Biliary Amino Acids and Telocytes in Gallstone Disease


Abstract
:1. Introduction
2. Materials and Methods
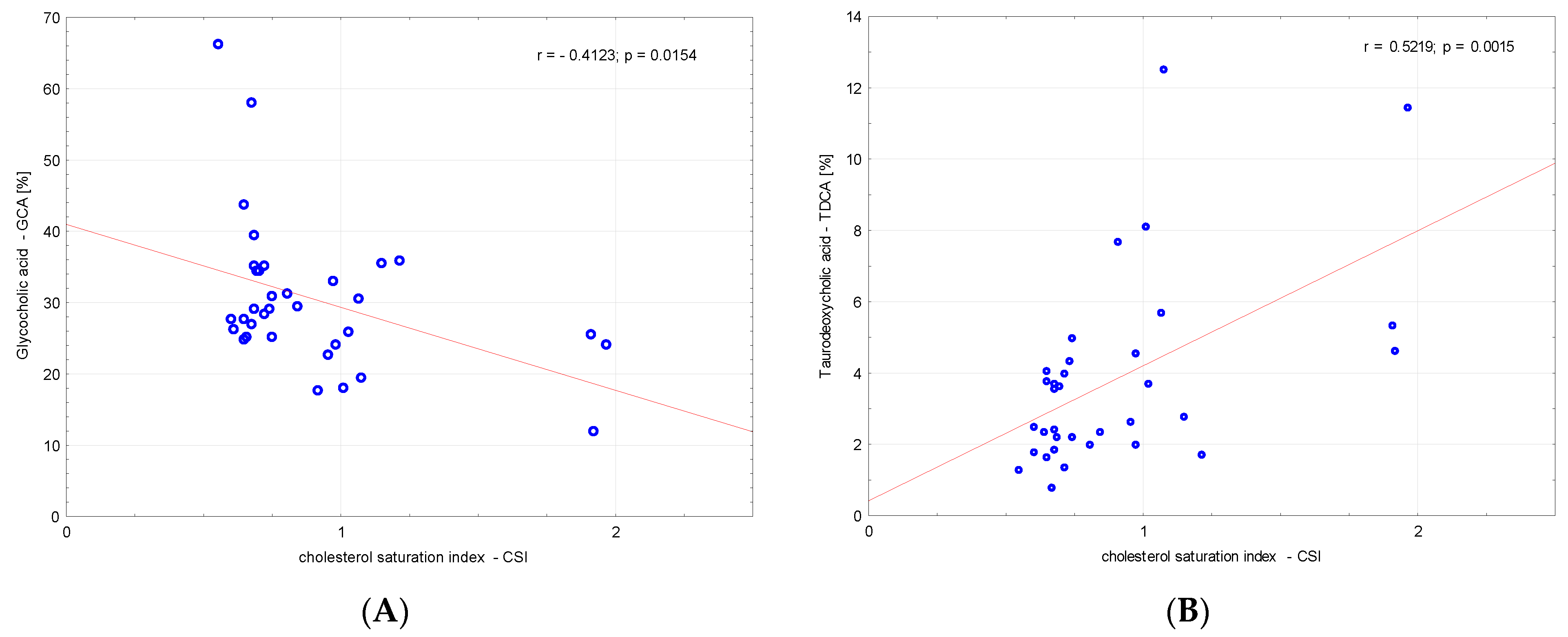
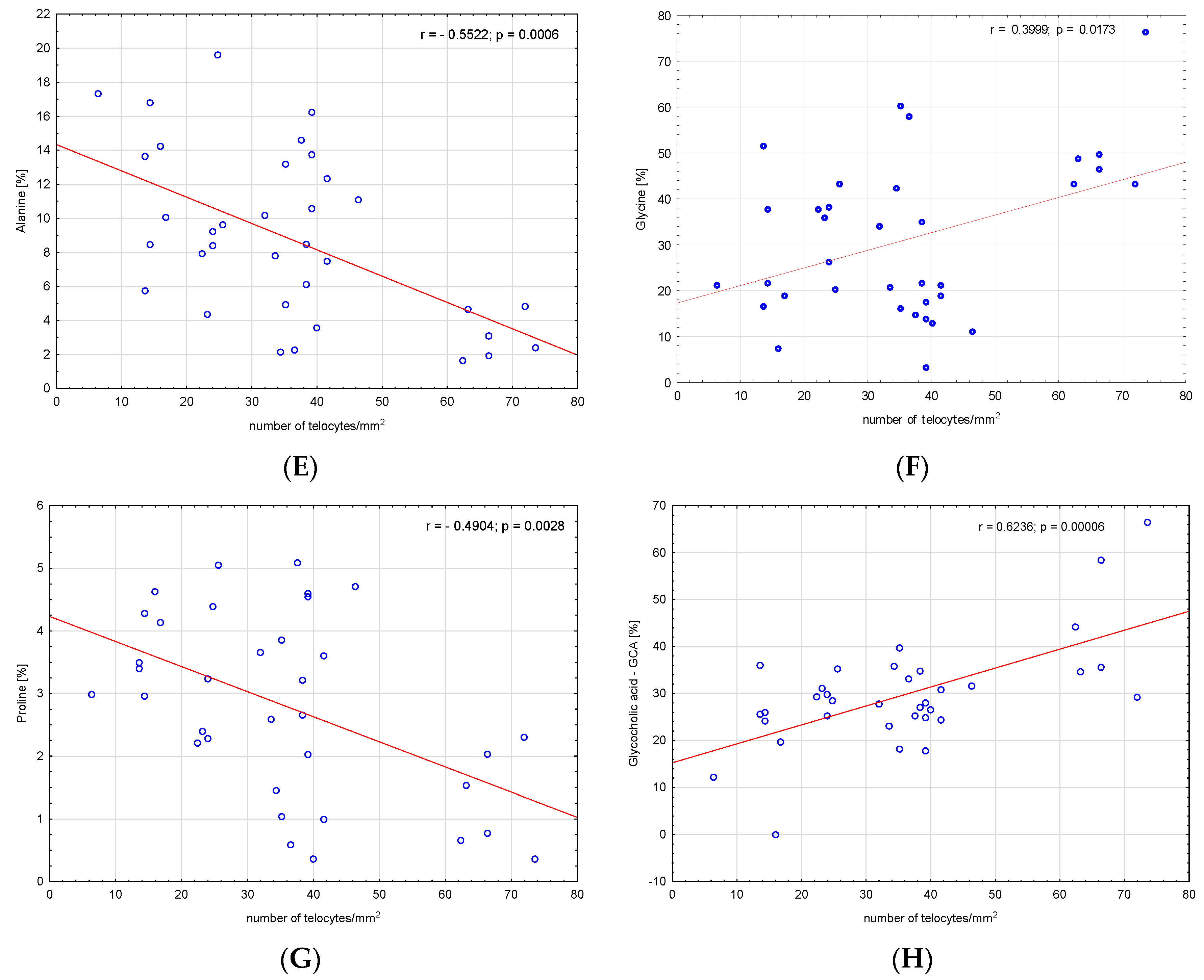
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stinton, L.M.; Shaffer, E.A. Epidemiology of gallbladder disease: Cholelithiasis and cancer. Gut Liver 2012, 6, 172–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratzer, W.; Mason, R.A.; Kachele, V. Prevalence of gallstones in sonographic surveys worldwide. J. Clin. Ultrasound 1999, 27, 1–7. [Google Scholar] [CrossRef]
- Adeva-Andany, M.; Souto-Adeva, G.; Ameneiros-Rodríguez, E.; Fernández-Fernández, C.; Donapetry-García, C.; Domínguez-Montero, A. Insulin resistance and glycine metabolism in humans. Amino Acids 2018, 50, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Monte, M.J.; Marin, J.J.G.; Antelo, A.; Vazquez-Tato, J. Bile acids: Chemistry, physiology, and pathophysiology. World J. Gastroenterol. 2009, 15, 804–816. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Kang, D.-J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.H.; Li, T.; Portincasa, P.; Ford, D.A.; Neuschwander-Tetri, B.A.; Tso, P.; Wang, D.Q.-H. New insights into the role of Lith genes in the formation of cholesterol-supersaturated bile. Liver Res. 2017, 1, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.-C.; Hsi, E.; Lee, K.-T. Mucin genes in gallstone disease. Clin. Chim. Acta 2012, 413, 1466–1471. [Google Scholar] [CrossRef]
- Kuipers, E.J. Encyclopedia of Gastroenterology; Academic Press: Cambridge, MA, USA, 2019; ISBN 012818728X. [Google Scholar]
- Carey, M.C.; Small, D.M. The physical chemistry of cholesterol solubility in bile. Relationship to gallstone formation and dissolution in man. J. Clin. Investig. 1978, 61, 998–1026. [Google Scholar] [CrossRef] [Green Version]
- Carey, M.C. Critical tables for calculating the cholesterol saturation of native bile. J. Clin. Investig. 1978, 19, 945–955. [Google Scholar] [CrossRef]
- Pasternak, A.; Bugajska, J.; Szura, M.; Walocha, J.A.; Matyja, A.; Gajda, M.; Sztefko, K.; Gil, K. Biliary Polyunsaturated Fatty Acids and Telocytes in Gallstone Disease. Cell Transplant. 2017, 26, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Matyja, A.; Gil, K.; Pasternak, A.; Sztefko, K.; Gajda, M.; Tomaszewski, K.A.; Matyja, M.; Walocha, J.A.; Kulig, J.; Thor, P. Telocytes: New insight into the pathogenesis of gallstone disease. J. Cell. Mol. Med. 2013, 17, 734–742. [Google Scholar] [CrossRef] [PubMed]
- Hinescu, M.E.; Ardeleanu, C.; Gherghiceanu, M.; Popescu, L.M. Interstitial Cajal-like cells in human gallbladder. J. Mol. Histol. 2007, 38, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Pasternak, A.; Gil, K.; Matyja, A.; Gajda, M.; Sztefko, K.; Walocha, J.A.; Kulig, J.; Thor, P. Loss of gallbladder interstitial Cajal-like cells in patients with cholelithiasis. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2013, 25, e17–e24. [Google Scholar] [CrossRef]
- Bugajska, J.; Gotfryd-Bugajska, K.; Szura, M.; Berska, J.; Pasternak, A.; Sztefko, K. Characteristics of amino acid profiles and incretin hormones in patients with gallstone disease: A pilot study. Pol. Arch. Intern. Med. 2019, 129, 883–888. [Google Scholar] [CrossRef]
- Bugajska, J.; Berska, J.; Hodorowicz-Zaniewska, D.; Sztefko, K. Walidacja metody oznaczania kwasów tłuszczowych frakcji fosfolipidów w surowicy krwi. Diagnostyka Lab. 2010, 46, 125–130. [Google Scholar]
- Mall, A.S. Analysis of mucins: Role in laboratory diagnosis. J. Clin. Pathol. 2008, 61, 1018–1024. [Google Scholar] [CrossRef]
- Inoue, T.; Mishima, Y. The pathophysiological characteristics of bile from patients with gallstones: The role of prostaglandins and mucin in gallstone formation. Jpn J. Surg. 1990, 20, 10–18. [Google Scholar] [CrossRef]
- Dowling, R.H. Review: Pathogenesis of gallstones. Aliment. Pharmacol. Ther. 2000, 14 (Suppl. S2), 39–47. [Google Scholar] [CrossRef]
- Liu, W.; Liu, J.; Wu, T.; Li, D.; Cui, Y. Therapeutic effect of yinchenhao decoction on cholelithiasis via mucin from gallbladder-intestine. Res. Sq. 2021, in press. [Google Scholar]
- Van Klinken, B.J.; Van Dijken, T.C.; Oussoren, E.; Büller, H.A.; Dekker, J.; Einerhand, A.W. Molecular cloning of human MUC3 cDNA reveals a novel 59 amino acid tandem repeat region. Biochem. Biophys. Res. Commun. 1997, 238, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Toribara, N.W.; Ho, S.B.; Gum, E.; Gum, J.R.J.; Lau, P.; Kim, Y.S. The carboxyl-terminal sequence of the human secretory mucin, MUC6. Analysis Of the primary amino acid sequence. J. Biol. Chem. 1997, 272, 16398–16403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toribara, N.W.; Roberton, A.M.; Ho, S.B.; Kuo, W.L.; Gum, E.; Hicks, J.W.; Gum, J.R.J.; Byrd, J.C.; Siddiki, B.; Kim, Y.S. Human gastric mucin. Identification of a unique species by expression cloning. J. Biol. Chem. 1993, 268, 5879–5885. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-T.; Liu, T.-S. Mucin gene expression in gallbladder epithelium. J. Formos. Med. Assoc. 2002, 101, 762–768. [Google Scholar] [PubMed]
- Hu, F.-L.; Chen, H.-T.; Guo, F.-F.; Yang, M.; Jiang, X.; Yu, J.-H.; Zhang, F.-M.; Xu, G.-Q. Biliary microbiota and mucin 4 impact the calcification of cholesterol gallstones. Hepatobiliary Pancreat. Dis. Int. 2021, 20, 61–66. [Google Scholar] [CrossRef]
- Lagow, E.; DeSouza, M.M.; Carson, D.D. Mammalian reproductive tract mucins. Hum. Reprod. Update 1999, 5, 280–292. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.S.; Gum, J.R.J.; Byrd, J.C.; Toribara, N.W. The structure of human intestinal apomucins. Am. Rev. Respir. Dis. 1991, 144, S10–S14. [Google Scholar] [CrossRef]
- Shimizu, S.; Sabsay, B.; Veis, A.; Ostrow, J.D.; Rege, R.V.; Dawes, L.G. Isolation of an acidic protein from cholesterol gallstones, which inhibits the precipitation of calcium carbonate in vitro. J. Clin. Investig. 1989, 84, 1990–1996. [Google Scholar] [CrossRef] [Green Version]
- Peters, W.H.M.; van Schaik, A.; Peters, J.H.; van Goor, H. Oxidised- and total non-protein bound glutathione and related thiols in gallbladder bile of patients with various gastrointestinal disorders. BMC Gastroenterol. 2007, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Bannai, S. Exchange of cystine and glutamate across plasma membrane of human fibroblasts. J. Biol. Chem. 1986, 261, 2256–2263. [Google Scholar] [CrossRef]
- Combs, J.A.; DeNicola, G.M. The Non-Essential Amino Acid Cysteine Becomes Essential for Tumor Proliferation and Survival. Cancers 2019, 11, 678. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Wang, Y.; Zhu, S.; Bao, X.; Fu, Z.; Zhen, T.; Yuan, Z.; Li, Q.; Deng, Z.; Sun, J.; et al. Changes in plasma bile acids are associated with gallbladder stones and polyps. BMC Gastroenterol. 2020, 20, 363. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | Control Group | Study Group | |
|---|---|---|---|
| Percentage of Total FAs Mean (SD) or Median (Interquartile Range) | p | ||
| SFAs | |||
| C 12 | 0.03 (0.02–0.04) | 0.03 (0.02–0.04) | 0.72 |
| C 14 | 0.45 (0.20) | 0.51 (0.18) | 0.43 |
| C 16 | 42.5 (40.9–43.5) | 40.4 (40.0–41.2) | 0.04 |
| C 18 | 5.91 (5.04–6.57) | 6.13 (5.43–6.80) | 0.61 |
| MUFAs | |||
| C 16:1 (n-7) | 2.42 (1.22) | 2.10 (0.97) | 0.41 |
| C 18:1 (n-9) | 12.2 (10.3–13.6) | 11.2 (9.9–13.0) | 0.39 |
| PUFAs | |||
| C 18:2 (n-6) | 23.4 (22.2–25.5) | 28.8 (25.8–31.2) | 0.008 |
| C 18:3 (n-3) | 0.39 (0.20) | 0.58 (0.21) | 0.01 |
| C 20:2 (n-6) | 0.14 (0.10–0.15) | 0.16 (0.13–0.20) | 0.15 |
| C 20:4 (n-6) | 6.77 (3.64) | 6.54 (1.28) | 0.78 |
| C 20:5 (n-3) | 1.26 (1.03) | 1.15 (0.61) | 0.68 |
| C 22:6 (n-3) | 2.26 (0.79) | 1.97 (0.65) | 0.26 |
| C 20 + C 18:3 (n-6) | 0.25 (0.15–0.50) | 0.28 (0.20–0.40) | 0.77 |
| Bile Acids | Control Group | Study Group | |
|---|---|---|---|
| Percentage of Total BAs Mean (SD) or Median (Interquartile Range) | p | ||
| Primary bile acids | |||
| GCA | 33.8 (27.8–39.9) | 27.0 (24.2–30.8) | 0.03 |
| TCA | 10.4 (6.42–19.1) | 7.57 (5.59–10.7) | 0.22 |
| GCDCA | 19.9 (16.5–29.6) | 25.8 (22.5–29.7) | 0.16 |
| TCDCA | 10.8 (7.0) | 8.6 (4.6) | 0.22 |
| Secondary bile acids | |||
| GDCA | 13.7 (9.9) | 23.4 (8.9) | 0.001 |
| TDCA | 2.02 (1.74–3.21) | 3.9 (2.4–5.0) | 0.03 |
| Amino Acids | Control Group | Study Group | |
|---|---|---|---|
| Percentage of Total AAs Mean (SD) or Median (Interquartile Range) | p | ||
| Valine | 0.93 (0.41–2.92) | 3.38 (1.92–4.51) | 0.01 |
| Isoleucine | 0.89 (0.46–2.65) | 2.01 (1.50–2.56) | 0.03 |
| Leucine | 0.66 (0.45–4.97) | 2.68 (1.81–3.85) | 0.06 |
| Threonine | 1.89 (1.56) | 2.97 (0.92) | 0.01 |
| Methionine | 0.02 (0.01–0.04) | 0.05 (0.03–0.19) | 0.02 |
| Phenylalanine | 0.30 (0.11–1.16) | 1.09 (0.68–1.50) | 0.045 |
| Lysine | 4.11 (2.87) | 4.98 (2.01) | 0.30 |
| Tryptophan | 0.38 (0.20) | 0.48 (0.24) | 0.18 |
| Histidine | 0.78 (0.49–1.13) | 0.67 (0.33–0.94) | 0.46 |
| Arginine | 2.15 (1.70–3.17) | 1.60 (1.11–2.27) | 0.10 |
| Tyrosine | 0.45 (0.32–0.90) | 1.09 (0.74–1.71) | 0.01 |
| Aspartic acid | 1.65 (0.85–1.89) | 2.08 (1.45–2.35) | 0.09 |
| Glutamic acid | 5.10 (2.48) | 11.4 (3.45) | <0.001 |
| Serine | 2.37 (1.96) | 4.03 (1.20) | 0.004 |
| Asparagine | 0.08 (0.00–0.36) | 0.16 (0.05–0.43) | 0.45 |
| Glycine | 38.8 (20.4) | 27.0 (13.8) | 0.05 |
| Taurine | 15.2 (9.7–25.4) | 10.0 (6.09–19.0) | 0.14 |
| Citrulline | 0.01 (0.00–0.19) | 0.10 (0.01–0.19) | 0.13 |
| Alanine | 3.3 (2.18–6.14) | 10.0 (7.9–13.7) | <0.001 |
| Proline | 1.22 (0.62–2.02) | 3.49 (2.65–4.39) | <0.001 |
| Ornithine | 3.31 (2.10–4.62) | 3.04 (2.36–4.16) | 0.79 |
| 3-methyl-histidine | 0.05 (0.02–0.07) | 0.03 (0.03–0.06) | 0.93 |
| Cystine | 2.86 (1.84–5.96) | 1.04 (0.59–1.97) | 0.003 |
| α-Aminobutyric acid | 0.32 (0.20–0.39) | 0.23 (0.12–0.35) | 0.22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bugajska, J.; Berska, J.; Pasternak, A.; Sztefko, K. Biliary Amino Acids and Telocytes in Gallstone Disease. Metabolites 2023, 13, 753. https://doi.org/10.3390/metabo13060753
Bugajska J, Berska J, Pasternak A, Sztefko K. Biliary Amino Acids and Telocytes in Gallstone Disease. Metabolites. 2023; 13(6):753. https://doi.org/10.3390/metabo13060753
Chicago/Turabian StyleBugajska, Jolanta, Joanna Berska, Artur Pasternak, and Krystyna Sztefko. 2023. "Biliary Amino Acids and Telocytes in Gallstone Disease" Metabolites 13, no. 6: 753. https://doi.org/10.3390/metabo13060753
APA StyleBugajska, J., Berska, J., Pasternak, A., & Sztefko, K. (2023). Biliary Amino Acids and Telocytes in Gallstone Disease. Metabolites, 13(6), 753. https://doi.org/10.3390/metabo13060753