Faskhutdinova, G.; Gareeva, A.; Khalilova, Z.; Khusnutdinova, E.; Kovacic, D.K.; et al. Ethnic differences in the serotonin transporter polymorphism (5-HTTLPR) in several European populations. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 1735–1739. [Google Scholar] [CrossRef]
- Bolton, J.L.; Marinero, S.; Hassanzadeh, T.; Natesan, D.; Le, D.; Belliveau, C.; Mason, S.N.; Auten, R.L.; Bilbo, S.D. Gestational Ex posure to Air Pollution Alters Cortical Volume, Microglial Morphology, and Microglia-Neuron Interactions in a Sex-Specific Manner. Front. Synaptic Neurosci. 2017, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Volk, H.E.; Kerin, T.; Lurmann, F.; Hertz-Picciotto, I.; McConnell, R.; Campbell, D.B. Autism spectrum disorder: Interaction of air pollution with the MET receptor tyrosine kinase gene. Epidemiology 2014, 25, 44–47. [Google Scholar] [CrossRef] [PubMed]
- von Ehrenstein, O.S.; Aralis, H.; Cockburn, M.; Ritz, B. In utero exposure to toxic air pollutants and risk of childhood autism. Epidemiology 2014, 25, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Raz, R.; Roberts, A.L.; Lyall, K.; Hart, J.E.; Just, A.C.; Laden, F.; Weisskopf, M.G. Autism spectrum disorder and particulate matter air pollution before, during, and after pregnancy: A nested case-control analysis within the Nurses’ Health Study II Cohort. Environ. Health Perspect. 2015, 123, 264–270. [Google Scholar] [CrossRef]
- Kalkbrenner, A.E.; Windham, G.C.; Serre, M.L.; Akita, Y.; Wang, X.; Hoffman, K.; Thayer, B.P.; Daniels, J.L. Particulate matter exposure, prenatal and postnatal windows of susceptibility, and autism spectrum disorders. Epidemiology 2015, 26, 30–42. [Google Scholar] [CrossRef]
- Ornoy, A. Valproic acid in pregnancy: How much are we endangering the embryo and fetus? Reprod. Toxicol. 2009, 28, 1–10. [Google Scholar] [CrossRef]
- Connors, S.L.; Crowell, D.E.; Eberhart, C.G.; Copeland, J.; Newschaffer, C.J.; Spence, S.J.; Zimmerman, A.W. β2-adrenergic receptor activation and genetic polymorphisms in autism: Data from dizygotic twins. J. Child Neurol. 2005, 20, 876–884. [Google Scholar] [CrossRef]
- Lyall, K.; Schmidt, R.J.; Hertz-Picciotto, I. Maternal lifestyle and environmental risk factors for autism spectrum disorders. Int. J. Epidemiol. 2014, 43, 443–464. [Google Scholar] [CrossRef]
- Rossignol, D.A.; Genuis, S.J.; Frye, R.E. Environmental toxicants and autism spectrum disorders: A systematic review. Transl. Psychiatry 2014, 4, e360. [Google Scholar] [CrossRef] [PubMed]
- Surén, P.; Roth, C.; Bresnahan, M.; Haugen, M.; Hornig, M.; Hirtz, D.; Lie, K.; Lipkin, I.; Magnus, P.; Reichborn-Kjennerud, T.; et al. Association between maternal use of folic acid supplements and risk of autism spectrum disorders in children. J. Am. Med. Assoc. 2013, 309, 570–577. [Google Scholar] [CrossRef] [PubMed]
- D’Onofrio, B.M.; Rickert, M.E.; Frans, E.; Kuja-Halkola, R.; Almqvist, C.; Sjölander, A.; Larsson, H.; Lichtenstein, P. Paternal age at childbearing and offspring psychiatric and academic morbidity. JAMA Psychiatry 2014, 71, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Cheslack-Postava, K.; Suominen, A.; Jokiranta, E.; Lehti, V.; McKeague, I.W.; Sourander, A.; Brown, A.S. Increased risk of autism spectrum disorders at short and long interpregnancy intervals. Child Adolesc. Psychiatry 2014, 53, 1074–1081. [Google Scholar] [CrossRef]


{kind=link}
| Prenatal Stress | 5-HTTLPR Genotype | N |
|---|---|---|
| High | S/S + S/L | 17 |
| LONG | 7 | |
| Low | S/S + S/L | 17 |
| LONG | 12 |
| N = 53 | |
|---|---|
| Maternal Age (years) | 34.8 ± 6.4 |
| Age of Child (years) | 6.8 ± 1.8 |
| Male gender (%) | 89.8% |
| Maternal 5-HTTLPR genotype: | |
| S/S | 12 (22.6%) |
| S/L | 22 (41.5%) |
| L/L | 19 (35.8%) |
| Number of Reported Stressors: | |
| Low (0–2) | 29 |
| High (3+) | 24 |
| Maternal autoantibody | 11 (21%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, A.N.; Ferguson, B.J.; Hawkins, E.; Coman, A.; Schauer, J.; Ramirez-Celis, A.; Hecht, P.M.; Bruce, D.; Tilley, M.; Talebizadeh, Z.; et al. The Relationship between Maternal Antibodies to Fetal Brain and Prenatal Stress Exposure in Autism Spectrum Disorder. Metabolites 2023, 13, 663. https://doi.org/10.3390/metabo13050663
Costa AN, Ferguson BJ, Hawkins E, Coman A, Schauer J, Ramirez-Celis A, Hecht PM, Bruce D, Tilley M, Talebizadeh Z, et al. The Relationship between Maternal Antibodies to Fetal Brain and Prenatal Stress Exposure in Autism Spectrum Disorder. Metabolites. 2023; 13(5):663. https://doi.org/10.3390/metabo13050663
Chicago/Turabian StyleCosta, Amy N., Bradley J. Ferguson, Emily Hawkins, Adriana Coman, Joseph Schauer, Alex Ramirez-Celis, Patrick M. Hecht, Danielle Bruce, Michael Tilley, Zohreh Talebizadeh, and et al. 2023. "The Relationship between Maternal Antibodies to Fetal Brain and Prenatal Stress Exposure in Autism Spectrum Disorder" Metabolites 13, no. 5: 663. https://doi.org/10.3390/metabo13050663
APA StyleCosta, A. N., Ferguson, B. J., Hawkins, E., Coman, A., Schauer, J., Ramirez-Celis, A., Hecht, P. M., Bruce, D., Tilley, M., Talebizadeh, Z., Van de Water, J., & Beversdorf, D. Q. (2023). The Relationship between Maternal Antibodies to Fetal Brain and Prenatal Stress Exposure in Autism Spectrum Disorder. Metabolites, 13(5), 663. https://doi.org/10.3390/metabo13050663