


The Effects of a High Concentration of Dissolved Oxygen on Actinobacteria from Lake Baikal

 , , , , , , , and
, , , , , , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sampling, Location, and Isolation of Actinobacteria
2.3. 16S rRNA Gene Sequencing and Analysis
2.4. Cultivation of Actinobacteria
2.5. Microscopy of the Strains
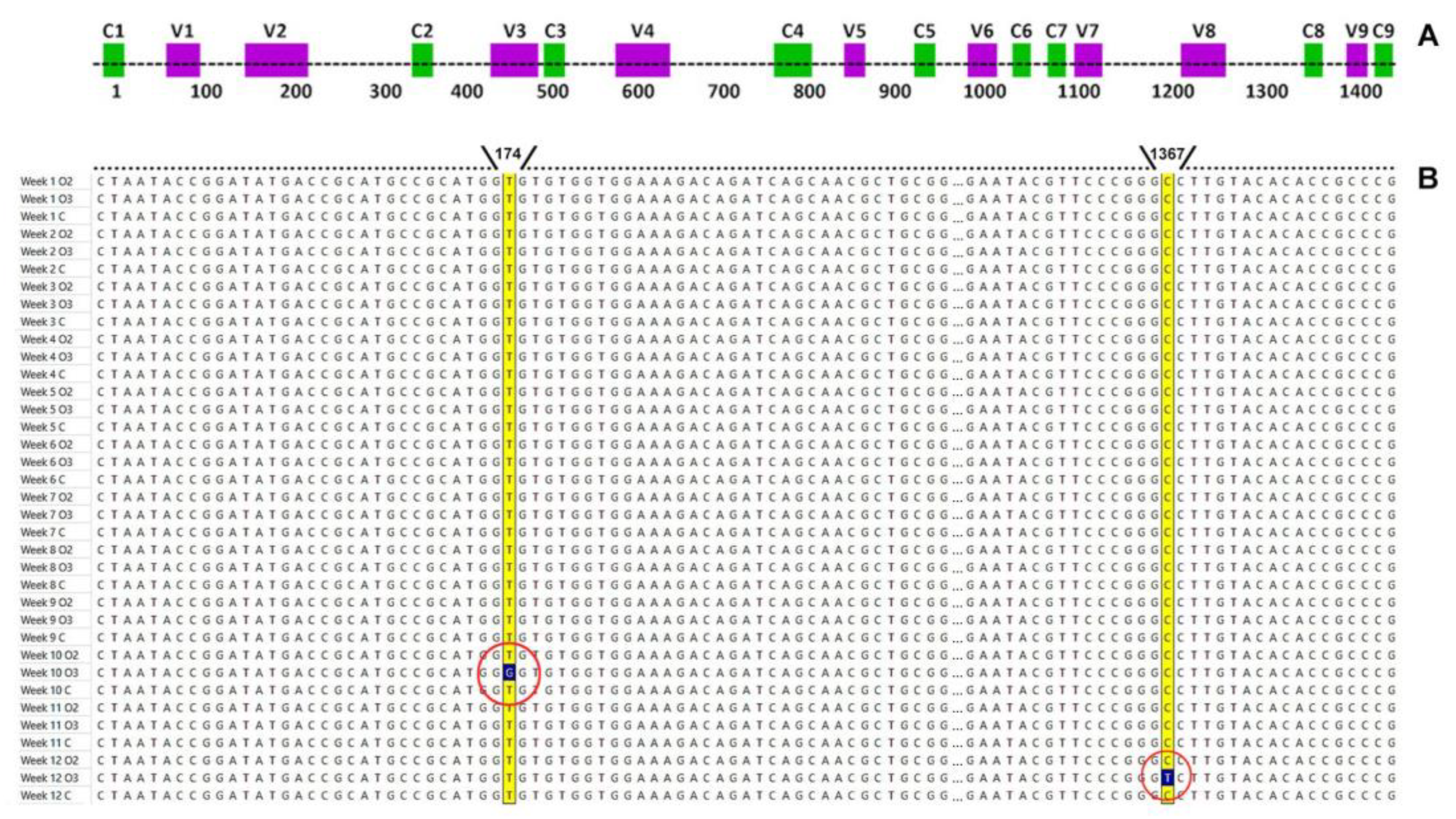
2.6. Searching for DNA Mutations
2.7. Extraction of Secondary Metabolites
2.8. Antioxidant Activity Assay of Extracts from Isolated Strains
2.9. Antibiotic Assay of Extracts from Isolated Strains
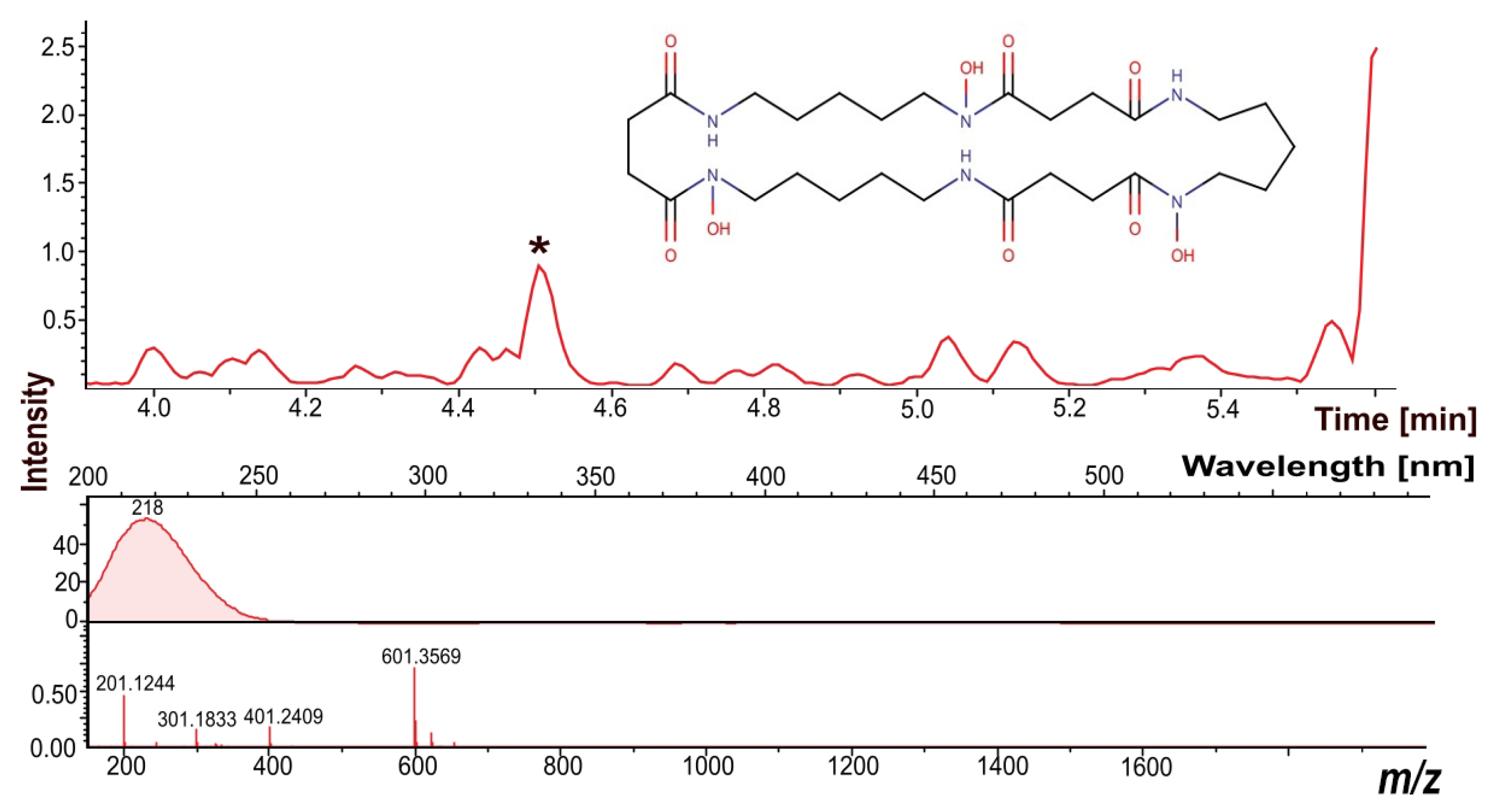
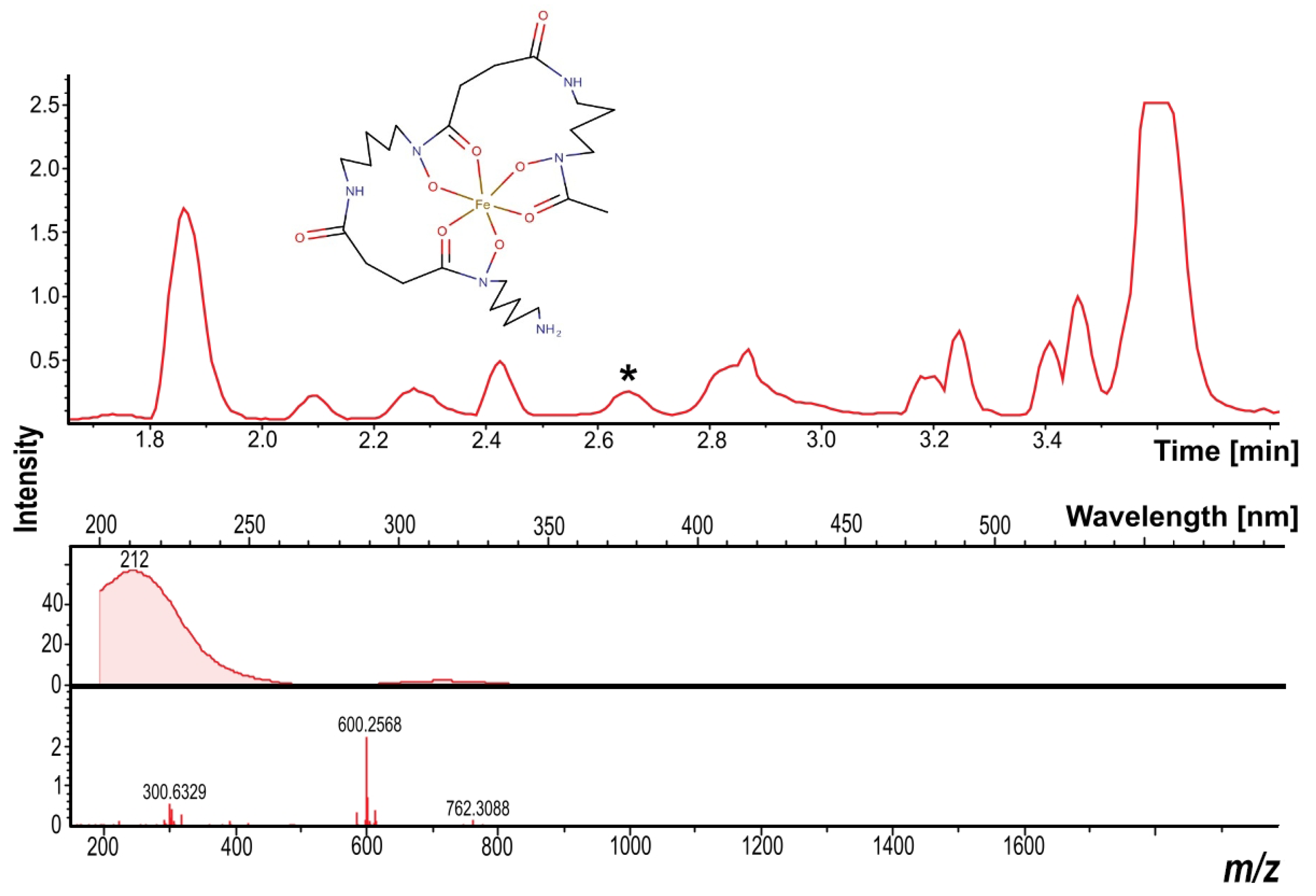
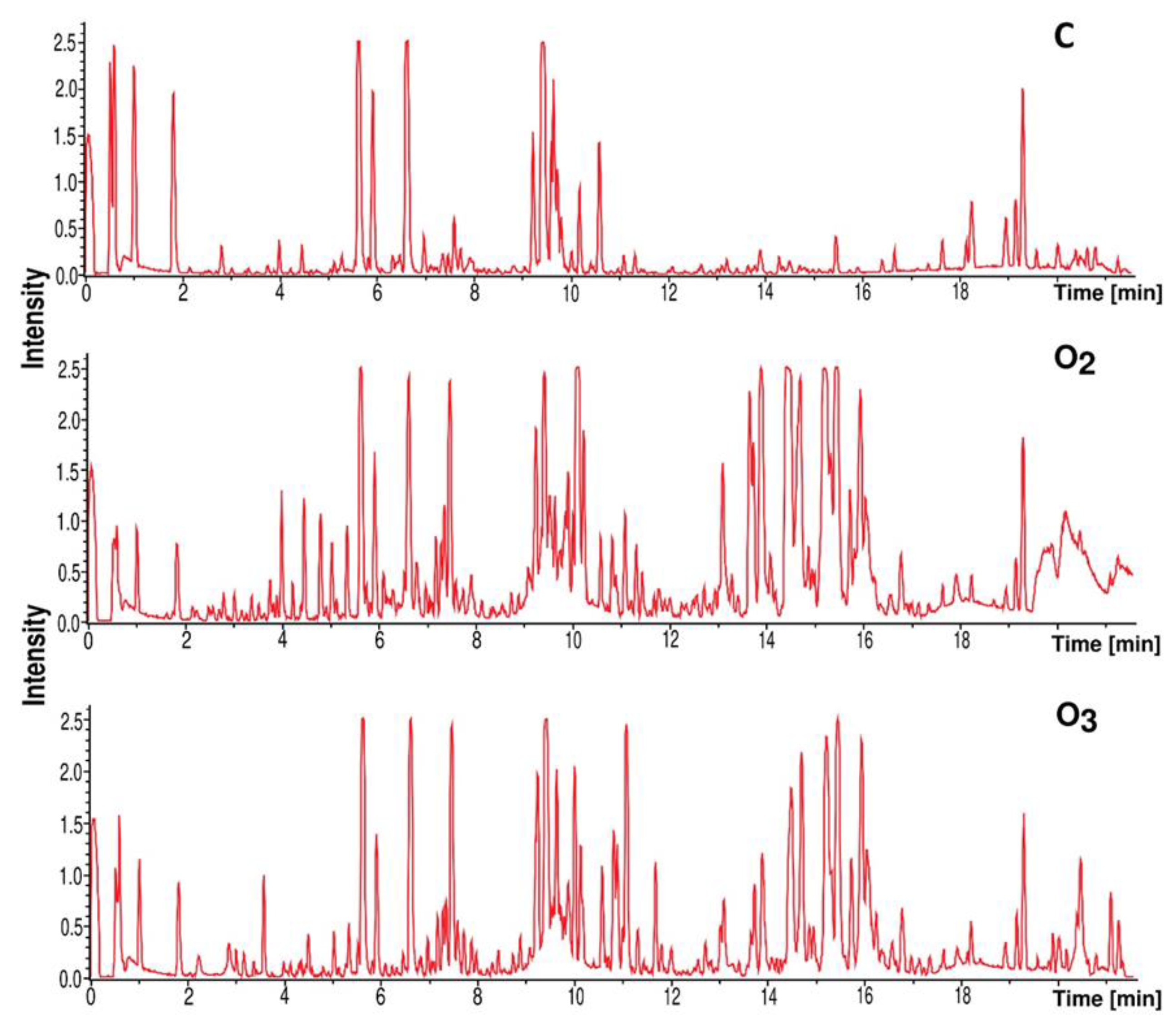
2.10. Liquid Chromatography–Mass Spectrometry (LC–MS) and Dereplication Analysis
3. Results
3.1. Isolation of Actinobacteria in Standard and Experimental Conditions
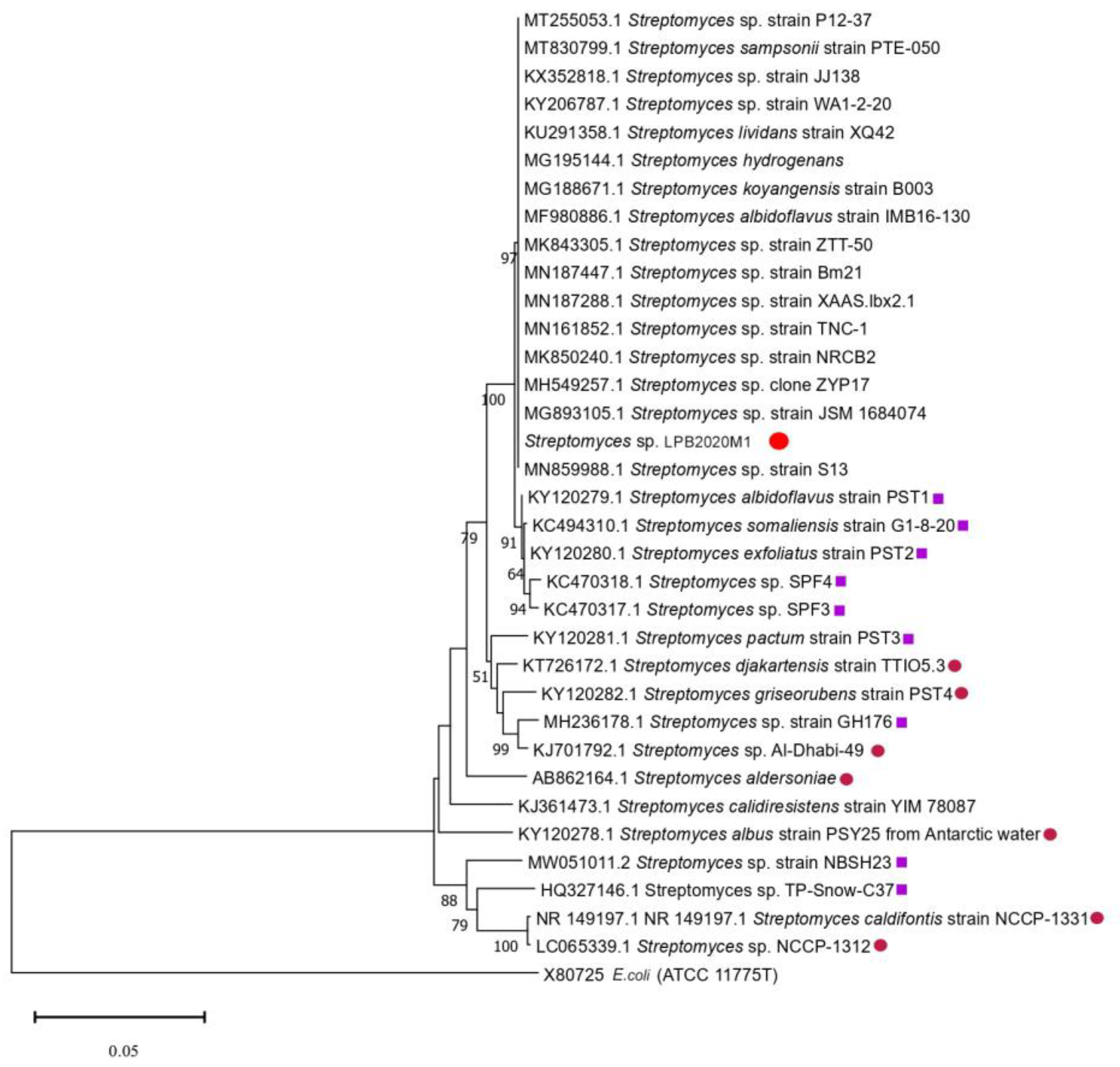
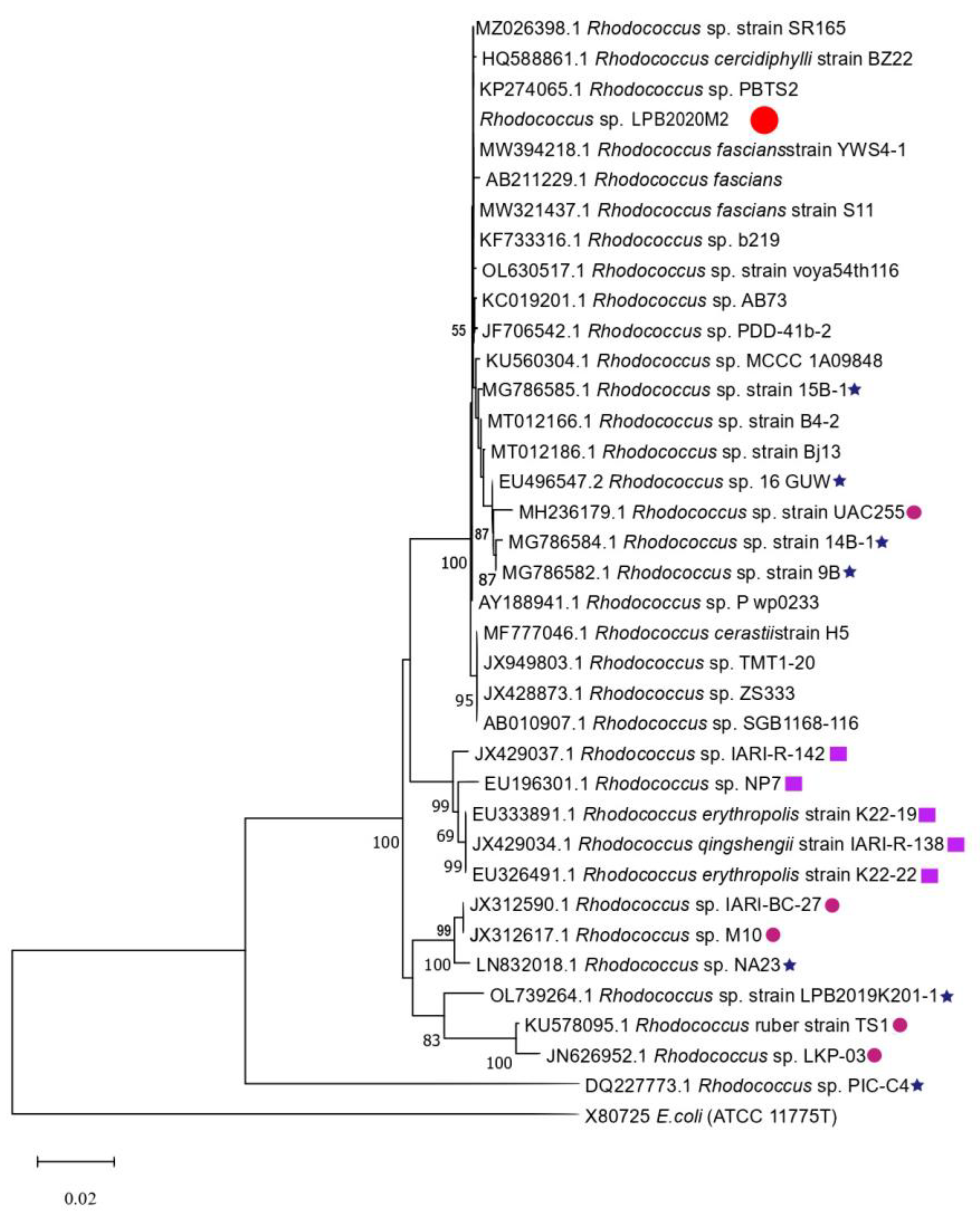
3.2. 16S rRNA Gene Sequencing and Analysis
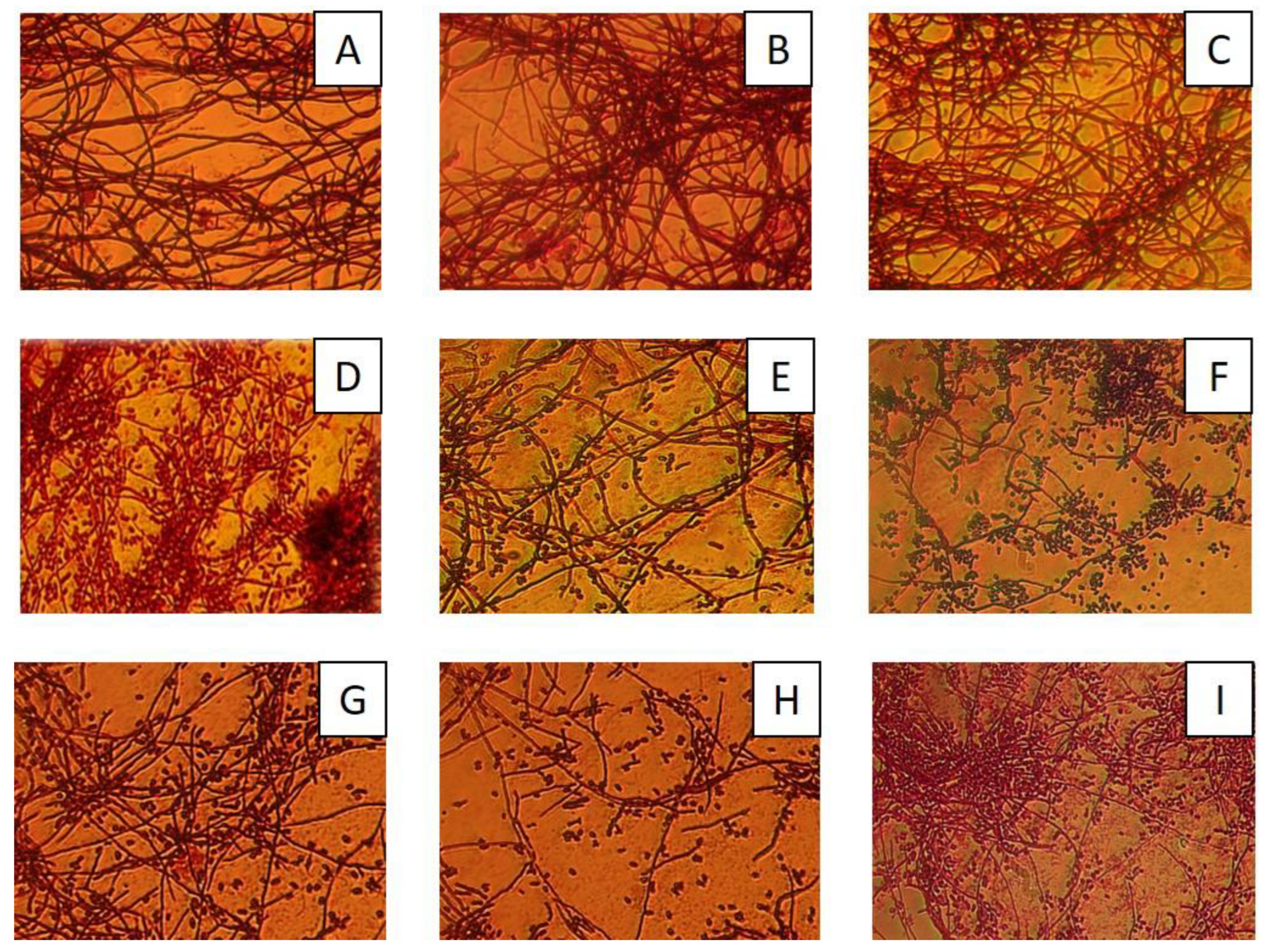
3.3. Assessment of Morphological Changes
3.4. DNA Mutations in 16S rRNA Gene
3.5. Antioxidant Activity of Extracts from Isolated Strains
3.6. Antibiotic Activity of Extracts from Isolated Strains
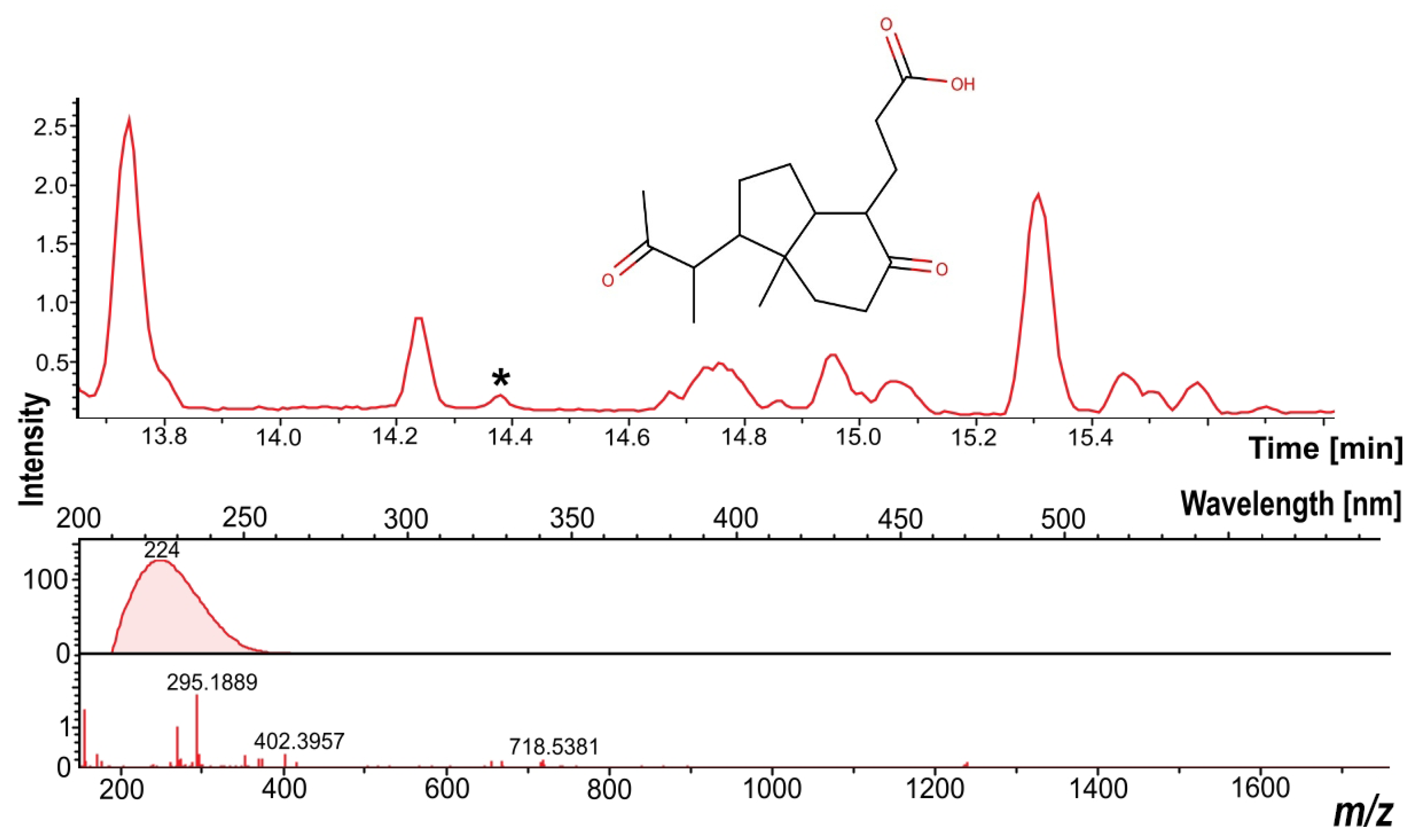
3.7. Dereplication of the Secondary Metabolites
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kebede, G.; Tafese, T.; Abda, E.M.; Kamaraj, M.; Assefa, F. Factors Influencing the Bacterial Bioremediation of Hydrocarbon Contaminants in the Soil: Mechanisms and Impacts. J. Chem. 2021, 2021, 9823362. [Google Scholar] [CrossRef]
- Taverne, Y.J.; Merkus, D.; Bogers, A.J.; Halliwell, B.; Duncker, D.J.; Lyons, T.W. Reactive Oxygen Species: Radical Factors in the Evolution of Animal Life. BioEssays 2018, 40, 1700158. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, J.B.; Rosen, P.C.; Sprenger, H.-G.; Puszynska, A.M.; Mann, J.L.; Roessler, J.M.; Cangelosi, A.L.; Henne, A.; Condon, K.J.; Zhang, T.; et al. Fumarate Is a Terminal Electron Acceptor in the Mammalian Electron Transport Chain. Science 2021, 374, 1227–1237. [Google Scholar] [CrossRef] [PubMed]
- Nelson, H.R.; Altieri, A.H. Oxygen: The Universal Currency on Coral Reefs. Coral Reefs 2019, 38, 177–198. [Google Scholar] [CrossRef]
- Sekretaryova, A.; Jones, S.M.; Solomon, E.I. O2 Reduction to Water by High Potential Multicopper Oxidases: Contributions of the T1 Copper Site Potential and the Local Environment of the Trinuclear Copper Cluster. J. Am. Chem. Soc. 2019, 141, 11304–11314. [Google Scholar] [CrossRef]
- Sies, H. Hydrogen Peroxide as a Central Redox Signaling Molecule in Physiological Oxidative Stress: Oxidative Eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [Green Version]
- Kumari, S.; Sharma, S.; Advani, D.; Khosla, A.; Kumar, P.; Ambasta, R.K. Unboxing the Molecular Modalities of Mutagens in Cancer. Env. Sci. Pollut. Res. 2022, 29, 62111–62159. [Google Scholar] [CrossRef]
- Bhagwat, A.S.; Hao, W.; Townes, J.P.; Lee, H.; Tang, H.; Foster, P.L. Strand-Biased Cytosine Deamination at the Replication Fork Causes Cytosine to Thymine Mutations in Escherichia coli. Proc. Natl. Acad. Sci. USA 2016, 113, 2176–2181. [Google Scholar] [CrossRef]
- Srinivas, U.S.; Tan, B.W.Q.; Vellayappan, B.A.; Jeyasekharan, A.D. ROS and the DNA Damage Response in Cancer. Redox Biol. 2019, 25, 101084. [Google Scholar] [CrossRef] [PubMed]
- Hahm, J.Y.; Park, J.; Jang, E.S.; Chi, S.W. 8-Oxoguanine: From Oxidative Damage to Epigenetic and Epitranscriptional Modification. Exp. Mol. Med. 2022, 54, 1626–1642. [Google Scholar] [CrossRef] [PubMed]
- Belykh, I.A.; Zinchenko, V.D.; Vysekantsev, I.P. Stimulating Effect of Ozone Low Doses on Microorganisms’ Growth. Theor. Exp. Cryobiol. 2004, 4, 41–45. [Google Scholar]
- Mitsugi, F.; Nagahama, K.; Ebihara, K. Influence of Ozone Generated by Atmospheric Pressure Plasma on Microorganisms and DNA in Soil. J. Biomater. Tissue Eng. 2018, 8, 915–918. [Google Scholar] [CrossRef]
- Axenov-Gribanov, D.V.; Morgunova, M.M.; Vasilieva, U.A.; Gamaiunov, S.V.; Dmitrieva (Krasnova), M.E.; Pereliaeva, E.V.; Belyshenko, A.Y.; Luzhetskyy, A.N. Composition of Nutrient Media and Temperature of Cultivation Imposes Effect on the Content of Secondary Metabolites of Nocardiopsis Sp. Isolated from a Siberian Cave. 3 Biotech 2021, 11, 386. [Google Scholar] [CrossRef]
- Genilloud, O. Actinomycetes: Still a Source of Novel Antibiotics. Nat. Prod. Rep. 2017, 34, 1203–1232. [Google Scholar] [CrossRef]
- Manivasagan, P.; Kang, K.H.; Sivakumar, K.; Li-Chan, E.C.Y.; Oh, H.M.; Kim, S.K. Marine Actinobacteria: An Important Source of Bioactive Natural Products. Environ. Toxicol. Pharmacol. 2014, 38, 172–188. [Google Scholar] [CrossRef]
- Selim, M.S.M.; Abdelhamid, S.A.; Mohamed, S.S. Secondary Metabolites and Biodiversity of Actinomycetes. J. Genet. Eng. Biotechnol. 2021, 19, 72. [Google Scholar] [CrossRef]
- Silva, G.d.C.; Kitano, I.T.; Ribeiro, I.A.d.F.; Lacava, P.T. The Potential Use of Actinomycetes as Microbial Inoculants and Biopesticides in Agriculture. Front. Soil Sci. 2022, 2. [Google Scholar] [CrossRef]
- Malavasi, V.; Soru, S.; Cao, G. Extremophile Microalgae: The Potential for Biotechnological Application. J. Phycol. 2020, 56, 559–573. [Google Scholar] [CrossRef]
- Khodzher, T.V.; Domysheva, V.M.; Sorokovikova, L.M.; Sakirko, M.V.; Tomberg, I.V. Current Chemical Composition of Lake Baikal Water. Inland Waters 2017, 7, 250–258. [Google Scholar] [CrossRef]
- Timoshkin, O.A.; Samsonov, D.P.; Yamamuro, M.; Moore, M.V.; Belykh, O.I.; Malnik, V.V.; Sakirko, M.V.; Shirokaya, A.A.; Bondarenko, N.A.; Domysheva, V.M.; et al. Rapid Ecological Change in the Coastal Zone of Lake Baikal (East Siberia): Is the Site of the World’s Greatest Freshwater Biodiversity in Danger? J. Great Lakes Res. 2016, 42, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Axenov-Gribanov, D.V.; Kostka, D.V.; Vasilieva, U.A.; Shatilina, Z.M.; Krasnova, M.E.; Pereliaeva, E.V.; Zolotovskaya, E.D.; Morgunova, M.M.; Rusanovskaya, O.O.; Timofeyev, M.A. Cultivable Actinobacteria First Found in Baikal Endemic Algae Is a New Source of Natural Products with Antibiotic Activity. Int. J. Microbiol. 2020, 2020, e5359816. [Google Scholar] [CrossRef]
- Cabello-Yeves, P.J.; Zemskaya, T.I.; Zakharenko, A.S.; Sakirko, M.V.; Ivanov, V.G.; Ghai, R.; Rodriguez-Valera, F. Microbiome of the Deep Lake Baikal, a Unique Oxic Bathypelagic Habitat. Limnol. Oceanogr. 2020, 65, 1471–1488. [Google Scholar] [CrossRef] [Green Version]
- Chernitsyna, S.M.; Khalzov, I.A.; Sitnikova, T.Y.; Naumova, T.V.; Khabuev, A.V.; Zemskaya, T.I. Microbial Communities Associated with Bentic Invertebrates of Lake Baikal. Curr. Microbiol. 2021, 78, 3020–3031. [Google Scholar] [CrossRef] [PubMed]
- Galachyants, A.D.; Krasnopeev, A.Y.; Podlesnaya, G.V.; Potapov, S.A.; Sukhanova, E.V.; Tikhonova, I.V.; Zimens, E.A.; Kabilov, M.R.; Zhuchenko, N.A.; Gorshkova, A.S.; et al. Diversity of Aerobic Anoxygenic Phototrophs and Rhodopsin-Containing Bacteria in the Surface Microlayer, Water Column and Epilithic Biofilms of Lake Baikal. Microorganisms 2021, 9, 842. [Google Scholar] [CrossRef]
- Lipko, I.A.; Krasnopeev, A.Y.; Tikhonova, I.V.; Timoshkin, O.A.; Kabilov, M.R.; Belykh, O.I. Genetic Diversity of Actinobacteria Inhabiting Water and Sponges of Lake Baikal. Limnol. Freshw. Biol. 2020, 4, 998–999. [Google Scholar] [CrossRef]
- Kurilkina, M.I.; Zakharova, Y.R.; Galachyants, Y.P.; Petrova, D.P.; Bukin, Y.S.; Domysheva, V.M.; Blinov, V.V.; Likhoshway, Y.V. Bacterial Community Composition in the Water Column of the Deepest Freshwater Lake Baikal as Determined by Next-Generation Sequencing. FEMS Microbiol. Ecol. 2016, 92, fiw094. [Google Scholar] [CrossRef] [Green Version]
- Kozhov, M. Lake Baikal and Its Life; Monographiae Biologicae; Springer Netherlands: Dordrecht, The Netherlands, 1963; Volume 11, ISBN 978-94-015-7390-0. [Google Scholar]
- Galaziy, G.I. Baikal in Questions and Answers; East-Siberian Publishing House: Irkutsk, Russia, 1984. (In Russian) [Google Scholar]
- Kieser, T. Practical Streptomyces Genetics; John Innes Foundation: Norwich, UK, 2000; ISBN 978-0-7084-0623-6. [Google Scholar]
- Pereliaeva, E.V.; Dmitrieva, M.E.; Morgunova, M.M.; Belyshenko, A.Y.; Imidoeva, N.A.; Ostyak, A.S.; Axenov-Gribanov, D.V. The Use of Baikal Psychrophilic Actinobacteria for Synthesis of Biologically Active Natural Products from Sawdust Waste. Fermentation 2022, 8, 213. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Moyes, R.B.; Reynolds, J.; Breakwell, D.P. Differential Staining of Bacteria: Gram Stain. Curr. Protoc. Microbiol. 2009, 15, A.3C.1–A.3C.8. [Google Scholar] [CrossRef] [PubMed]
- Sarker, S.D.; Nahar, L. (Eds.) Natural Products Isolation; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2012; Volume 864, ISBN 978-1-61779-623-4. [Google Scholar]
- Kansci, G.; Dongo, E.; Genot, C. 2,2-Diphenyl-1-Picrylhydrazyl (DPPH∗) Test Demonstrates Antiradical Activity of Dorstenia Psilurus and Dorstenia Ciliata Plant Extracts. Food Nahr. 2003, 47, 434–437. [Google Scholar] [CrossRef] [PubMed]
- Ruangpan, L.; Tendencia, E. Laboratory Manual of Standardized Methods for Antimicrobial Sensitivity Tests for Bacteria Isolated from Aquatic Animals and Environment; Aquaculture Department, Southeast Asian Fisheries Development Center: Quezon City, Philippines, 2004; ISBN 978-971-8511-74-9. [Google Scholar]
- Pinheiro, F.C.; Bortolotto, V.C.; Araujo, S.M.; Poetini, M.R.; Sehn, C.P.; Neto, J.S.S.; Zeni, G.; Prigol, M. Antimicrobial Effect of 2-Phenylethynyl-Butyltellurium in Escherichia Coli and Its Association with Oxidative Stress. J. Microbiol. Biotechnol. 2018, 28, 1209–1216. [Google Scholar] [CrossRef]
- Bhargav, H.S.; Shastri, S.D.; Poornav, S.P.; Darshan, K.M.; Nayak, M.M. Measurement of the Zone of Inhibition of an Antibiotic. In Proceedings of the 2016 IEEE 6th International Conference on Advanced Computing (IACC), Andhra Pradesh, India, 27–28 February 2016; pp. 409–414. [Google Scholar]
- Błajet-Kosicka, A.; Kosicki, R.; Twarużek, M.; Grajewski, J. Determination of Moulds and Mycotoxins in Dry Dog and Cat Food Using Liquid Chromatography with Mass Spectrometry and Fluorescence Detection. Food Addit. Contam. Part B Surveill. 2014, 7, 302–308. [Google Scholar] [CrossRef]
- Ram, J.L.; Karim, A.S.; Sendler, E.D.; Kato, I. Strategy for Microbiome Analysis Using 16S RRNA Gene Sequence Analysis on the Illumina Sequencing Platform. Syst. Biol. Reprod. Med. 2011, 57, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Ueki, M.; Suzuki, R.; Takamatsu, S.; Takagi, H.; Uramoto, M.; Ikeda, H.; Osada, H. Nocardamin Production by Streptomyces Avermitilis. Actinomycetologica 2009, 23, 34–39. [Google Scholar] [CrossRef] [Green Version]
- Bu, Z.Q.; Yu, H.Y.; Wang, J.; He, X.; Cui, Y.R.; Feng, J.C.; Feng, J. Emerging Role of Ferroptosis in the Pathogenesis of Ischemic Stroke: A New Therapeutic Target? ASN Neuro 2021, 13, 175909142110375. [Google Scholar] [CrossRef]
- Köster, A.; Thol, M.; Vrabec, J. Molecular Models for the Hydrogen Age: Hydrogen, Nitrogen, Oxygen, Argon, and Water. J. Chem. Eng. Data 2018, 63, 305–320. [Google Scholar] [CrossRef]
- Karlov, D.S.; Marie, D.; Sumbatyan, D.A.; Chuvochina, M.S.; Kulichevskaya, I.S.; Alekhina, I.A.; Bulat, S.A. Microbial Communities within the Water Column of Freshwater Lake Radok, East Antarctica: Predominant 16S RDNA Phylotypes and Bacterial Cultures. Polar Biol. 2017, 40, 823–836. [Google Scholar] [CrossRef]
- Karlov, D.S.; Marie, D.; Chuvochina, M.S.; Alekhina, I.A.; Bulat, S.A. Microbial communities of water column of Lake Radok, East Antarctica, dominated by abundant actinobacterium “Candidatus Planktophila limnetica”. Mikrobiologiia 2011, 80, 571–574. [Google Scholar] [CrossRef] [PubMed]
- Epova, E.Y.; Shevelev, A.B.; Akbayev, R.M.; Biryukova, Y.K.; Zylkova, M.V.; Bogdanova, E.S.; Guseva, M.A.; Tynio, Y.Y.; Egorov, V.V. Heterotrophic Microbiota from the Oligotrophic Waters of Lake Vostok, Antarctica. Int. J. Environ. Res. Public Health 2022, 19, 4025. [Google Scholar] [CrossRef] [PubMed]
- Parfenova, V.V.; Gladkikh, A.S.; Belykh, O.I. Comparative Analysis of Biodiversity in the Planktonic and Biofilm Bacterial Communities in Lake Baikal. Microbiology 2013, 82, 91–101. [Google Scholar] [CrossRef]
- Western Society of Naturalists. Contributions to Marine Biology: Lectures and Symposia Given at the Hopkins Marine Station, December 20–21, 1929, at the Midwinter Meeting of the Western Society of Naturalists; Stanford University Press: Redwood City, CA, USA, 1931. [Google Scholar]
- Daniel, Q.; Udelsman, R. (Eds.) Surgery of the Thyroid and Parathyroid Glands; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Abdelmohsen, U.R.; Szesny, M.; Othman, E.M.; Schirmeister, T.; Grond, S.; Stopper, H.; Hentschel, U. Antioxidant and Anti-Protease Activities of Diazepinomicin from the Sponge-Associated Micromonospora Strain RV115. Mar. Drugs 2012, 10, 2208–2221. [Google Scholar] [CrossRef]
- Zhao, Y.-X.; Chen, S.-R.; Su, P.-P.; Huang, F.-H.; Shi, Y.-C.; Shi, Q.-Y.; Lin, S. Using Mesenchymal Stem Cells to Treat Female Infertility: An Update on Female Reproductive Diseases. Stem Cells Int. 2019, 2019, 9071720. [Google Scholar] [CrossRef]
- Agarwal, A.; Maldonado Rosas, I.; Anagnostopoulou, C.; Cannarella, R.; Boitrelle, F.; Munoz, L.V.; Finelli, R.; Durairajanayagam, D.; Henkel, R.; Saleh, R. Oxidative Stress and Assisted Reproduction: A Comprehensive Review of Its Pathophysiological Role and Strategies for Optimizing Embryo Culture Environment. Antioxidants 2022, 11, 477. [Google Scholar] [CrossRef]
- Goud, A.P.; Goud, P.T.; Diamond, M.P.; Gonik, B.; Abu-Soud, H.M. Reactive Oxygen Species and Oocyte Aging: Role of Superoxide, Hydrogen Peroxide, and Hypochlorous Acid. Free Radic. Biol. Med. 2008, 44, 1295–1304. [Google Scholar] [CrossRef] [Green Version]
- Ghafarizadeh, A.; Malmir, M.; Naderi Noreini, S.; Faraji, T. Antioxidant Effects of N-Acetylcysteine on the Male Reproductive System: A Systematic Review. Andrologia 2021, 53, e13898. [Google Scholar] [CrossRef]
- Heidari, B.; Mohammadipanah, F. Isolation and Identification of Two Alkaloid Structures with Radical Scavenging Activity from Actinokineospora Sp. UTMC 968, a New Promising Source of Alkaloid Compounds. Mol. Biol. Rep. 2018, 45, 2325–2332. [Google Scholar] [CrossRef]
- Simpson, D.S.A.; Oliver, P.L. ROS Generation in Microglia: Understanding Oxidative Stress and Inflammation in Neurodegenerative Disease. Antioxidants 2020, 9, 743. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.D.; Zhao, X.; Li, Y.; Li, G.R.; Liu, X.L. Damage to Dopaminergic Neurons by Oxidative Stress in Parkinson’s Disease (Review). Int. J. Mol. Med. 2018, 41, 1817–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winiarska-Mieczan, A.; Baranowska-Wójcik, E.; Kwiecień, M.; Grela, E.R.; Szwajgier, D.; Kwiatkowska, K.; Kiczorowska, B. The Role of Dietary Antioxidants in the Pathogenesis of Neurodegenerative Diseases and Their Impact on Cerebral Oxidoreductive Balance. Nutrients 2020, 12, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bignell, D.R.D.; Warawa, J.L.; Strap, J.L.; Chater, K.F.; Leskiw, B.K. Study of the BldG Locus Suggests That an Anti-Anti-Sigma Factor and an Anti-Sigma Factor May Be Involved in Streptomyces Coelicolor Antibiotic Production and Sporulation. The GenBank Accession Numbers for the Sequences Reported in This Paper Are AF134889 and AL035636. Microbiology 2000, 146, 2161–2173. [Google Scholar] [CrossRef] [Green Version]
- Taylor, N.J. Isolation and Identification of Actinomycetes from Fire Mountain, China, and the Elicitation of Antimicrobial Production. Ph.D. Thesis, Newcastle University, Newcastle, UK, 2020. [Google Scholar]
- Grkovic, T.; Abdelmohsen, U.S.; Othman, E.M.; Stopper, H.; Edrada-Ebel, R.; Hentschel, U.; Quinn, R.J. Two New Antioxidant Actinosporin Analogues from the Calcium Alginate Beads Culture of Sponge-Associated Actinokineospora Sp. Strain EG49. Bioorganic Med. Chem. Lett. 2014, 24, 5089–5092. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Othman, E.M.; Reimer, A.; Grüne, M.; Kozjak-Pavlovic, V.; Stopper, H.; Hentschel, U.; Abdelmohsen, U.R. Ageloline A, New Antioxidant and Antichlamydial Quinolone from the Marine Sponge-Derived Bacterium Streptomyces Sp. SBT345. Tetrahedron Lett. 2016, 57, 2786–2789. [Google Scholar] [CrossRef]
- Majithiya, V.R.; Gohel, S.D.; Majithiya, V.R.; Gohel, S.D. Actinobacteria Associated with Marine Invertebrates: Diversity and Biological Significance. In Actinobacteria—Diversity, Applications and Medical Aspects; IntechOpen: London, UK, 2022; ISBN 978-1-80355-097-8. [Google Scholar]
- Sun, Y.; Pham, A.N.; Hider, R.C.; Zheng, H.; Waite, T.D. Effectiveness of the Iron Chelator CN128 in Mitigating the Formation of Dopamine Oxidation Products Associated with the Progression of Parkinson’s Disease. ACS Chem. Neurosci. 2020, 11, 3646–3657. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, H.; Yao, G.; Qiao, P.; Li, L.; Wu, S. Therapeutic Potential of Iron Chelators on Osteoporosis and Their Cellular Mechanisms. Biomed. Pharmacother. 2021, 137, 111380. [Google Scholar] [CrossRef]
- Lan, J.; Jiang, D.H. Desferrioxamine and Vitamin E Protect against Iron and MPTP-Induced Neurodegeneration in Mice. J. Neural Transm. 1997, 104, 469–481. [Google Scholar] [CrossRef]
- Durland, J.; Ahmadian-Moghadam, H. Genetics, Mutagenesis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Hinarejos, I.; Machuca, C.; Sancho, P.; Espinós, C. Mitochondrial Dysfunction, Oxidative Stress and Neuroinflammation in Neurodegeneration with Brain Iron Accumulation (NBIA). Antioxidants 2020, 9, 1020. [Google Scholar] [CrossRef]
- Baumberg, S.; Krügel, H.; Noack, D. (Eds.) Genetics and Product Formation in Streptomyces; Springer: Boston, MA, USA, 1991; ISBN 978-1-4684-5924-1. [Google Scholar]
- Alvarez, H.M. (Ed.) Biology of Rhodococcus; Microbiology Monographs; Springer International Publishing: Cham, Switzerland, 2019; Volume 1, ISBN 978-3-030-11460-2. [Google Scholar]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural Products in Drug Discovery: Advances and Opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
 ». The strains isolated from cold sources are marked with a purple square «
». The strains isolated from cold sources are marked with a purple square « ». The strains isolated from hot sources are marked with a pink circle «
». The strains isolated from hot sources are marked with a pink circle « ».
». The strains isolated from cold sources are marked with a purple square «». The strains isolated from hot sources are marked with a pink circle «».
».
». The strains isolated from cold sources are marked with a purple square «». The strains isolated from hot sources are marked with a pink circle «». ». The strains isolated from cold sources are marked with a purple square «». The strains isolated from hot sources are marked with a pink circle «». The strains isolated from Lake Baikal are marked with a blue star «
». The strains isolated from cold sources are marked with a purple square «». The strains isolated from hot sources are marked with a pink circle «». The strains isolated from Lake Baikal are marked with a blue star « ».
». The strains isolated from cold sources are marked with a purple square «». The strains isolated from hot sources are marked with a pink circle «». The strains isolated from Lake Baikal are marked with a blue star «».
».
». The strains isolated from cold sources are marked with a purple square «». The strains isolated from hot sources are marked with a pink circle «». The strains isolated from Lake Baikal are marked with a blue star «».







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultural Liquid | Biomass | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Streptomyces sp. LPB2020M1 | Week No | Week No | ||||||||||
| 1 | 3 | 5 | 7 | 9 | 11 | 1 | 3 | 5 | 7 | 9 | 11 | |
| O2 | + | + | + | + | + | + | + | + | + | + | + | + |
| O3 | + | + | − | + | + | + | + | + | − | + | + | + |
| C | − | + | − | − | − | − | − | + | − | − | − | − |
| Cultural Liquid | Biomass | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Streptomyces sp. LPB2020M1 | Week No | Week No | ||||||||||
| 1 | 3 | 5 | 7 | 9 | 11 | 1 | 3 | 5 | 7 | 9 | 11 | |
| O2 | − | − | + | + | + | + | − | − | − | − | − | − |
| O3 | − | − | + | + | + | + | − | − | − | − | − | + |
| C | + | − | − | − | − | − | − | − | − | − | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dmitrieva, M.E.; Malygina, E.V.; Belyshenko, A.Y.; Shelkovnikova, V.N.; Imidoeva, N.A.; Morgunova, M.M.; Telnova, T.Y.; Vlasova, A.A.; Axenov-Gribanov, D.V. The Effects of a High Concentration of Dissolved Oxygen on Actinobacteria from Lake Baikal. Metabolites 2023, 13, 830. https://doi.org/10.3390/metabo13070830
Dmitrieva ME, Malygina EV, Belyshenko AY, Shelkovnikova VN, Imidoeva NA, Morgunova MM, Telnova TY, Vlasova AA, Axenov-Gribanov DV. The Effects of a High Concentration of Dissolved Oxygen on Actinobacteria from Lake Baikal. Metabolites. 2023; 13(7):830. https://doi.org/10.3390/metabo13070830
Chicago/Turabian StyleDmitrieva, Maria E., Ekaterina V. Malygina, Alexander Y. Belyshenko, Victoria N. Shelkovnikova, Natalia A. Imidoeva, Maria M. Morgunova, Tamara Y. Telnova, Anfisa A. Vlasova, and Denis V. Axenov-Gribanov. 2023. "The Effects of a High Concentration of Dissolved Oxygen on Actinobacteria from Lake Baikal" Metabolites 13, no. 7: 830. https://doi.org/10.3390/metabo13070830
APA StyleDmitrieva, M. E., Malygina, E. V., Belyshenko, A. Y., Shelkovnikova, V. N., Imidoeva, N. A., Morgunova, M. M., Telnova, T. Y., Vlasova, A. A., & Axenov-Gribanov, D. V. (2023). The Effects of a High Concentration of Dissolved Oxygen on Actinobacteria from Lake Baikal. Metabolites, 13(7), 830. https://doi.org/10.3390/metabo13070830