



Effects of RIPC on the Metabolomical Profile during Lower Limb Digital Subtraction Angiography: A Randomized Controlled Trial

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Trial Design
2.2. Participants
2.3. Randomization
2.4. Interventions
2.5. Blinding
2.6. Outcomes and Data Cleaning
2.7. Statistical Analysis
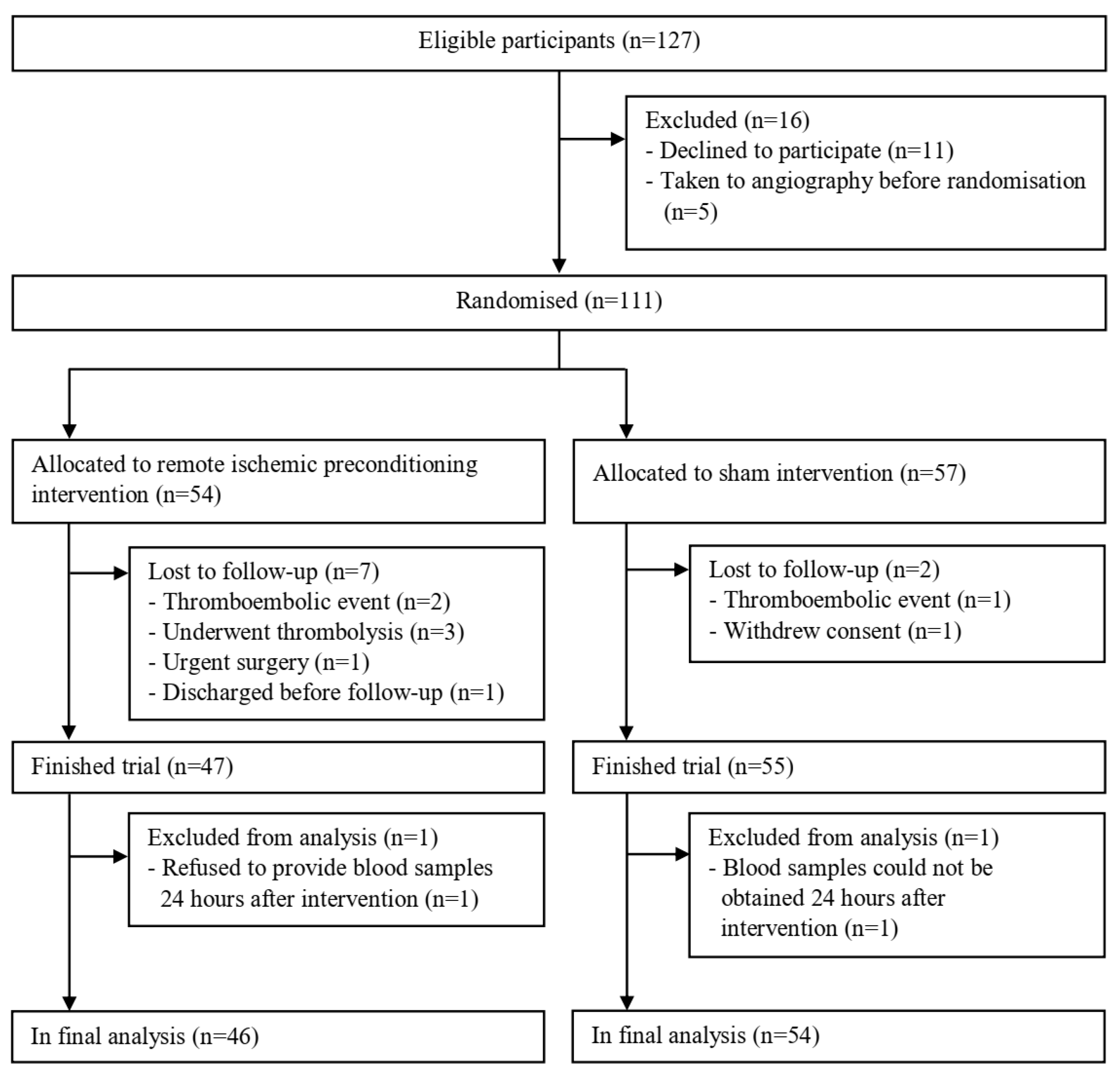
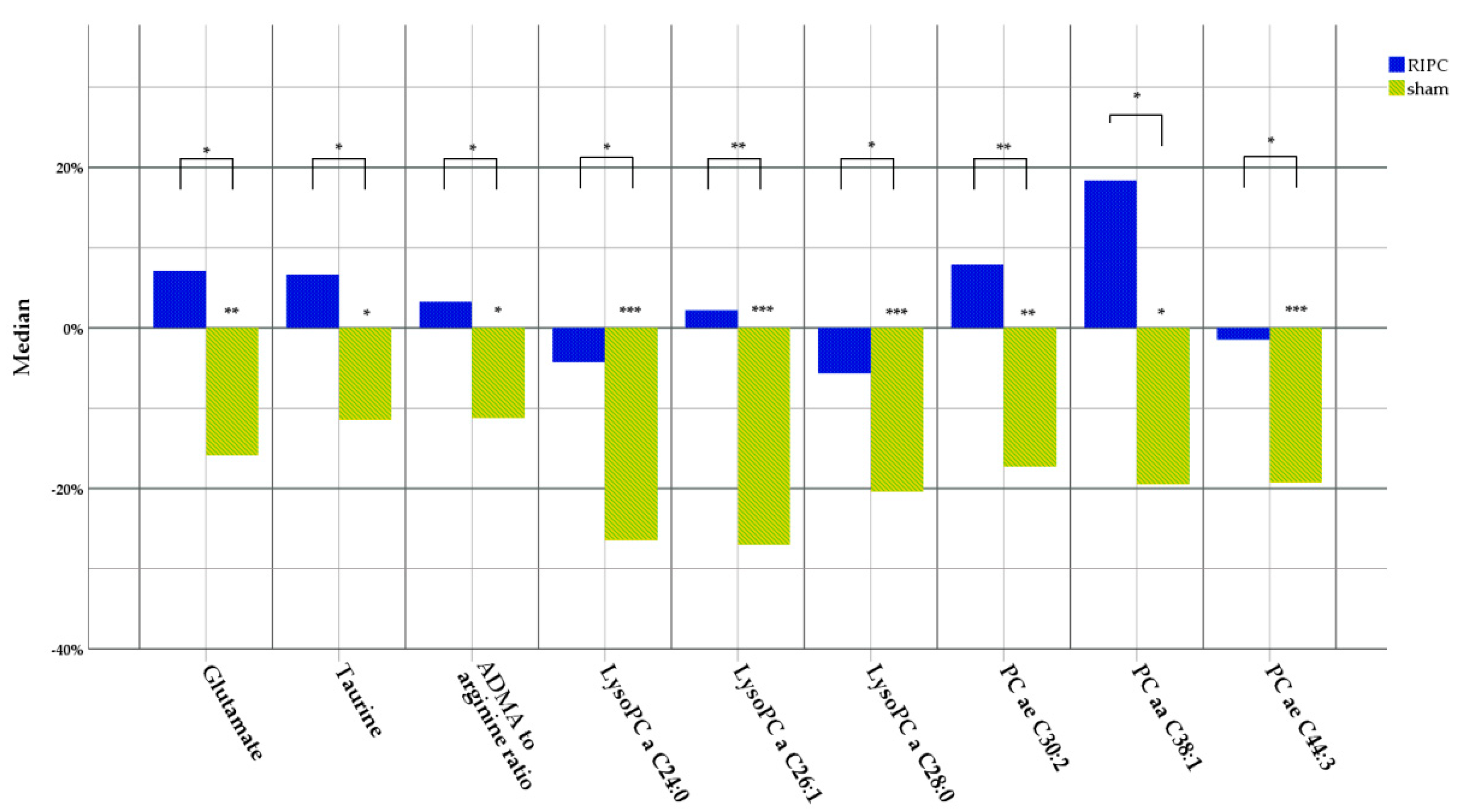
3. Results
4. Discussion
4.1. Taurine
4.2. Asymmetric Dimethyl Arginine and Arginine
4.3. Glutamate
4.4. Lysophosphatidylcholines and Phosphatidylcholines
4.5. Adiponectin and Its Effect on the Metabolomical Profile
4.6. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murry, C.E.; Jennings, R.B.; Reimer, K.A. Preconditioning with ischemia: A delay of lethal cell injury in ischemic myocardium. Circulation 1986, 74, 1124–1136. [Google Scholar] [CrossRef] [Green Version]
- Comità, S.; Rubeo, C.; Giordano, M.; Penna, C.; Pagliaro, P. Pathways for Cardioprotection in Perspective: Focus on Remote Conditioning and Extracellular Vesicles. Biology 2023, 12, 308. [Google Scholar] [CrossRef] [PubMed]
- Heusch, G. Molecular Basis of Cardioprotection. Circ. Res. 2015, 116, 674–699. [Google Scholar] [CrossRef]
- Yang, J.; Shakil, F.; Cho, S. Peripheral Mechanisms of Remote Ischemic Conditioning. Cond. Med. 2019, 2, 61–68. [Google Scholar]
- Kasepalu, T.; Kuusik, K.; Lepner, U.; Starkopf, J.; Zilmer, M.; Eha, J.; Vähi, M.; Kals, J. Remote Ischaemic Preconditioning Reduces Kidney Injury Biomarkers in Patients Undergoing Open Surgical Lower Limb Revascularisation: A Randomised Trial. Oxid. Med. Cell. Longev. 2020, 2020, 7098505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kepler, T.; Kuusik, K.; Lepner, U.; Starkopf, J.; Zilmer, M.; Eha, J.; Vähi, M.; Kals, J. Remote Ischaemic Preconditioning Attenuates Cardiac Biomarkers during Vascular Surgery: A Randomised Clinical Trial. Eur. J. Vasc. Endovasc. Surg. 2020, 59, 301–308. [Google Scholar] [CrossRef]
- Liang, F.; Liu, S.; Liu, G.; Liu, H.; Wang, Q.; Song, S.; Yao, L. Remote ischaemic preconditioning versus no remote ischaemic preconditioning for vascular and endovascular surgical procedures. Cochrane Database Syst. Rev. 2023, 1, CD008472. [Google Scholar] [CrossRef] [PubMed]
- Menting, T.P.; Wever, K.E.; Ozdemir-van Brunschot, D.M.; Van der Vliet, D.J.; Rovers, M.M.; Warle, M.C. Ischaemic preconditioning for the reduction of renal ischaemia reperfusion injury. Cochrane Database Syst. Rev. 2017, 3, CD010777. [Google Scholar] [CrossRef]
- Paapstel, K.; Kals, J. Metabolomics of Arterial Stiffness. Metabolites 2022, 12, 370. [Google Scholar] [CrossRef]
- Ismaeel, A.; Lavado, R.; Smith, R.S.; Eidson, J.L.; Sawicki, I.; Kirk, J.S.; Bohannon, W.T.; Koutakis, P. Effects of Limb Revascularization Procedures on Oxidative Stress. J. Surg. Res. 2018, 232, 503–509. [Google Scholar] [CrossRef]
- Pearce, L.; Davidson, S.M.; Yellon, D.M. Does remote ischaemic conditioning reduce inflammation? A focus on innate immunity and cytokine response. Basic Res. Cardiol. 2021, 116, 12. [Google Scholar] [CrossRef] [PubMed]
- Kasepalu, T.; Kuusik, K.; Lepner, U.; Starkopf, J.; Zilmer, M.; Eha, J.; Vähi, M.; Kals, J. Remote ischaemic preconditioning influences the levels of acylcarnitines in vascular surgery: A randomised clinical trial. Nutr. Metab. 2020, 17, 76. [Google Scholar] [CrossRef] [PubMed]
- Kuusik, K.; Kepler, T.; Zilmer, M.; Eha, J.; Vähi, M.; Kals, J. Effects of Remote Ischaemic Preconditioning on Arterial Stiffness in Patients Undergoing Lower Limb Angiographic Procedures: A Randomised Clinical Trial. Eur. J. Vasc. Endovasc. Surg. 2019, 58, 875–882. [Google Scholar] [CrossRef]
- Siskos, A.P.; Jain, P.; Römisch-Margl, W.; Bennett, M.; Achaintre, D.; Asad, Y.; Marney, L.; Richardson, L.; Koulman, A.; Griffin, J.L.; et al. Interlaboratory Reproducibility of a Targeted Metabolomics Platform for Analysis of Human Serum and Plasma. Anal. Chem. 2017, 89, 656–665. [Google Scholar] [CrossRef]
- Yu, Z.; Kastenmüller, G.; He, Y.; Belcredi, P.; Möller, G.; Prehn, C.; Mendes, J.; Wahl, S.; Roemisch-Margl, W.; Ceglarek, U.; et al. Differences between human plasma and serum metabolite profiles. PLoS ONE 2011, 6, e21230. [Google Scholar] [CrossRef]
- Breier, M.; Wahl, S.; Prehn, C.; Fugmann, M.; Ferrari, U.; Weise, M.; Banning, F.; Seissler, J.; Grallert, H.; Adamski, J.; et al. Targeted metabolomics identifies reliable and stable metabolites in human serum and plasma samples. PLoS ONE 2014, 9, e89728. [Google Scholar] [CrossRef] [PubMed]
- Liebisch, G.; Fahy, E.; Aoki, J.; Dennis, E.A.; Durand, T.; Ejsing, C.S.; Fedorova, M.; Feussner, I.; Griffiths, W.J.; Köfeler, H.; et al. Update on LIPID MAPS classification, nomenclature, and shorthand notation for MS-derived lipid structures. J. Lipid Res. 2020, 61, 1539–1555. [Google Scholar] [CrossRef]
- Stekhoven, D.J.; Bühlmann, P. MissForest—Non-parametric missing value imputation for mixed-type data. Bioinformatics 2012, 28, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Duszka, K. Versatile Triad Alliance: Bile Acid, Taurine and Microbiota. Cells 2022, 11, 2337. [Google Scholar] [CrossRef]
- Baliou, S.; Adamaki, M.; Ioannou, P.; Pappa, A.; Panayiotidis, M.I.; Spandidos, D.A.; Christodoulou, I.; Kyriakopoulos, A.M.; Zoumpourlis, V. Protective role of taurine against oxidative stress. Mol. Med. Rep. 2021, 24, 605. [Google Scholar] [CrossRef]
- Chao de la Barca, J.M.; Bakhta, O.; Kalakech, H.; Simard, G.; Tamareille, S.; Catros, V.; Callebert, J.; Gadras, C.; Tessier, L.; Reynier, P.; et al. Metabolic Signature of Remote Ischemic Preconditioning Involving a Cocktail of Amino Acids and Biogenic Amines. J. Am. Heart Assoc. 2016, 5, e003891. [Google Scholar] [CrossRef]
- Maric, S.; Restin, T.; Muff, J.L.; Camargo, S.M.; Guglielmetti, L.C.; Holland-Cunz, S.G.; Crenn, P.; Vuille-dit-Bille, R.N. Citrulline, Biomarker of Enterocyte Functional Mass and Dietary Supplement. Metabolism, Transport, and Current Evidence for Clinical Use. Nutrients 2021, 13, 2794. [Google Scholar] [CrossRef]
- Ismaeel, A.; Franco, M.E.; Lavado, R.; Papoutsi, E.; Casale, G.P.; Fuglestad, M.; Mietus, C.J.; Haynatzki, G.R.; Smith, R.S.; Bohannon, W.T.; et al. Altered Metabolomic Profile in Patients with Peripheral Artery Disease. J. Clin. Med. 2019, 8, 1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Chen, W.; Rezvan, A.; Jo, H.; Harrison, D.G. Tetrahydrobiopterin deficiency and nitric oxide synthase uncoupling contribute to atherosclerosis induced by disturbed flow. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1547–1554. [Google Scholar] [CrossRef] [Green Version]
- Westman, B.; Weidenhielm, L.; Rooyackers, O.; Fredriksson, K.; Wernerman, J.; Hammarqvist, F. Knee replacement surgery as a human clinical model of the effects of ischaemia/reperfusion upon skeletal muscle. Clin. Sci. 2007, 113, 313–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eerik, K.; Kasepalu, T.; Kuusik, K.; Eha, J.; Vähi, M.; Kilk, K.; Zilmer, M.; Kals, J. Effects of RIPC on the Metabolome in Patients Undergoing Vascular Surgery: A Randomized Controlled Trial. Biomolecules 2022, 12, 1312. [Google Scholar] [CrossRef]
- Shen, S.; Wang, J.F.; Wu, J.Q.; Zhou, J.X.; Meng, S.D.; Ma, J.; Zhu, C.; Deng, G.G.; Liu, D. GC/MS-based metabolomic analysis of alleviated renal ischemia-reperfusion injury induced by remote ischemic preconditioning. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 765–774. [Google Scholar] [PubMed]
- Polonis, K.; Wawrzyniak, R.; Daghir-Wojtkowiak, E.; Szyndler, A.; Chrostowska, M.; Melander, O.; Hoffmann, M.; Kordalewska, M.; Raczak-Gutknecht, J.; Bartosińska, E.; et al. Metabolomic Signature of Early Vascular Aging (EVA) in Hypertension. Front. Mol. Biosci. 2020, 7, 12. [Google Scholar] [CrossRef]
- Kennelly, J.P.; van der Veen, J.N.; Nelson, R.C.; Leonard, K.A.; Havinga, R.; Buteau, J.; Kuipers, F.; Jacobs, R.L. Intestinal de novo phosphatidylcholine synthesis is required for dietary lipid absorption and metabolic homeostasis. J. Lipid Res. 2018, 59, 1695–1708. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.P.; Riederer, M.; Lechleitner, M.; Hermansson, M.; Desoye, G.; Hallström, S.; Graier, W.F.; Frank, S. Acyl Chain-Dependent Effect of Lysophosphatidylcholine on Endothelium-Dependent Vasorelaxation. PLoS ONE 2013, 8, e65155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.Y.; Kim, O.Y.; Paik, J.K.; Kwon, D.Y.; Kim, H.J.; Lee, J.H. Association of age-related changes in circulating intermediary lipid metabolites, inflammatory and oxidative stress markers, and arterial stiffness in middle-aged men. Age 2013, 35, 1507–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, S.H.; Chan, M.L.; Marathe, G.K.; Parveen, F.; Chen, C.H.; Ke, L.Y. An Updated Review of Lysophosphatidylcholine Metabolism in Human Diseases. Int. J. Mol. Sci. 2019, 20, 1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stegemann, C.; Pechlaner, R.; Willeit, P.; Langley, S.R.; Mangino, M.; Mayr, U.; Menni, C.; Moayyeri, A.; Santer, P.; Rungger, G.; et al. Lipidomics profiling and risk of cardiovascular disease in the prospective population-based Bruneck study. Circulation 2014, 129, 1821–1831. [Google Scholar] [CrossRef] [Green Version]
- Kuusik, K.; Kasepalu, T.; Zilmer, M.; Eha, J.; Vähi, M.; Torop, L.A.; Lieberg, J.; Kals, J. The Role of RIPC in Preventing Organ Damage, Inflammation, and Oxidative Stress during Lower Limb DSA: A Randomised Controlled Trial. Oxid. Med. Cell. Longev. 2021, 2021, 6043550. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Liu, M. Adiponectin: Friend or foe in obesity and inflammation. Med. Rev. 2022, 2, 349–362. [Google Scholar] [CrossRef]
- Ahlstrom, P.; Rai, E.; Chakma, S.; Cho, H.H.; Rengasamy, P.; Sweeney, G. Adiponectin improves insulin sensitivity via activation of autophagic flux. J. Mol. Endocrinol. 2017, 59, 339–350. [Google Scholar] [CrossRef] [Green Version]
- Jeong, I.H.; Bae, W.Y.; Choi, J.S.; Jeong, J.W. Ischemia induces autophagy of endothelial cells and stimulates angiogenic effects in a hindlimb ischemia mouse model. Cell Death Dis. 2020, 11, 624. [Google Scholar] [CrossRef]
- Scalabrin, M.; Engman, V.; Maccannell, A.; Critchlow, A.; Roberts, L.D.; Yuldasheva, N.; Bowen, T.S. Temporal analysis of skeletal muscle remodeling post hindlimb ischemia reveals intricate autophagy regulation. Am. J. Physiol. Cell Physiol. 2022, 323, C1601–C1610. [Google Scholar] [CrossRef]
- Geng, J.; Zhang, Y.; Li, S.; Li, S.; Wang, J.; Wang, H.; Aa, J.; Wang, G. Metabolomic Profiling Reveals That Reprogramming of Cerebral Glucose Metabolism Is Involved in Ischemic Preconditioning-Induced Neuroprotection in a Rodent Model of Ischemic Stroke. J. Proteome Res. 2019, 18, 57–68. [Google Scholar] [CrossRef]
- Stenzel-Poore, M.P.; Stevens, S.L.; Xiong, Z.; Lessov, N.S.; Harrington, C.A.; Mori, M.; Meller, R.; Rosenzweig, H.L.; Tobar, E.; Shaw, T.E.; et al. Effect of ischaemic preconditioning on genomic response to cerebral ischaemia: Similarity to neuroprotective strategies in hibernation and hypoxia-tolerant states. Lancet 2003, 362, 1028–1037. [Google Scholar] [CrossRef]
- Gonzalez, N.R.; Hamilton, R.; Bilgin-Freiert, A.; Dusick, J.; Vespa, P.; Hu, X.; Asgari, S. Cerebral hemodynamic and metabolic effects of remote ischemic preconditioning in patients with subarachnoid hemorrhage. Acta Neurochir. Suppl. 2013, 115, 193–198. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Inclusion criteria |
|
| Exclusion criteria |
|
| Characteristics | RIPC (n = 46) | SHAM (n = 54) | p-Value | ||
|---|---|---|---|---|---|
| Mean/ Median | SD/IQR | Mean/ Median | SD/IQR | ||
| Demographic | |||||
| Male (n) | 33 (71.7%) | 47 (87.0%) | 0.098 | ||
| Mean age (y) | 66.1 | ±10.3 | 65.0 | ±11.4 | 0.61 |
| Weight (kg) | 75.2 | ±17.3 | 78.0 | ±16.7 | 0.42 |
| Body mass index (kg/m2) | 25.4 | (22.7–30.0) | 25.3 | (23.5–29.4) | 0.66 |
| Renal function at inclusion | |||||
| eGFR <90 (n) # | 27 (58.7%) | 30 (55.6%) | 0.91 | ||
| 60–89 (n) # | 19 (41.3%) | 20 (37.0%) | |||
| 30–59 (n) # | 8 (18.5%) | 10 (18.5%) | |||
| History of smoking (n) † | 35 (76.1%) | 41 (75.9%) | 1 | ||
| Concomitant diseases | |||||
| Stage of LEAD III or more ‡ | 23 (50.0%) | 25 (46.3%) | 0.87 | ||
| Stage of LEAD III (n) ‡ | 9 (19.6%) | 10 (18.5%) | |||
| Stage of LEAD IV (n) ‡ | 14 (30.4%) | 15 (27.8%) | |||
| Diabetes (n) | 10 (21.7%) | 13 (24.1%) | 0.97 | ||
| Hypertension (n) ◊ | 32 (69.6%) | 28 (51.9%) | 0.11 | ||
| Medications | |||||
| ACE inhibitors (n) | 18 (39.1%) | 14 (25.9%) | 0.16 | ||
| ARBs (n) | 12 (26.1%) | 11 (20.4%) | 0.50 | ||
| Calcium channel blockers (n) | 18 (39.1%) | 15 (29.8%) | 0.23 | ||
| Beta blockers (n) | 12 (26.1%) | 12 (22.2%) | 0.65 | ||
| Diuretics (n) | 16 (34.8%) | 12 (22.2%) | 0.16 | ||
| Antiagregants (n) | 24 (52.2%) | 26 (48.1%) | 0.69 | ||
| Anticoagulants (n) | 1 (2.2%) | 1 (1.9%) | 0.91 | ||
| Naftidrofuryl/pentoxifylline (n) | 33 (71.7%) | 35 (64.8%) | 0.46 | ||
| Statins (n) | 18 (39.1%) | 16 (29.6%) | 0.39 | ||
| Insulin therapy (n) | 6 (13.0%) | 8 (14.8%) | 0.80 | ||
| Oral antidiabetic agents (n) | 3 (6.5%) | 5 (9.3%) | 0.62 | ||
| Creatinine (μmol/L) | 78 | (65–92) | 77 | (67–92) | 0.80 |
| eGFR (mL/min/1.73 m2) | 84 | (68–94) | 91 | (69–100) | 0.17 |
| Urea (mmol/L) | 5.0 | (4.4–6.6) | 5.5 | (4.4–6.6) | 0.76 |
| Cholesterol (mmol/L) | 4.66 | ±1.38 | 4.85 | ±1.42 | 0.52 |
| HDL (mmol/L) | 1.17 | (0.96–1.55) | 1.12 | (0.94–1.45) | 0.58 |
| LDL (mmol/L) | 2.70 | (2.07–3.63) | 3.02 | (2.05–3.91) | 0.50 |
| TG (mmol/L) | 1.30 | (0.98–2.06) | 1.43 | (1.1–1.98) | 0.36 |
| Adiponectin (ng/mL) | 6322 | (3769–8523) | 5541 | (3327–9406) | 0.48 |
| Group | Change | IQR/SD | p-Value | |
|---|---|---|---|---|
| Asymmetric dimethyl arginine | SHAM | −0.075 | (−0.367–0.217) | 0.005 |
| Citrulline-to-arginine ratio | SHAM | −0.042 | (−0.157–0.074) | 0.003 |
| Citrulline | SHAM | −5.3 | (−25.2–14.6) | 0.003 |
| Glutamate | SHAM | −18.0 | (−66.3–30.3) | 0.004 |
| Total dimethylamide | SHAM | −0.173 | (−0.876–0.530) | 0.001 |
| Tyrosine | RIPC | −9.8 | (−32.5–12.9) | 0.004 |
| Tyrosine-to-phenylalanine ratio♦ | SHAM | 0.08 | 0.20 | 0.004 |
| Tyrosine-to-phenylalanine ratio♦ | RIPC | 0.12 | 0.19 | <0.001 |
| LysoPC a C16:0 | SHAM | −25.0 | (−89.6–39.7) | 0.003 |
| LysoPC a C17:0 | SHAM | −0.59 | (−1.91–0.73) | <0.001 |
| LysoPC a C18:0 | SHAM | −4.9 | (−15.3–5.6) | <0.001 |
| LysoPC a C18:0 | RIPC | −6.2 | (−20.2–7.8) | 0.002 |
| LysoPC a C18:1 | SHAM | −8.8 | (−23.3–5.7) | <0.001 |
| LysoPC a C18:1 | RIPC | −7.07 | (−26.1–11.9) | 0.001 |
| LysoPC a C18:2 | SHAM | −16.1 | (−48.4–16.2) | <0.001 |
| LysoPC a C18:2 | RIPC | −13.0 | (−50.4–24.3) | <0.001 |
| LysoPC a C20:3 | SHAM | −0.94 | (−2.74–0.87) | <0.001 |
| LysoPC a C20:3 | RIPC | −1.0 | (−3.5–1.4) | 0.001 |
| LysoPC a C20:4 | SHAM | −2.5 | (−7.5–2.5) | <0.001 |
| LysoPC a C20:4 | RIPC | −3.3 | (−11.3–4.7) | 0.002 |
| LysoPC a C24:0 | SHAM | −0.21 | (−0.73–0.32) | 0.001 |
| LysoPC a C26:1 | SHAM | −0.25 | (−1.04–0.55) | 0.001 |
| LysoPC a C28:0 | SHAM | −0.23 | (−0.86–0.39) | 0.001 |
| PC ae C30:0 | SHAM | −0.026 | (−0.132–0.081) | 0.005 |
| PC ae C30:2 | SHAM | −0.027 | (−0.123–0.069) | 0.002 |
| PC ae C34:3 | RIPC | −0.75 | (−2.30–0.80) | 0.001 |
| PC ae C42:1 | SHAM | −0.094 | (−0.323–0.136) | 0.005 |
| PC ae C44:3 | SHAM | −0.041 | (−0.154–0.071) | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuusik, K.; Kasepalu, T.; Zilmer, M.; Eha, J.; Paapstel, K.; Kilk, K.; Rehema, A.; Kals, J. Effects of RIPC on the Metabolomical Profile during Lower Limb Digital Subtraction Angiography: A Randomized Controlled Trial. Metabolites 2023, 13, 856. https://doi.org/10.3390/metabo13070856
Kuusik K, Kasepalu T, Zilmer M, Eha J, Paapstel K, Kilk K, Rehema A, Kals J. Effects of RIPC on the Metabolomical Profile during Lower Limb Digital Subtraction Angiography: A Randomized Controlled Trial. Metabolites. 2023; 13(7):856. https://doi.org/10.3390/metabo13070856
Chicago/Turabian StyleKuusik, Karl, Teele Kasepalu, Mihkel Zilmer, Jaan Eha, Kaido Paapstel, Kalle Kilk, Aune Rehema, and Jaak Kals. 2023. "Effects of RIPC on the Metabolomical Profile during Lower Limb Digital Subtraction Angiography: A Randomized Controlled Trial" Metabolites 13, no. 7: 856. https://doi.org/10.3390/metabo13070856
APA StyleKuusik, K., Kasepalu, T., Zilmer, M., Eha, J., Paapstel, K., Kilk, K., Rehema, A., & Kals, J. (2023). Effects of RIPC on the Metabolomical Profile during Lower Limb Digital Subtraction Angiography: A Randomized Controlled Trial. Metabolites, 13(7), 856. https://doi.org/10.3390/metabo13070856