
The Interplay between Gut Microbiota and Oral Medications and Its Impact on Advancing Precision Medicine



Abstract
:1. Introduction
2. Susceptibility of Oral Medications to Microbial Metabolism
3. Microbial Drug Metabolism for Different Oral Medications Classes
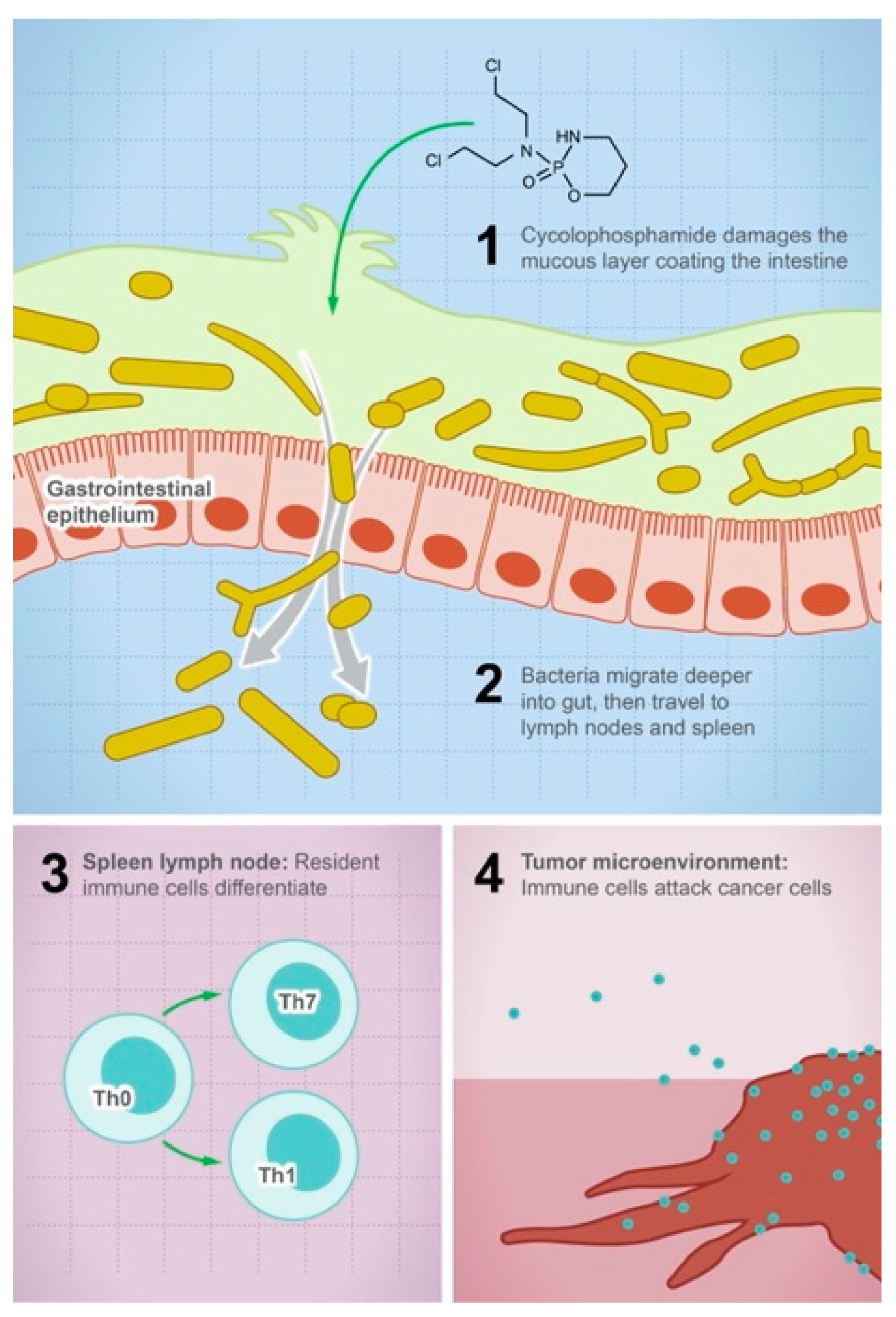
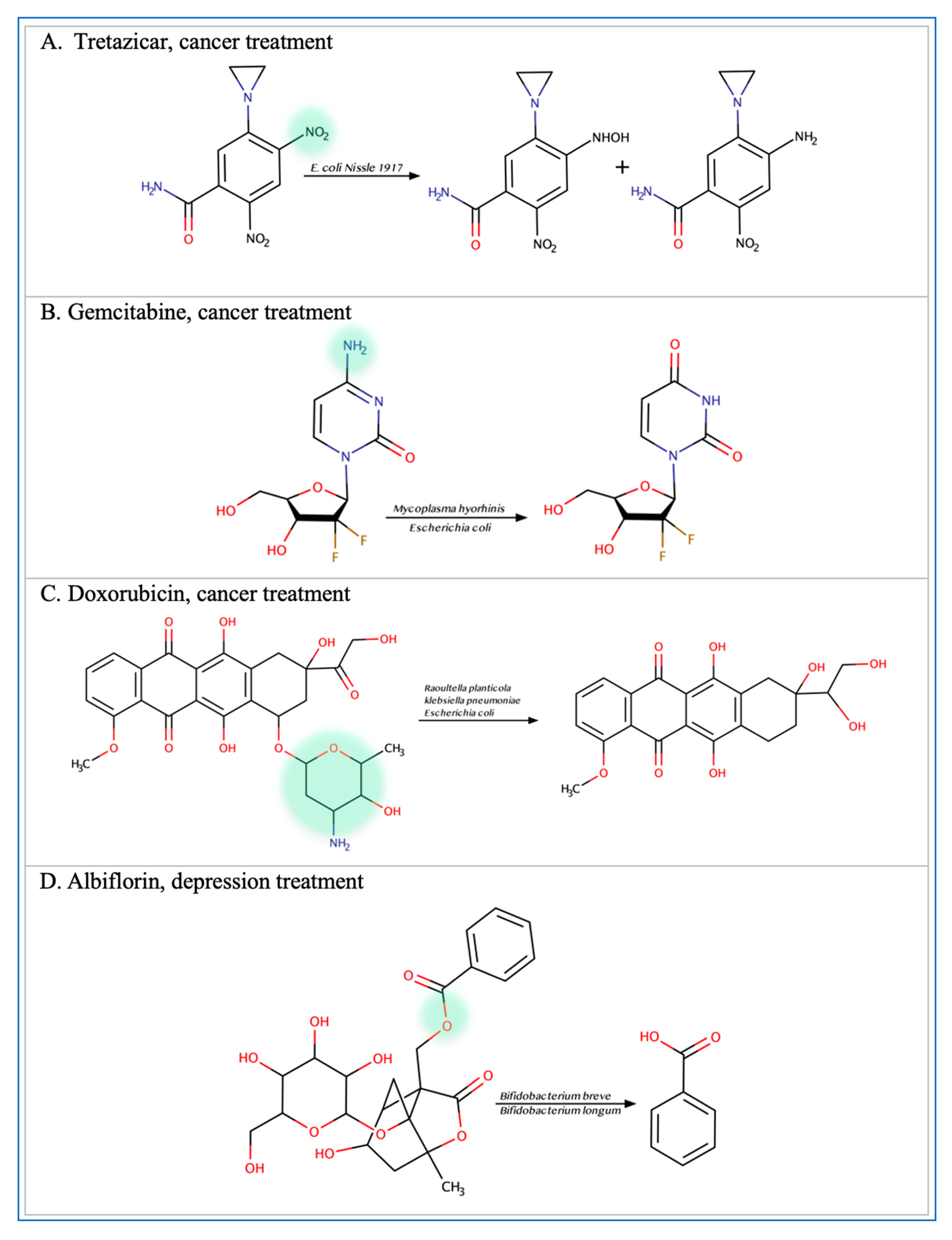
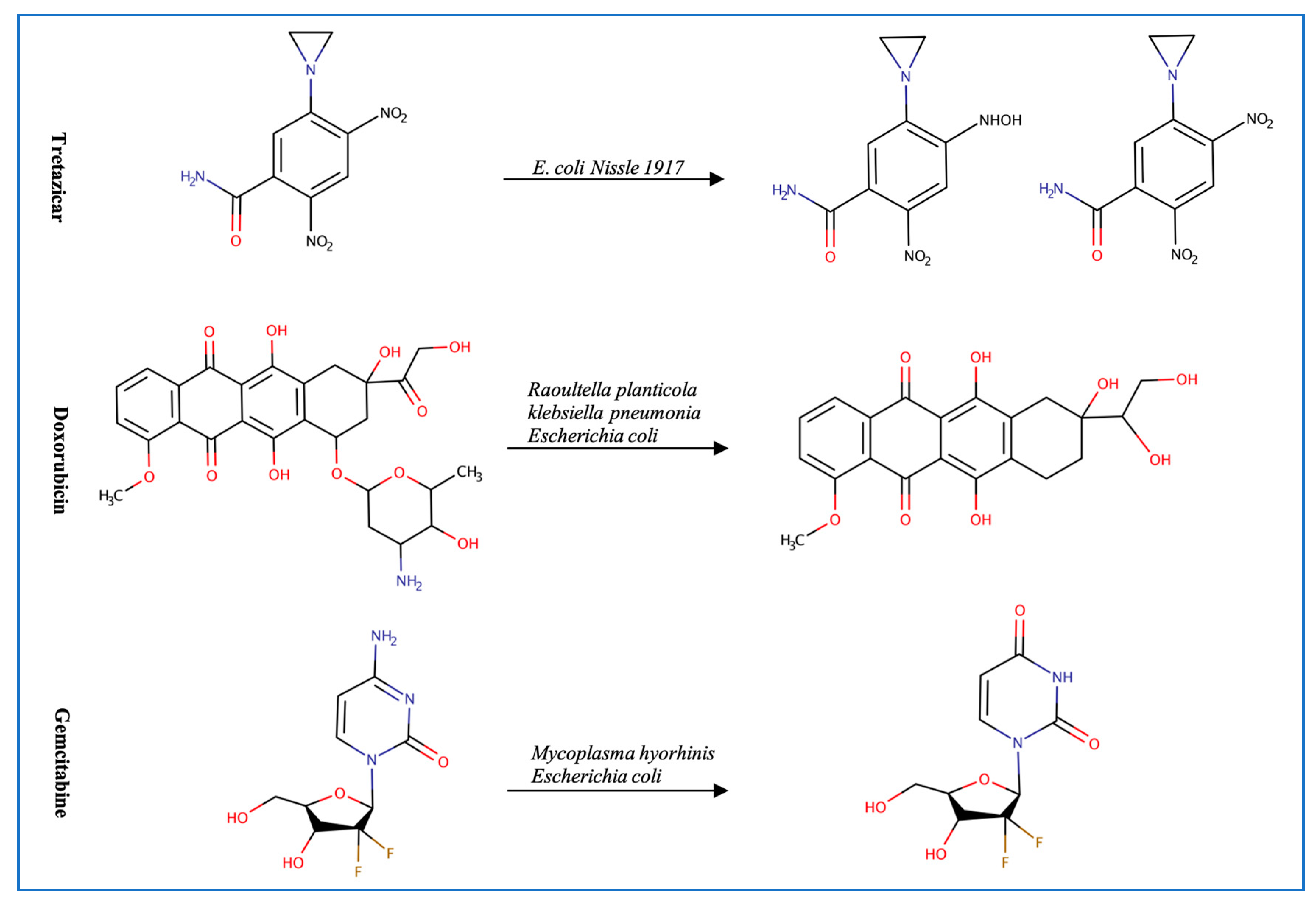
3.1. Anticancer Drugs
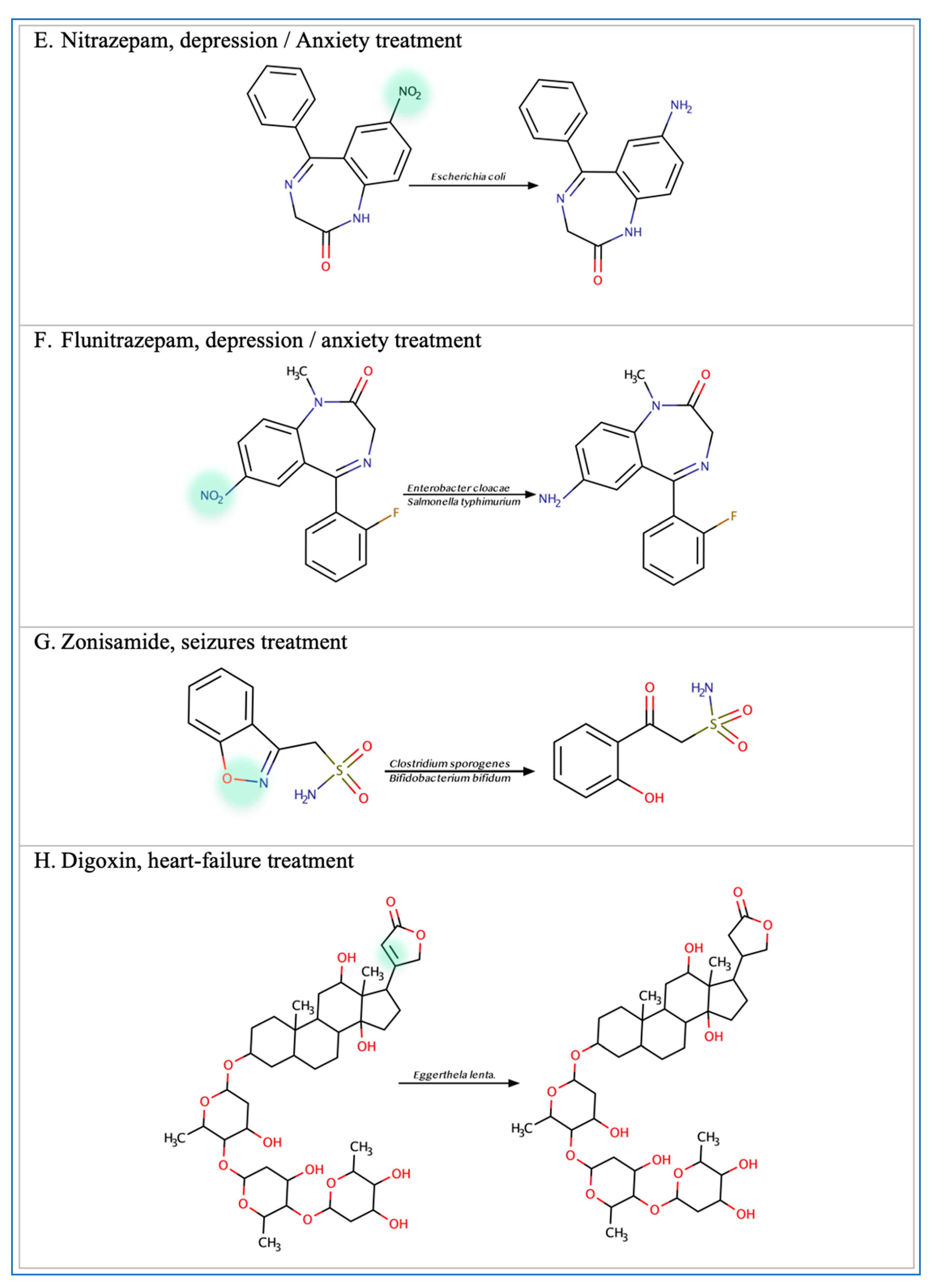
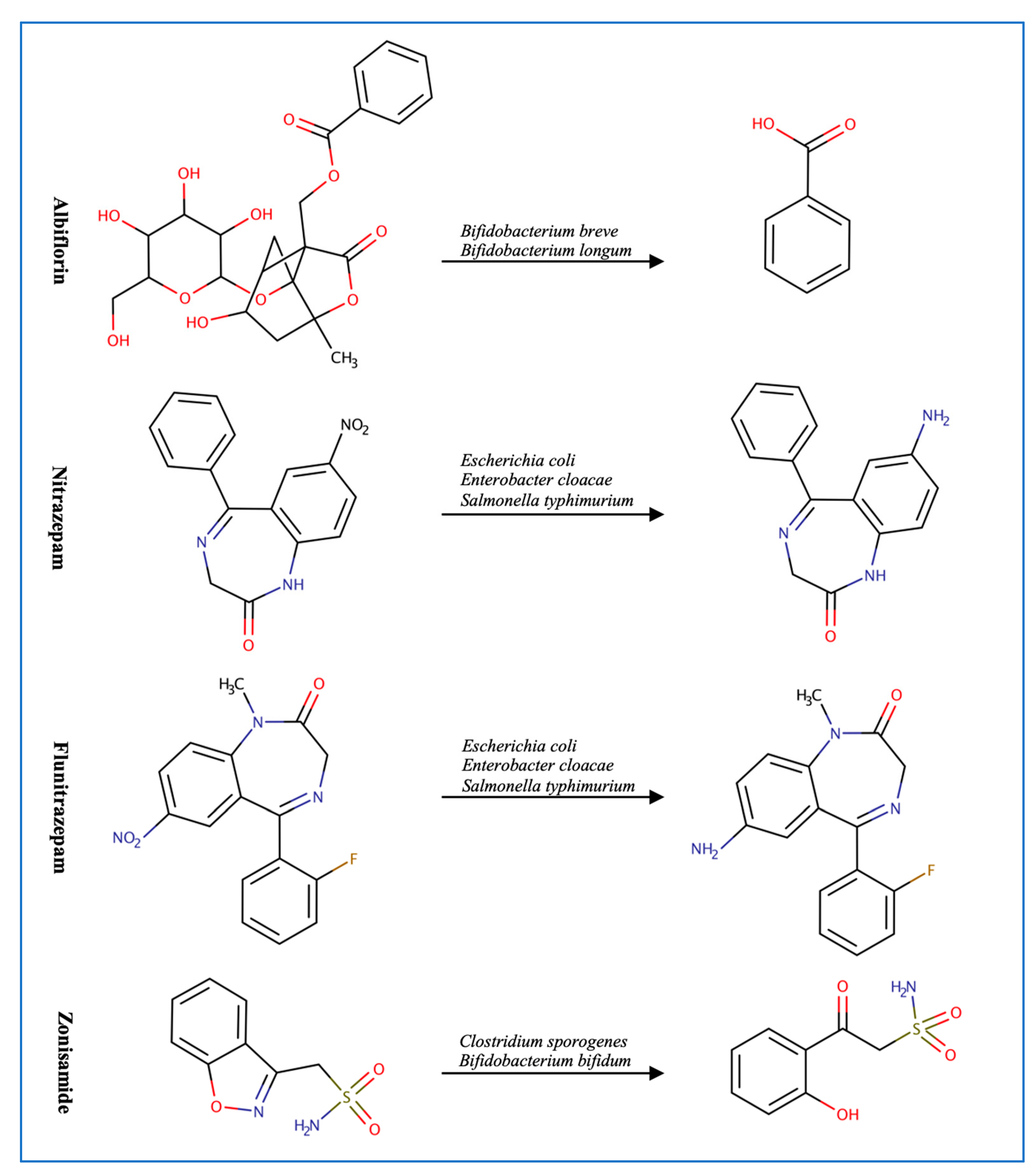
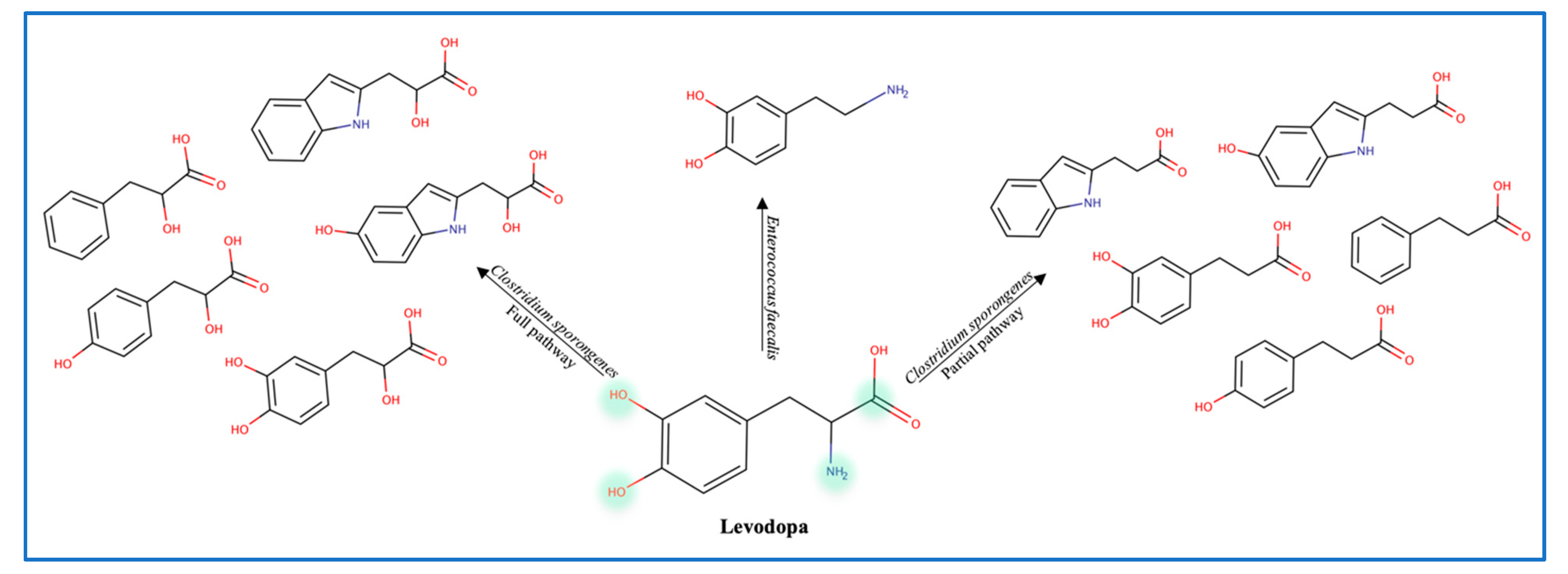
3.2. Central Nervous System Drugs
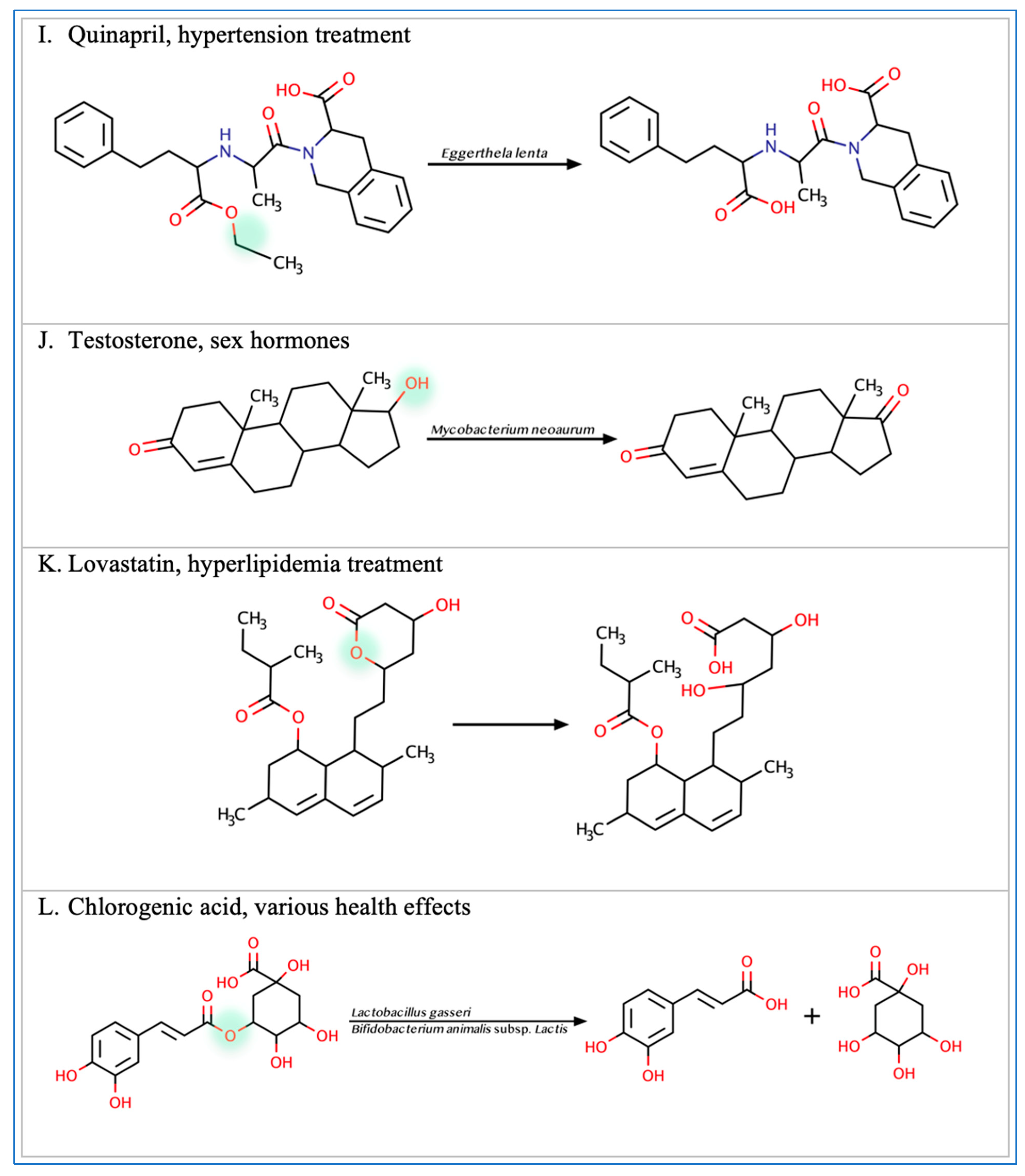
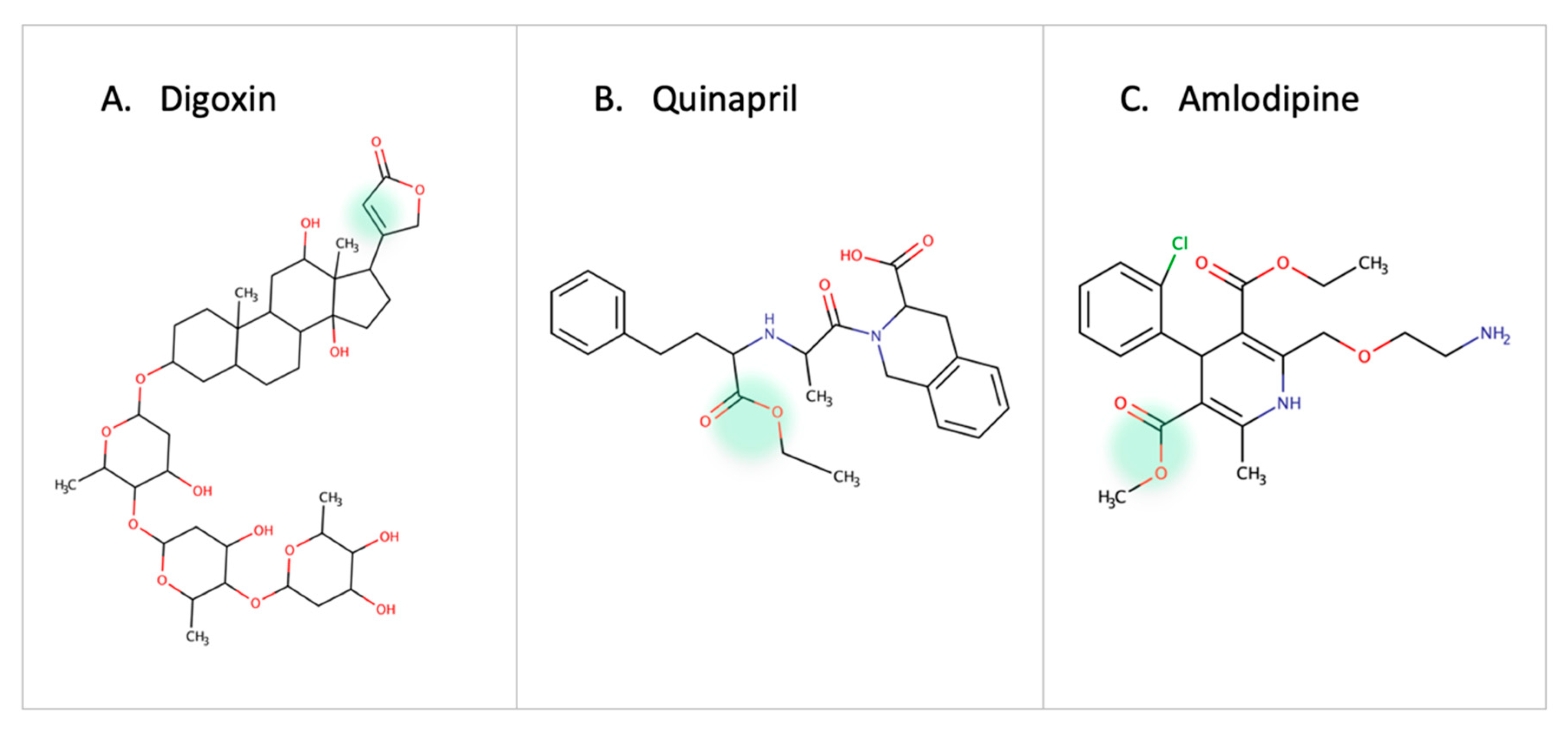
3.3. Cardiovascular Drugs
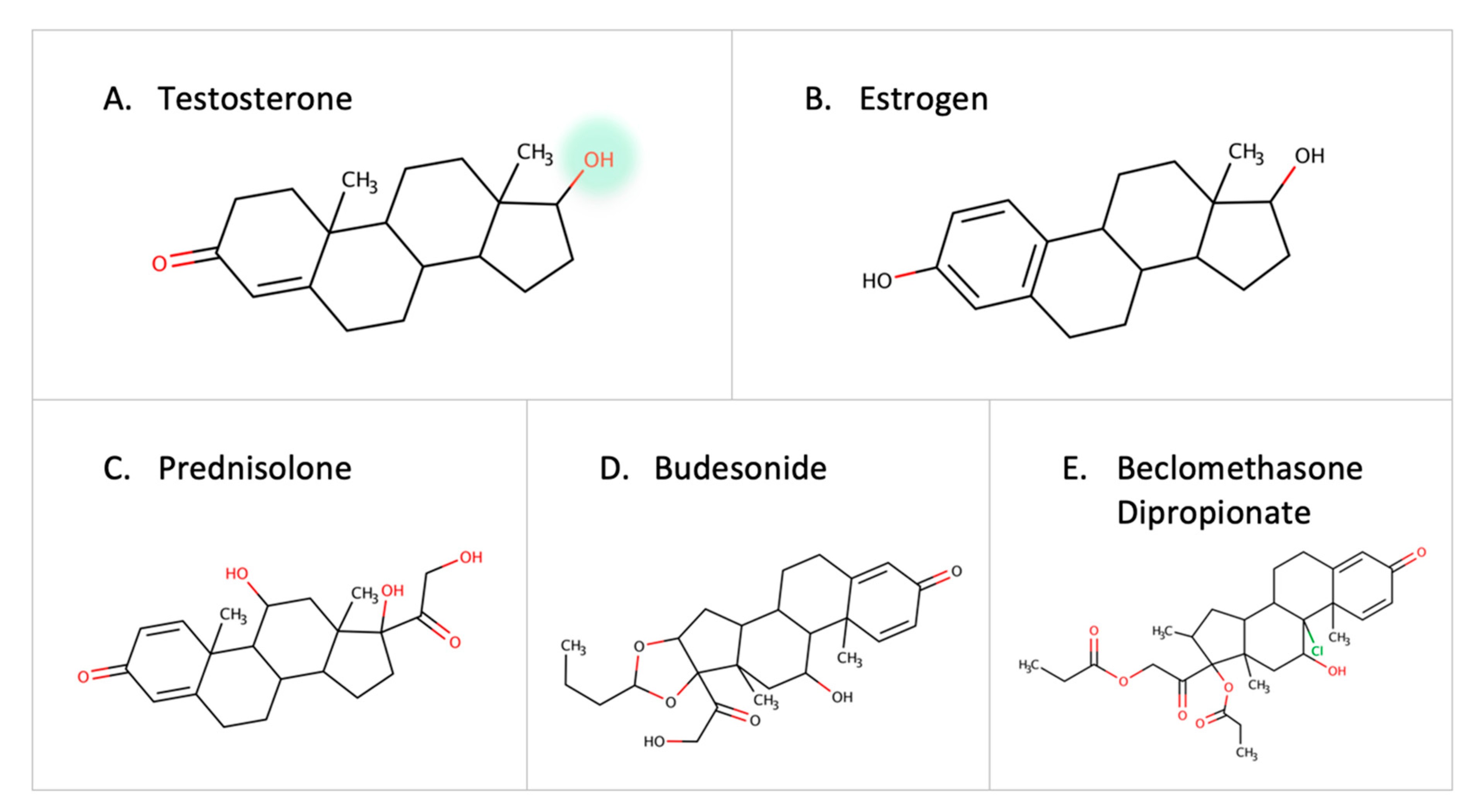
3.4. Steroids and Corticosteroids
3.5. Miscellaneous Xenobiotics and Natural Substances
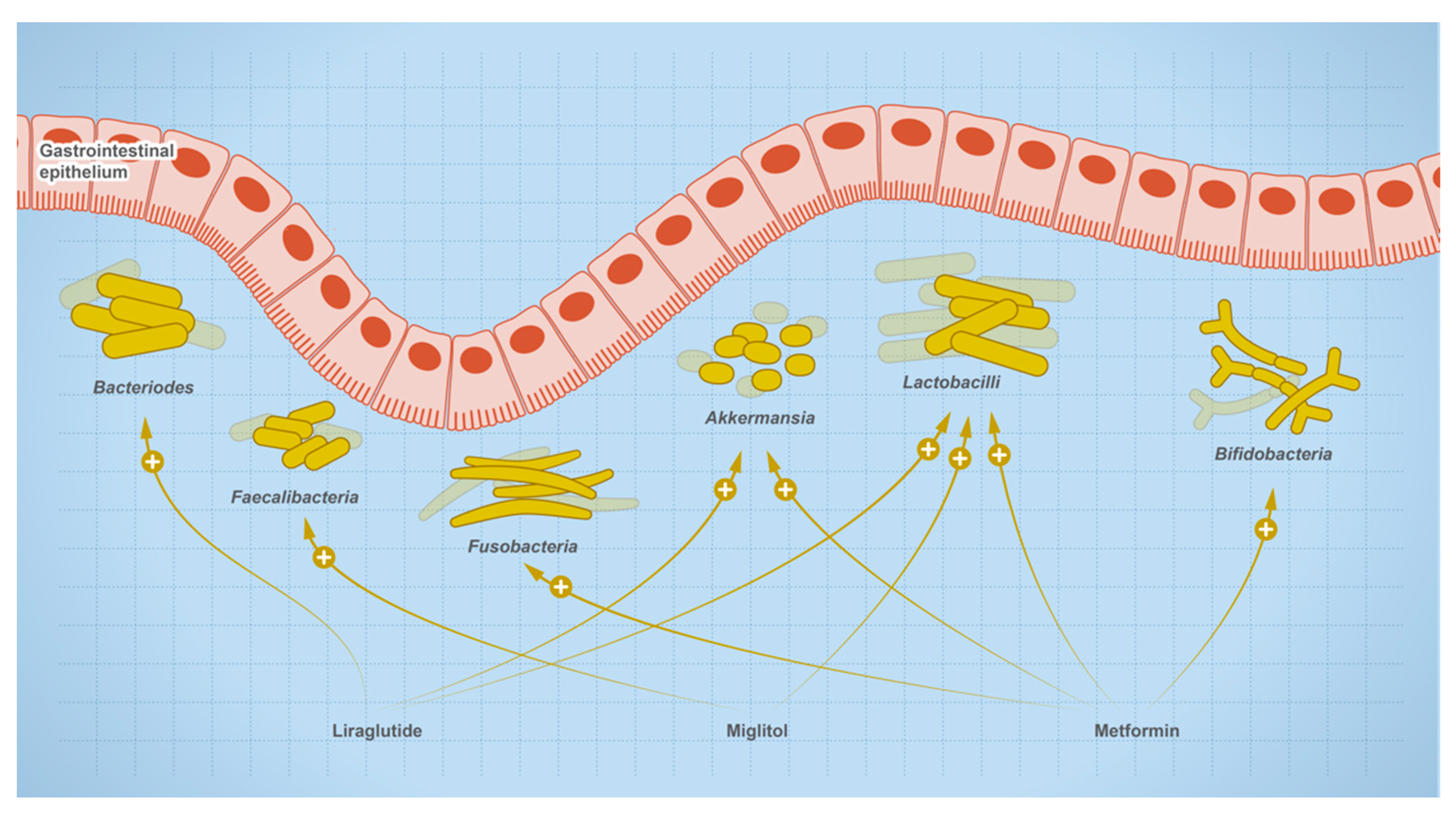
4. Effect of Oral Medications on Gut Microbes
5. Computational Prediction of Microbial Drug Metabolism
6. Discussion
6.1. The Implementation of Pharmacomicrobiomic in Therapeutic Regimes
6.1.1. Challenges of Microbiome-Based Personalized Medicine
6.1.2. Promises of Considering the Microbiome Signature in Prescriptions
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Jia, H.; Cai, X.; Zhong, H.; Feng, Q.; Sunagawa, S.; Arumugam, M.; Kultima, J.R.; Prifti, E.; Nielsen, T.; et al. An integrated catalog of reference genes in the human gut microbiome. Nat. Biotechnol. 2014, 32, 834–841. [Google Scholar] [CrossRef]
- Koppel, N.; Rekdal, V.M.; Balskus, E.P. Chemical transformation of xenobiotics by the human gut microbiota. Science 2017, 356, 1246–1257. [Google Scholar] [CrossRef] [PubMed]
- Garajová, I.; Balsano, R.; Wang, H.; Leonardi, F.; Giovannetti, E.; Deng, D.; Peters, G.J. The role of the microbiome in drug resistance in gastrointestinal cancers. Expert Rev. Anticancer. Ther. 2020, 21, 165–176. [Google Scholar] [CrossRef]
- Lehouritis, P.; Cummins, J.; Stanton, M.; Murphy, C.T.; McCarthy, F.O.; Reid, G.; Tangney, M. Local bacteria affect the efficacy of chemo-therapeutic drugs. Sci. Rep. 2015, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.; Zimmermann-Kogadeeva, M.; Wegmann, R.; Goodman, A.L. Separating host and microbiome contributions to drug pharmacokinetics and toxicity. Science 2019, 363, eaat9931. [Google Scholar] [CrossRef]
- Zimmermann, M.; Zimmermann-Kogadeeva, M.; Wegmann, R.; Goodman, A.L. Mapping human microbiome drug metabolism by gut bacteria and their genes. Nature 2019, 570, 462–467. [Google Scholar] [CrossRef]
- Pant, A.; Maiti, T.K.; Mahajan, D.; Das, B. Human Gut Microbiota and Drug Metabolism. Microb. Ecol. 2022, 1–15. [Google Scholar] [CrossRef]
- Collins, S.L.; Patterson, A.D. The gut microbiome: An orchestrator of xenobiotic metabolism. Acta Pharm. Sin. B 2020, 10, 19–32. [Google Scholar] [CrossRef]
- Yang, T.; Mei, X.; Tackie-Yarboi, E.; Akere, M.T.; Kyoung, J.; Mell, B.; Yeo, J.-Y.; Cheng, X.; Zubcevic, J.; Richards, E.M.; et al. Identification of a Gut Commensal That Compromises the Blood Pressure-Lowering Effect of Ester Angiotensin-Converting Enzyme Inhibitors. Hypertension 2022, 79, 1591–1601. [Google Scholar] [CrossRef]
- Yoo, D.H.; Kim, I.S.; Van Le, T.K.; Jung, I.H.; Yoo, H.H.; Kim, D.H. Gut Microbiota-Mediated Drug. Interactions between Lovastatin and Antibiotics. Drug Metab. Dis. 2016, 42, 1508–1513. [Google Scholar]
- Rekdal, V.M.; Bess, E.N.; Bisanz, J.E.; Turnbaugh, P.J.; Balskus, E.P. Discovery and inhibition of an interspecies gut bacterial pathway for Levodopa metabolism. Science 2019, 364, eaau6323. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.; Han, P.; Fu, J.; Zhang, Z.-W.; Ma, S.-R.; Pan, L.-B.; Xia, Y.-Y.; Yu, H.; Xu, H.; Liu, C.-X.; et al. Esterases from Bifidobacteria Exhibit the Conversion of Albiflorin in Gut Microbiota. Front. Microbiol. 2022, 13, 1174. [Google Scholar] [CrossRef] [PubMed]
- Hashim, H.; Azmin, S.; Razlan, H.; Yahya, N.W.; Tan, H.J.; Manaf, M.R.A.; Ibrahim, N.M. Eradication of Helicobacter pylori infection improves levodopa action, clinical symptoms and quality of life in patients with parkinson’s disease. PLoS ONE 2014, 9, e112330. [Google Scholar] [CrossRef]
- Voorde, J.V.; Sabuncuoğlu, S.; Noppen, S.; Hofer, A.; Ranjbarian, F.; Fieuws, S.; Balzarini, J.; Liekens, S. Nucleoside-catabolizing Enzymes in Mycoplasma-infected Tumor Cell Cultures Compromise the Cytostatic Activity of the Anticancer Drug Gemcitabine. J. Biol. Chem. 2014, 289, 13054–13065. [Google Scholar] [CrossRef]
- Wurdeman, S.R.; Stevens, P.M.; Campbell, J.H.; Davie-Smith, F.; Coulter, E.; Kennon, B.; Geller, L.T.; Barzily-Rokni, M.; Danino, T.; Jonas, O.H.; et al. Potential role of intratumor bacteria in mediating tumor resistance to the chemotherapeutic drug gemcitabine. Physiol. Behav. 2017, 176, 498–503. [Google Scholar]
- Yan, A.; Culp, E.; Perry, J.; Lau, J.T.; MacNeil, L.T.; Surette, M.G.; Wright, G.D. Transformation of the Anticancer Drug Doxorubicin in the Human Gut Microbiome. ACS Infect. Dis. 2018, 4, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Daillère, R.; Hannani, D.; Enot, D.P.; Pfirschke, C.; Engblom, C.; Pittet, M.J.; et al. The Intestinal Microbiota Modulates the Anticancer Immune Effects of Cyclophosphamide. Science 2013, 342, 971–976. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, X. Effects of cyclophosphamide on immune system and gut microbiota in mice. Microbiol. Res. 2015, 171, 97–106. [Google Scholar] [CrossRef]
- Khan, T.J.; Ahmed, Y.M.; Zamzami, M.A.; Siddiqui, A.M.; Khan, I.; Baothman, O.A.S.; Mehanna, M.G.; Kuerban, A.; Kaleemuddin, M.; Yasir, M. Atorvastatin Treatment Modulates the Gut Microbiota of the Hypercholesterolemic Patients. OMICS A J. Integr. Biol. 2018, 22, 154–163. [Google Scholar] [CrossRef]
- Maier, L.; Pruteanu, M.; Kuhn, M.; Zeller, G.; Telzerow, A.; Anderson, E.E.; Brochado, A.R.; Fernandez, K.C.; Dose, H.; Mori, H.; et al. Extensive impact of non-antibiotic drugs on human gut bacteria. Nature 2018, 555, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Xie, C.; Wang, G.; Wu, Y.; Wu, Q.; Wang, X.; Jiang, C. Gut microbiota and intestinal FXR mediate the clinical benefits of met-formin. Nat. Med. 2018, 24, 1919–1929. [Google Scholar] [CrossRef] [PubMed]
- Charoenngam, N.; Shirvani, A.; Kalajian, T.A.; Song, A.; Holick, M.F. The effect of various doses of oral Vitamin D3 supplementation on gut microbiota in healthy adults: A Randomized, Double-blinded, Dose-response Study. Anticancer. Res. 2020, 40, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Dikeocha, I.J.; Al-Kabsi, A.M.; Miftahussurur, M.; Alshawsh, M.A. Pharmacomicrobiomics: Influence of gut microbiota on drug and xenobiotic metabolism. FASEB J. 2022, 36, e22350. [Google Scholar] [CrossRef]
- Chen, H.-Q.; Gong, J.-Y.; Xing, K.; Liu, M.-Z.; Ren, H.; Luo, J.-Q. Pharmacomicrobiomics: Exploiting the Drug-Microbiota Interactions in Antihypertensive Treatment. Front. Med. 2022, 8, 3110. [Google Scholar] [CrossRef]
- Nkera-Gutabara, C.K.; Kerr, R.; Scholefield, J.; Hazelhurst, S.; Naidoo, J. Microbiomics: The Next Pillar of Precision Medicine and Its Role in African Healthcare. Front. Genet. 2022, 13, 616. [Google Scholar] [CrossRef]
- Petrosino, J.F. The microbiome in precision medicine: The way forward. Genome Med. 2018, 10, 12. [Google Scholar] [CrossRef]
- Takeno, S.; Sakai, T. Involvement of the intestinal microflora in nitrazepam-induced teratogenicity in rats and its relationship to nitroreduction. Teratology 1991, 44, 209–214. [Google Scholar] [CrossRef]
- LinWu, S.-W.; Syu, C.-J.; Chen, Y.-L.; Wang, A.H.-J.; Peng, F.-C. Characterization of Escherichia coli nitroreductase NfsB in the metabolism of nitrobenzodiazepines. Biochem. Pharmacol. 2009, 78, 96–103. [Google Scholar] [CrossRef]
- Xie, Y.; Hu, F.; Xiang, D.; Lu, H.; Li, W.; Zhao, A.; Huang, L.; Wang, R. The metabolic effect of gut microbiota on drugs. Drug. Metab. Rev. 2020, 52, 139–156. [Google Scholar] [CrossRef]
- Gingell, R.; Bridges, J.W.; Williams, R.T. The Role of the Gut Flora in the Metabolism of Prontosil and Neoprontosil in the Rat. Xenobiotica 1971, 129, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Hayllar, J.; Bjarnason, I. Sulphasalazine in ulcerative colitis: In memoriam? Gut 1991, 32, 462–463. [Google Scholar] [CrossRef] [PubMed]
- Chan, R.P.; Pope, D.J.; Gilbert, A.P.; Sacra, P.J.; Baron, J.H.; Lennard-Jones, J.E. Studies of two novel sulfasalazine analogs, ipsalazide and balsalazide. Dig. Dis. Sci. 1983, 28, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Wadword, A.N.; Fitton, A. Olsalazine: A review of its pharmacodynamic and pharmacokinetic properties, and therapeutic potential in inflammatory bowel disease. Drugs 1991, 41, 647–664. [Google Scholar] [CrossRef]
- Basit, A.W.; Newton, J.; Lacey, L.F. Susceptibility of the H2-receptor antagonists cimetidine, famotidine and nizatidine, to metabolism by the gastrointestinal microflora. Int. J. Pharm. 2002, 237, 23–33. [Google Scholar] [CrossRef]
- Kim, D.-H.; Hyun, S.-H.; Shim, S.-B.; Kobashi, K. The Role of Intestinal Bacteria in the Transformation of Sodium Picosulfate. Jpn. J. Pharmacol. 1992, 59, 1–5. [Google Scholar] [CrossRef]
- Kodawara, T.; Higashi, T.; Negoro, Y.; Kamitani, Y.; Igarashi, T.; Watanabe, K.; Tsukamoto, H.; Yano, R.; Masada, M.; Iwasaki, H.; et al. The Inhibitory Effect of Ciprofloxacin on the β-Glucuronidase-mediated Deconjugation of the Irinotecan Metabolite SN-38-G. Basic Clin. Pharmacol. Toxicol. 2016, 118, 333–337. [Google Scholar]
- Shu, Y.Z.; Kingston, D.G.I.; Van Tassell, R.L.; Wilkins, T.D. Metabolism of levamisole, an anti-colon cancer drug, by human intestinal bacteria. Xenobiotica 1991, 21, 737–750. [Google Scholar] [CrossRef]
- Rafii, F.; Sutherland, J.B.; Hansen, J.E.B.; Cerniglia, C.E.; Hansen, E.B. Reduction of nitrazepam by Clostridium leptum, a nitroreductase-producing bacterium isolated from the human intestinal tract. Clin. Infect. Dis. 1997, 25, S121–S122. [Google Scholar] [CrossRef]
- Magnusson, J.; Bergdahl, B.; Bogentoft, C.; Jonsson, U. Metabolism of digoxin and absorption site. Br. J. Clin. Pharmacol. 1982, 14, 284–285. [Google Scholar] [CrossRef]
- Tozaki, H.; Emi, Y.; Horisaka, E.; Fujita, T.; Yamamoto, A.; Muranishi, S. Degradation of Insulin and Calcitonin and Their Protection by Various Protease Inhibitors in Rat Caecal Contents: Implications in Peptide Delivery to the Colon. J. Pharm. Pharmacol. 1997, 49, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Sousa, T.; Paterson, R.; Moore, V.; Carlsson, A.; Abrahamsson, B.; Basit, A.W. The gastrointestinal microbiota as a site for the bio-transformation of drugs. Int. J. Pharm. 2008, 363, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Noh, K.; Kang, Y.R.; Nepal, M.R.; Shakya, R.; Kang, M.J.; Kang, W.; Lee, S.; Jeong, H.G.; Jeong, T.C. Impact of gut microbiota on drug metabolism: An update for safe and effective use of drugs. Arch. Pharmacal Res. 2017, 40, 1345–1355. [Google Scholar] [CrossRef] [PubMed]
- Trinh, H.-T.; Joh, E.-H.; Kwak, H.-Y.; Baek, N.-I.; Kim, D.-H. Anti-pruritic effect of baicalin and its metabolites, baicalein and oroxylin A, in mice. Acta Pharmacol. Sin. 2010, 31, 718–724. [Google Scholar] [CrossRef]
- Zhang, M.; Peng, C.S.; Li, X.B. In vivo and in vitro metabolites from the main diester and monoester diterpenoid alkaloids in a traditional Chinese herb, the aconitum species. Evid. Based Complement. Altern. Med. 2015, 252434. [Google Scholar]
- Roowi, S.; Mullen, W.; Edwards, C.A.; Crozier, A. Yoghurt impacts on the excretion of phenolic acids derived from colonic breakdown of orange juice flavanones in humans. Mol. Nutr. Food Res. 2009, 53, S68–S75. [Google Scholar] [CrossRef]
- Liu, X.; Suzuki, N.; Laxmi, Y.S.; Okamoto, Y.; Shibutani, S. Anti-breast cancer potential of daidzein in rodents. Life Sci. 2012, 91, 415–419. [Google Scholar] [CrossRef]
- Park, E.-K.; Shin, J.; Bae, E.-A.; Lee, Y.-C.; Kim, D.-H. Intestinal Bacteria Activate Estrogenic Effect of Main Constituents Puerarin and Daidzin of Pueraria thunbergiana. Biol. Pharm. Bull. 2006, 29, 2432–2435. [Google Scholar] [CrossRef]
- Derosa, L.; Hellmann, M.D.; Spaziano, M.; Halpenny, D.; Fidelle, M.; Rizvi, H.; Long, N.; Plodkowski, A.J.; Arbour, K.C.; Chaft, J.E.; et al. Negative association of antibiotics on clinical activity of immune checkpoint inhibitors in patients with advanced renal cell and non-small-cell lung cancer. Ann. Oncol. 2018, 29, 1437–1444. [Google Scholar] [CrossRef]
- Routy, B.; le Chatelier, E.; DeRosa, L.; Duong, C.P.M.; Alou, M.T.; Daillère, R.; Fluckiger, A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut microbiome influences efficacy of PD-1–based immunotherapy against epithelial tumors. Science 2018, 97, 91–97. [Google Scholar] [CrossRef]
- Huang, J.; Liu, W.; Kang, W.; He, Y.; Yang, R.; Mou, X.; Zhao, W. Effects of microbiota on anticancer drugs: Current knowledge and potential applications. eBioMedicine 2022, 83, 104197. [Google Scholar] [CrossRef]
- Matson, V.; Fessler, J.; Bao, R.; Chongsuwat, T.; Zha, Y.; Alegre, M.-L.; Luke, J.J.; Gajewski, T.F. The commensal microbiome is associated with anti–PD-1 efficacy in metastatic melanoma patients. Science 2018, 359, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnan, V.; Spencer, C.N.; Nezi, L.; Reuben, A.; Andrews, M.C.; Karpinets, T.V.; Prieto, P.A.; Vicente, D.; Hoffman, K.; Wei, S.C.; et al. Gut microbiome modulates response to anti–PD-1 immunotherapy in melanoma patients. Science 2018, 359, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Chaput, N.; Lepage, P.; Coutzac, C.; Soularue, E.; Le Roux, K.; Monot, C.; Boselli, L.; Routier, E.; Cassard, L.; Collins, M.; et al. Baseline gut microbiota predicts clinical response and colitis in metastatic melanoma patients treated with ipilimumab. Ann. Oncol. 2017, 28, 1368–1379, Erratum in Ann. Oncol. 2019, 30, 2012. [Google Scholar] [CrossRef] [PubMed]
- Capuco, A.; Urits, I.; Hasoon, J.; Chun, R.; Gerald, B.; Wang, J.K.; Kassem, H.; Ngo, A.L.; Abd-Elsayed, A.; Simopoulos, T.; et al. Current Perspectives on Gut Microbiome Dysbiosis and Depression. Adv. Ther. 2020, 37, 1328–1346. [Google Scholar] [CrossRef]
- Wingfield, B.; Lapsley, C.; McDowell, A.; Miliotis, G.; McLafferty, M.; O’neill, S.M.; Coleman, S.; McGinnity, T.M.; Bjourson, A.J.; Murray, E.K. Variations in the oral microbiome are associated with depression in young adults. Sci. Rep. 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Lach, G.; Schellekens, H.; Dinan, T.G.; Cryan, J.F. Anxiety, Depression, and the Microbiome: A Role for Gut Peptides. Neurotherapeutics 2018, 15, 36–59. [Google Scholar] [CrossRef]
- Elmer, G.W.; Remmel, R.P. Role of the intestinal microflora in clonazepam metabolism in the rat. Xenobiotica 1984, 14, 829–840. [Google Scholar] [CrossRef]
- Kitamura, S.; Sugihara, K.; Kuwasako, M.; Tatsumi, K. The Role of Mammalian Intestinal Bacteria in the Reductive Metabolism of Zonisamide. J. Pharm. Pharmacol. 1997, 49, 253–256. [Google Scholar] [CrossRef]
- Connil, N.; Le Breton, Y.; Dousset, X.; Auffray, Y.; Rincé, A.; Prévost, H. Identification of the Enterococcus faecalis tyrosine decarboxylase operon involved in tyramine production. Appl. Environ. Microbiol. 2002, 68, 3537–3544. [Google Scholar] [CrossRef]
- Fasano, A.; Bove, F.; Gabrielli, M.; Petracca, M.; Zocco, M.A.; Ragazzoni, E.; Barbaro, F.; Piano, C.; Fortuna, S.; Tortora, A.; et al. The role of small intestinal bacterial overgrowth in Parkinson’s disease. Mov. Disord. 2013, 28, 1241–1249. [Google Scholar] [CrossRef]
- Van Kessel, S.P.; de Jong, H.R.; Winkel, S.L.; van Leeuwen, S.S.; Nelemans, S.A.; Permentier, H.; Keshavarzian, A.; El Aidy, S. Gut bacterial deamination of residual levodopa medication for Parkinson’s disease. BMC Biol. 2020, 18, 137. [Google Scholar] [CrossRef]
- Haiser, H.J.; Seim, K.L.; Balskus, E.P.; Turnbaugh, P.J. Mechanistic insight into digoxin inactivation by Eggerthella lenta augments our understanding of its pharmacokinetics. Gut Microbes. 2014, 5, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Jaiswal, S.K.; Dhoke, G.V.; Srivastava, G.N.; Sharma, A.K.; Sharma, V.K. Mechanistic and structural insight into promis-cuity based metabolism of cardiac drug digoxin by gut microbial enzyme. J. Cell. Biochem. 2018, 119, 5287–5296. [Google Scholar] [CrossRef]
- Dobkin, J.F.; Saha, J.R.; Butler, V.P., Jr.; Neu, H.C.; Lindenbaum, J. Digoxin-Inactivating Bacteria: Identification in Human Gut Flora. Science 1982, 5, 5–7. [Google Scholar] [CrossRef]
- Yoo, H.H.; Kim, I.S.; Yoo, D.H.; Kim, D.H. Effects of orally administered antibiotics on the bioavailability of amlodipine: Gut microbio-ta-mediated drug interaction. J. Hypertens. 2016, 34, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Holert, J.; Cardenas, E.; Bergstrand, L.H.; Zaikova, E.; Hahn, A.S.; Hallam, S.J.; Mohn, W.W. Metagenomes Reveal Global Distribution of Bacterial Steroid Catabolism in Natural, Engineered, and Host Environments. mBio 2018, 9, e02345-17. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, R.; Wang, M.; Peng, R.; Fu, S.; Fu, A.; Liu, T. 3β-Hydroxysteroid dehydrogenase expressed by gut microbes degrades tes-tosterone and is linked to depression in males. Cell Host Microbe 2022, 30, 329–339.e5. [Google Scholar] [CrossRef]
- Bhasin, S.; Seidman, S. Testosterone Treatment of Depressive Disorders in Men: Too Much Smoke, Not Enough High-Quality Evidence. JAMA Psychiatry 2019, 76, 9–10. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.C.; Tajar, A.; Beynon, J.M.; Pye, S.R.; Silman, A.J.; Finn, J.D.; O’Neill, T.W.; Bartfai, G.; Casanueva, F.F.; Forti, G.; et al. Identification of Late-Onset Hypogonadism in Middle-Aged and Elderly Men. N. Engl. J. Med. 2010, 363, 123–135. [Google Scholar] [CrossRef]
- García-Gómez, E.; González-Pedrajo, B.; Camacho-Arroyo, I. Role of Sex Steroid Hormones in Bacterial-Host Interactions. BioMed Res. Int. 2013, 2013, 928290. [Google Scholar] [CrossRef]
- Morris, G.N.; Winter, J.; Cato, E.P. Clostridium scindens sp. nov., a human intestinal bacterium with desmolytic activity on cor-ticoids. Int. J. Syst. Bacteriol. 1985, 35, 478–481. [Google Scholar] [CrossRef]
- Flores, R.; Shi, J.; Fuhrman, B.; Xu, X.; Veenstra, T.D.; Gail, M.H.; Gajer, P.; Ravel, J.; Goedert, J.J. Fecal microbial determinants of fecal and systemic estrogens and estrogen metabolites: A cross-sectional study. J. Transl. Med. 2012, 10, 253. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Wu, J.; Chen, J. The Role of Gut Microbial β-Glucuronidase in Estrogen Reactivation and Breast Cancer. Front. Cell Dev. Biol. 2021, 9, 631552. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.; Gaisford, S.; Merchant, H.A.; Basit, A.W. Colonic bacterial metabolism of corticosteroids. Int. J. Pharm. 2013, 457, 268–274. [Google Scholar] [CrossRef]
- Needham, B.D.; Kaddurah-Daouk, R.; Mazmanian, S.K. Gut microbial molecules in behavioural and neurodegenerative condi-tions. Nat. Rev. Neurosci. 2020, 21, 717–731. [Google Scholar] [CrossRef]
- Bora, S.A.; Kennett, M.J.; Smith, P.B.; Patterson, A.D.; Cantorna, M.T. The Gut Microbiota Regulates Endocrine Vitamin D Metabolism through Fibroblast Growth Factor 23. Front. Immunol. 2018, 9, 408. [Google Scholar] [CrossRef]
- Ran, L.; Liu, A.B.; Lee, M.; Xie, P.; Lin, Y.; Yang, C.S. Effects of antibiotics on degradation and bioavailability of different vitamin E forms in mice. Biofactors 2019, 45, 450–462. [Google Scholar] [CrossRef]
- Liang, N.; Kitts, D.D. Role of Chlorogenic Acids in Controlling Oxidative and Inflammatory Stress Conditions. Nutrients 2015, 8, 16. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, J.; Ballevre, O.; Luo, H.; Zhang, W. Antihypertensive effects and mechanisms of chlorogenic acids. Hypertens. Res. 2012, 35, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Cao, J.; Feng, Q.; Peng, J.; Hu, Y. Roles of Chlorogenic Acid on Regulating Glucose and Lipids Metabolism: A Review. Evidence-Based Complement. Altern. Med. 2013, 2013, 801457. [Google Scholar] [CrossRef]
- Couteau, D.; McCartney, A.L.; Gibson, G.R.; Williamson, G. Isolation and characterization of human colonic bacteria able to hy-drolyse chlorogenic acid. J. Appl. Microbiol. 2001, 90, 873–881. [Google Scholar] [CrossRef]
- Curiel, J.A.; Rodríguez, H.; Landete, J.M.; de las Rivas, B.; Muñoz, R. Ability of Lactobacillus brevis strains to degrade food phenolic acids. Food Chem. 2010, 120, 225–229. [Google Scholar] [CrossRef]
- Shin, N.R.; Lee, J.C.; Lee, H.Y.; Kim, M.S.; Whon, T.W.; Lee, M.S.; Bae, J.W. An increase in the Akkermansia spp. population induced by metformin treatment improves glucose homeostasis in diet-induced obese mice. Gut 2014, 63, 727–735. [Google Scholar] [CrossRef]
- Cabreiro, F.; Au, C.; Leung, K.-Y.; Vergara-Irigaray, N.; Cochemé, H.M.; Noori, T.; Weinkove, D.; Schuster, E.; Greene, N.D.; Gems, D. Metformin Retards Aging in C. elegans by Altering Microbial Folate and Methionine Metabolism. Cell 2013, 153, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Mato, J.M.; Martínez-Chantar, M.L.; Lu, S.C. Methionine Metabolism and Liver Disease. Annu. Rev. Nutr. 2008, 28, 273–293. [Google Scholar] [CrossRef] [PubMed]
- Weersma, R.K.; Zhernakova, A.; Fu, J. Interaction between drugs and the gut microbiome. Gut 2020, 69, 1510–1519. [Google Scholar] [CrossRef]
- Vich Vila, A.; Collij, V.; Sanna, S.; Sinha, T.; Imhann, F.; Bourgonje, A.; Mujagic, Z.; Jonkers, D.M.A.E.; Masclee, A.A.M.; Fu, J.; et al. Impact of commonly used drugs on the composition and metabolic function of the gut microbiota. Nat. Commun. 2020, 11, 362. [Google Scholar] [CrossRef]
- Nayak, R.R.; Alexander, M.; Deshpande, I.; Stapleton-Gray, K.; Rimal, B.; Patterson, A.D.; Ubeda, C.; Scher, J.U.; Turnbaugh, P.J. Methotrexate impacts conserved pathways in diverse human gut bacteria leading to decreased host immune activation. Cell Host Microbe 2021, 29, 362–377.e11. [Google Scholar] [CrossRef]
- Yao, Z.; Zhao, M.; Gong, Y.; Chen, W.; Wang, Q.; Fu, Y.; Guo, T.; Zhao, J.; Gao, L.; Bo, T. Relation of Gut Microbes and L-Thyroxine Through Altered Thyroxine Metabolism in Subclinical Hypothyroidism Subjects. Front. Cell. Infect. Microbiol. 2020, 10, 495. [Google Scholar] [CrossRef]
- Jackson, M.A.; Verdi, S.; Maxan, M.-E.; Shin, C.M.; Zierer, J.; Bowyer, R.C.E.; Martin, T.; Williams, F.M.K.; Menni, C.; Bell, J.T.; et al. Gut microbiota associations with common diseases and prescription medications in a population-based cohort. Nat. Commun. 2018, 9, 2655. [Google Scholar] [CrossRef]
- ElRakaiby, M.; Dutilh, B.E.; Rizkallah, M.R.; Boleij, A.; Cole, J.N.; Aziz, R.K. Pharmacomicrobiomics: The Impact of Human Microbiome Variations on Systems Pharmacology and Personalized Therapeutics. OMICS: A J. Integr. Biol. 2014, 18, 402–414. [Google Scholar] [CrossRef]
- Zeng, X.; Yang, X.; Fan, J.; Tan, Y.; Ju, L.; Shen, W.; Wang, Y.; Wang, X.; Chen, W.; Ju, D.; et al. MASI: Microbiota—Active substance interactions database. Nucleic Acids Res. 2021, 49, D776–D782. [Google Scholar] [CrossRef]
- Zhou, J.; Ouyang, J.; Gao, Z.; Qin, H.; Jun, W.; Shi, T. MagMD: Database summarizing the metabolic action of gut microbiota to drugs. Comput. Struct. Biotechnol. J. 2022, 20, 6427–6430. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.K.; Jaiswal, S.K.; Chaudhary, N.; Sharma, V.K. A novel approach for the prediction of species-specific biotransformation of xenobiotic/drug molecules by the human gut microbiota. Sci. Rep. 2017, 7, 9751. [Google Scholar] [CrossRef] [PubMed]
- McCoubrey, L.E.; Thomaidou, S.; Elbadawi, M.; Gaisford, S.; Basit, A.W.; Orlu, M. Machine Learning Predicts Drug Metabolism and Bioaccumulation by Intestinal Microbiota. Pharmaceutics 2021, 13, 2001. [Google Scholar] [CrossRef] [PubMed]
- McCoubrey, L.E.; Elbadawi, M.; Orlu, M.; Gaisford, S.; Basit, A.W. Machine Learning Uncovers Adverse Drug Effects on Intestinal Bacteria. Pharmaceutics 2021, 13, 1026. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, L.; Kelly, L. Bringing microbiome-drug interaction research into the clinic. eBioMedicine 2019, 44, 708–715. [Google Scholar] [CrossRef]
- Sabater, A.; Ciudad, C.J.; Cendros, M.; Dobrokhotov, D.; Sabater-Tobella, J. g-Nomic: A new pharmacogenetics interpretation software. Pharmacogenomics Pers. Med. 2019, 12, 75–85. [Google Scholar] [CrossRef]
- Karp, P.D.; Paley, S.; Krummenacker, M.; Kothari, A.; Wannemuehler, M.J.; Phillips, G.J. Pathway Tools Management of Path-way/Genome Data for Microbial Communities. Front. Bioinform. 2022, 2, 43. [Google Scholar] [CrossRef]
- Kim, J.; Choi, J.-P.; Kim, M.S.; Bhak, J. PharmaKoVariome database for supporting genetic testing. Database 2022, 2022, baac092. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Tran, P.Q.; Breister, A.M.; Liu, Y.; Kieft, K.; Cowley, E.S.; Karaoz, U.; Anantharaman, K. METABOLIC: High-throughput profiling of microbial genomes for functional traits, metabolism, biogeochemistry, and community-scale functional networks. Microbiome 2022, 10, 33. [Google Scholar] [CrossRef]
- Bajinka, O.; Tan, Y.; Abdelhalim, K.A.; Özdemir, G.; Qiu, X. Extrinsic factors influencing gut microbes, the immediate consequences and restoring eubiosis. AMB Express 2020, 10, 130. [Google Scholar] [CrossRef] [PubMed]
- Kurilshikov, A.; Medina-Gomez, C.; Bacigalupe, R.; Radjabzadeh, D.; Wang, J.; Demirkan, A.; Le Roy, C.I.; Garay, J.A.R.; Finnicum, C.T.; Liu, X.; et al. Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat. Genet. 2021, 53, 156–165. [Google Scholar] [CrossRef]
- Johnsen, A.R.; Kroer, N. Effects of stress and other environmental factors on horizontal plasmid transfer assessed by direct quantification of discrete transfer events. FEMS Microbiol. Ecol. 2007, 59, 718–728. [Google Scholar] [CrossRef]
- Heinken, A.; Hertel, J.; Acharya, G.; Ravcheev, D.A.; Nyga, M.; Okpala, O.E.; Hogan, M.; Magnúsdóttir, S.; Martinelli, F.; Nap, B.; et al. Genome-scale metabolic reconstruction of 7,302 human microorganisms for personalized medicine. Nat. Biotechnol. 2023, 1–12. [Google Scholar] [CrossRef]
- Pakti, M.; Saraswat, A.; Bhutkar, S.; Dukhande, V.; Patel, K. Combination of gemcitabine and cytidine deaminase inhibitor: A breakthrough cocktail for the treatment of pancreatic ductal adenocarcinom. Am. Assoc. Cancer Res. 2021, 81, 1059. [Google Scholar]
- Rudzińska, M.; Daglioglu, C.; Savvateeva, L.V.; Kaci, F.N.; Antoine, R.; Zamyatnin, A.A. Status and perspectives of pro-tease inhibitors and their combination with nanosized drug delivery systems for targeted cancer therapy. Drug Des. Devel. Ther. 2021, 15, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Nyholm, D.; Hellström, P.M. Effects of helicobacter pylori on levodopa pharmacokinetics. J. Parkinsons Dis. 2021, 11, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Lolekha, P.; Sriphanom, T.; Vilaichone, R.-K. Helicobacter pylori eradication improves motor fluctuations in advanced Parkinson’s disease patients: A prospective cohort study (HP-PD trial). PLoS ONE 2021, 16, e0251042. [Google Scholar] [CrossRef] [PubMed]
- Purdel, C.; Ungurianu, A.; Adam-Dima, I.; Margină, D. Exploring the potential impact of probiotic use on drug metabolism and efficacy. Biomed. Pharmacother. 2023, 161, 114468. [Google Scholar] [CrossRef]
- Olendzki, B.; Bucci, V.; Cawley, C.; Maserati, R.; McManus, M.; Olednzki, E.; Madziar, C.; Chiang, D.; Ward, D.V.; Pellish, R.; et al. Dietary manipulation of the gut microbiome in inflammatory bowel disease patients: Pilot study. Gut Microbes 2022, 14, 2046244. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal–response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 9, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Falà, A.K.; Álvarez-Ordóñez, A.; Filloux, A.; Gahan, C.G.M.; Cotter, P.D. Quorum sensing in human gut and food microbiomes: Sig-nificance and potential for therapeutic targeting. Front. Microbiol. 2022, 13, 4389. [Google Scholar] [CrossRef]
- Rutherford, S.T.; Bassler, B.L. Bacterial Quorum Sensing: Its Role in Virulence and Possibilities for Its Control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar]
- Wu, S.; Yang, S.; Wang, M.; Song, N.; Feng, J.; Wu, H.; Yang, A.; Liu, C.; Li, Y.; Guo, F.; et al. Quorum sensing-based interactions among drugs, microbes, and diseases. Sci. China Life Sci. 2022, 66, 137–151. [Google Scholar] [CrossRef]
- Lam, K.N.; Alexander, M.; Turnbaugh, P.J. Precision Medicine Goes Microscopic: Engineering the Microbiome to Improve Drug Outcomes. Cell Host Microbe 2019, 26, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Steiner, H.E.; Patterson, H.K.; Giles, J.B.; Karnes, J.H. Bringing pharmacomicrobiomics to the clinic through well-designed studies. Clin. Transl. Sci. 2022, 15, 2303–2315. [Google Scholar] [CrossRef]
- Van Tran, T.T.; Tayara, H.; Chong, K.T. Artificial Intelligence in Drug Metabolism and Excretion Prediction: Recent Advances, Challenges, and Future Perspectives. Pharmaceutics 2023, 15, 1260. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microbe | Drug | Microbial Metabolite | Metabolic Reaction | Reference |
|---|---|---|---|---|
| Unknown GI bacterial species | Prontosil Neoprontosil | Sulfanilamide | Azo reduction | [30,31] |
| Unknown GI bacterial species | Sulfasalazine Balsalazide Olsalazine | 5-aminosalicylic acid | Azo reduction | [30,32,33,34] |
| Clostridium leptum | Nitrazepam Clonezepam | 7-amino clonazepam | Nitro reduction | [30,39,40] |
| Eubacterium lentum | Digoxin | Dihydrodigoxin | Reduction | [30,40] |
| Unknown GI bacterial species | Loperamide Oxide | Loperamide | N-oxide bond cleavage | [30,35] |
| Eubacterium | Sodium picosulfate | 4,4′-dihydroxy diphenyl-(2-pyridyl)-methane | Hydrolysis | [30,36] |
| Enterobacteriaceae, primarily | Irinotecan | Glucuronidase enzyme | Hydrolysis | [30,37] |
| Bacteroidetes and Clostridium species | Levamisole | Levametabol-I, II, III | Oxidation | [30,31,38] |
| Unknown GI bacterial species | Insulin Calcitonin | Proteolytic enzymes | Peptide degradation | [30,41] |
| Helicobacter pylori | Levodopa | Cell adhesions | Epithelial cell binding | [30,42] |
| Escherichia coli | Baicalin | Baicalein | Hydrolysis | [30,43,44,45] |
| Bifidobacterium bifidum | Hesperidin | Aglycones hesperetin | Hydrolysis | [30,46] |
| Eubacterium rectaleStreptococcus faecium | Daidzin | Daidzein | Hydrolysis | [30,47,48] |
| Microbiota Shift | Drug Type | References |
|---|---|---|
| ↑ (increase) Streptococcus salivarius, Lactobacillaceae, Eubacteriaceae | Opiates | [88,91] |
| ↑ Firmicutes Akkermansia muciniphila, Faecalibacterium prausnitzii ↓ (decrease) Proteobacteria Enterobacteriaceae Desulfovibrio, Prevotella Streptococcus, Collinsella | Atorvastatin | [20,91] |
| ↓ Bacteroidetes | Methotrexate | [89] |
| ↑ Actinomyces Clostridium leptum | L-thyroxine | [88,90] |
| ↑ Streptococcus salivarius Rothia | Oral steroids | [88] |
| ↑ Streptococcus salivarius | Platelet aggregation inhibitors | [88,91] |
| ↑ Streptococcus salivarius | Vitamin D supplements | [23,88] |
| ↑ Haemophilus parainfluenzae | Benzodiazepine | [88] |
| ↑ Bifidobacterium dentium, Streptococcus salivarius | Proton pump inhibitors | [88,91] |
| ↑ Eubacterium ramulus, Streptococcus salivarius | SSRI antidepressants | [88,91] |
| ↑ Clostridium leptum | Tricyclic antidepressants | [88,91] |
| ↑ Alistipes and Bacteroides | Laxatives | [88,91] |
| ↑ Streptococcus mutans Bifidobacterium dentium | Steroid inhalers | [88] |
| ↑ Escherichia coli, Streptococcaceae Akkermansia, Alistipes ↓ Lactonifactor, Odoribacter, Lactococcus, Blautia, Bacteroides | Metformin | [84,88,91] |
| Database/Software Name | Search Result/Prediction | Reference |
|---|---|---|
| Database | ||
| MASI: Microbiota Active-Substance Interaction Database | Information on: Transformation of bioactive substances by gut microbes and vice versa. Disease and microbe interactions. | [93] |
| MagMD: Metabolic action of gut Microbiota to Drugs | Similar information to MASI, but inclusive of the information on enzyme name and effect on drug efficacy. | [94] |
| Prediction Software | ||
| DrugBug | List of microbes with metabolic enzymes suggested to degrade the drug. | [95] |
| Machine learning model | Predicts the effect of administered drugs on the microbiome. Predicts if the drug will be depleted/not depleted. | [96,97] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mousa, S.; Sarfraz, M.; Mousa, W.K. The Interplay between Gut Microbiota and Oral Medications and Its Impact on Advancing Precision Medicine. Metabolites 2023, 13, 674. https://doi.org/10.3390/metabo13050674
Mousa S, Sarfraz M, Mousa WK. The Interplay between Gut Microbiota and Oral Medications and Its Impact on Advancing Precision Medicine. Metabolites. 2023; 13(5):674. https://doi.org/10.3390/metabo13050674
Chicago/Turabian StyleMousa, Sara, Muhammad Sarfraz, and Walaa K. Mousa. 2023. "The Interplay between Gut Microbiota and Oral Medications and Its Impact on Advancing Precision Medicine" Metabolites 13, no. 5: 674. https://doi.org/10.3390/metabo13050674
APA StyleMousa, S., Sarfraz, M., & Mousa, W. K. (2023). The Interplay between Gut Microbiota and Oral Medications and Its Impact on Advancing Precision Medicine. Metabolites, 13(5), 674. https://doi.org/10.3390/metabo13050674