
Metabolic Bile Acid Profile Impairments in Dogs Affected by Chronic Inflammatory Enteropathy

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Faecal Sample Preparation
2.3. HPLC-ESI-MS/MS Conditions
2.4. Study Population
2.5. Statistical Analysis
3. Results
3.1. Pathological Subjects
3.2. Pathological vs. Control Subjects
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jiang, L.; Zhang, H.; Xiao, D.; Wei, H.; Chen, Y. Farnesoid X Receptor (FXR): Structures and Ligands. Comput. Struct. Biotechnol. J. 2021, 19, 2148–2159. [Google Scholar] [CrossRef]
- Balakrishnan, A.; Polli, J.E. Apical Sodium Dependent Bile Acid Transporter (ASBT, SLC10A2): A Potential Prodrug Target. Mol. Pharm. 2006, 3, 223–230. [Google Scholar] [CrossRef]
- Washizu, T.; Ishida, T.; Washizu, M.; Tomoda, I.; Kaneko, J.J. Changes in Bile Acid Composition of Serum and Gallbladder Bile in Bile Duct Ligated Dogs. J. Vet. Med. Sci. 1994, 56, 299–303. [Google Scholar] [CrossRef]
- Lin, Q.; Tan, X.; Wang, W.; Zeng, W.; Gui, L.; Su, M.; Liu, C.; Jia, W.; Xu, L.; Lan, K. Species Differences of Bile Acid Redox Metabolism: Tertiary Oxidation of Deoxycholate Is Conserved in Preclinical Animals. Drug Metab. Dispos. 2020, 48, 499–507. [Google Scholar] [CrossRef]
- Porru, E.; Scicchitano, D.; Interino, N.; Tavella, T.; Candela, M.; Roda, A.; Fiori, J. Analysis of Fecal Bile Acids and Metabolites by High Resolution Mass Spectrometry in Farm Animals and Correlation with Microbiota. Sci. Rep. 2022, 12, 2866. [Google Scholar] [CrossRef]
- Collins, S.L.; Stine, J.G.; Bisanz, J.E.; Okafor, C.D.; Patterson, A.D. Bile Acids and the Gut Microbiota: Metabolic Interactions and Impacts on Disease. Nat. Rev. Microbiol. 2023, 21, 236–247. [Google Scholar] [CrossRef]
- Jahnel, J.; Fickert, P.; Hauer, A.C.; Högenauer, C.; Avian, A.; Trauner, M. Inflammatory Bowel Disease Alters Intestinal Bile Acid Transporter Expression. Drug Metab. Dispos. 2014, 42, 1423–1431. [Google Scholar] [CrossRef]
- Cornet, N.; Battat, R. Diet, Deficiencies, and Inflammatory Bowel Diseases: A Comprehensive Review. Curr. Treat. Options Gastroenterol. 2023, 21, 157–171. [Google Scholar] [CrossRef]
- Dawson, P.A.; Haywood, J.; Craddock, A.L.; Wilson, M.; Tietjen, M.; Kluckman, K.; Maeda, N.; Parks, J.S. Targeted Deletion of the Ileal Bile Acid Transporter Eliminates Enterohepatic Cycling of Bile Acids in Mice. J. Biol. Chem. 2003, 278, 33920–33927. [Google Scholar] [CrossRef]
- Jung, D.; Inagaki, T.; Gerard, R.D.; Dawson, P.A.; Kliewer, S.A.; Mangelsdorf, D.J.; Moschetta, A. FXR Agonists and FGF15 Reduce Fecal Bile Acid Excretion in a Mouse Model of Bile Acid Malabsorption. J. Lipid Res. 2007, 48, 2693–2700. [Google Scholar] [CrossRef]
- Vivian, D.; Cheng, K.; Khurana, S.; Xu, S.; Kriel, E.H.; Dawson, P.A.; Raufman, J.-P.; Polli, J.E. In Vivo Performance of a Novel Fluorinated Magnetic Resonance Imaging Agent for Functional Analysis of Bile Acid Transport. Mol. Pharm. 2014, 11, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Blake, A.B.; Suchodolski, J.S. Importance of Gut Microbiota for the Health and Disease of Dogs and Cats. Anim. Front. 2016, 6, 37–42. [Google Scholar] [CrossRef]
- Honneffer, J.B.; Minamoto, Y.; Suchodolski, J.S. Microbiota Alterations in Acute and Chronic Gastrointestinal Inflammation of Cats and Dogs. World J. Gastroenterol. 2014, 20, 16489–16497. [Google Scholar] [CrossRef] [PubMed]
- Suchodolski, J.S.; Markel, M.E.; Garcia-Mazcorro, J.F.; Unterer, S.; Heilmann, R.M.; Dowd, S.E.; Kachroo, P.; Ivanov, I.; Minamoto, Y.; Dillman, E.M.; et al. The Fecal Microbiome in Dogs with Acute Diarrhea and Idiopathic Inflammatory Bowel Disease. PLoS ONE 2012, 7, e51907. [Google Scholar] [CrossRef] [PubMed]
- Xenoulis, P.G.; Palculict, B.; Allenspach, K.; Steiner, J.M.; Van House, A.M.; Suchodolski, J.S. Molecular-Phylogenetic Characterization of Microbial Communities Imbalances in the Small Intestine of Dogs with Inflammatory Bowel Disease. FEMS Microbiol. Ecol. 2008, 66, 579–589. [Google Scholar] [CrossRef]
- Isaiah, A.; Parambeth, J.C.; Steiner, J.M.; Lidbury, J.A.; Suchodolski, J.S. The Fecal Microbiome of Dogs with Exocrine Pancreatic Insufficiency. Anaerobe 2017, 45, 50–58. [Google Scholar] [CrossRef]
- Giaretta, P.R.; Rech, R.R.; Guard, B.C.; Blake, A.B.; Blick, A.K.; Steiner, J.M.; Lidbury, J.A.; Cook, A.K.; Hanifeh, M.; Spillmann, T.; et al. Comparison of Intestinal Expression of the Apical Sodium-Dependent Bile Acid Transporter between Dogs with and without Chronic Inflammatory Enteropathy. J. Vet. Intern. Med. 2018, 32, 1918–1926. [Google Scholar] [CrossRef]
- Guard, B.C.; Honneffer, J.B.; Jergens, A.E.; Jonika, M.M.; Toresson, L.; Lawrence, Y.A.; Webb, C.B.; Hill, S.; Lidbury, J.A.; Steiner, J.M.; et al. Longitudinal Assessment of Microbial Dysbiosis, Fecal Unconjugated Bile Acid Concentrations, and Disease Activity in Dogs with Steroid-responsive Chronic Inflammatory Enteropathy. J. Vet. Intern. Med. 2019, 33, 1295–1305. [Google Scholar] [CrossRef]
- Wang, S.; Martins, R.; Sullivan, M.C.; Friedman, E.S.; Misic, A.M.; El-Fahmawi, A.; De Martinis, E.C.P.; O’Brien, K.; Chen, Y.; Bradley, C.; et al. Diet-Induced Remission in Chronic Enteropathy Is Associated with Altered Microbial Community Structure and Synthesis of Secondary Bile Acids. Microbiome 2019, 7, 126. [Google Scholar] [CrossRef]
- Franco, P.; Porru, E.; Fiori, J.; Gioiello, A.; Cerra, B.; Roda, G.; Caliceti, C.; Simoni, P.; Roda, A. Identification and Quantification of Oxo-Bile Acids in Human Faeces with Liquid Chromatography–Mass Spectrometry: A Potent Tool for Human Gut Acidic Sterolbiome Studies. J. Chromatogr. A 2019, 1585, 70–81. [Google Scholar] [CrossRef]
- Porru, E.; Giorgi, E.; Turroni, S.; Helg, R.; Silani, M.; Candela, M.; Fiori, J.; Roda, A. Bile Acids and Oxo-Metabolites as Markers of Human Faecal Input in the Ancient Pompeii Ruins. Sci. Rep. 2021, 11, 3650. [Google Scholar] [CrossRef] [PubMed]
- Allenspach, K.; Wieland, B.; Gröne, A.; Gaschen, F. Chronic Enteropathies in Dogs: Evaluation of Risk Factors for Negative Outcome. J. Vet. Intern. Med. 2007, 21, 700–708. [Google Scholar] [CrossRef]
- Suchodolski, J.S. Diagnosis and Interpretation of Intestinal Dysbiosis in Dogs and Cats. Vet. J. 2016, 215, 30–37. [Google Scholar] [CrossRef]
- Duboc, H.; Rainteau, D.; Rajca, S.; Humbert, L.; Farabos, D.; Maubert, M.; Grondin, V.; Jouet, P.; Bouhassira, D.; Seksik, P.; et al. Increase in Fecal Primary Bile Acids and Dysbiosis in Patients with Diarrhea-Predominant Irritable Bowel Syndrome: Bile Acids and Dysbiosis in IBS-D Patients. Neurogastroenterol. Motil. 2012, 24, 513-e247. [Google Scholar] [CrossRef] [PubMed]
- Kriaa, A.; Mariaule, V.; Jablaoui, A.; Rhimi, S.; Mkaouar, H.; Hernandez, J.; Korkmaz, B.; Lesner, A.; Maguin, E.; Aghdassi, A.; et al. Bile Acids: Key Players in Inflammatory Bowel Diseases? Cells 2022, 11, 901. [Google Scholar] [CrossRef] [PubMed]
- Midtvedt, T.; Norman, A. Anaerobic, bile acid transforming microorganisms in rat intestinal content. Acta Pathol. Microbiol. Scand. 2009, 72, 337–344. [Google Scholar] [CrossRef]
- MacDonald, I.A.; Noel Williams, C.; Mahony, D.E. A Rapid Non-Chromatographic Analysis of Individual Bile Acids in Human Bile Extracts. J. Theor. Biol. 1976, 57, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Fiorucci, S.; Carino, A.; Baldoni, M.; Santucci, L.; Costanzi, E.; Graziosi, L.; Distrutti, E.; Biagioli, M. Bile Acid Signaling in Inflammatory Bowel Diseases. Dig. Dis. Sci. 2021, 66, 674–693. [Google Scholar] [CrossRef]
- Honneffer, J.; Guard, B.; Steiner, J.M.; Suchodolski, J.S. Mo1805 Untargeted Metabolomics Reveals Disruption Within Bile Acid, Cholesterol, and Tryptophan Metabolic Pathways in Dogs With Idiopathic Inflammatory Bowel Disease. Gastroenterology 2015, 148, S-715. [Google Scholar] [CrossRef]
- Xu, J.; Verbrugghe, A.; Lourenço, M.; Janssens, G.P.J.; Liu, D.J.X.; Van de Wiele, T.; Eeckhaut, V.; Van Immerseel, F.; Van de Maele, I.; Niu, Y.; et al. Does Canine Inflammatory Bowel Disease Influence Gut Microbial Profile and Host Metabolism? BMC Vet. Res. 2016, 12, 114. [Google Scholar] [CrossRef]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef] [PubMed]
- Hang, S.; Paik, D.; Yao, L.; Kim, E.; Trinath, J.; Lu, J.; Ha, S.; Nelson, B.N.; Kelly, S.P.; Wu, L.; et al. Bile Acid Metabolites Control TH17 and Treg Cell Differentiation. Nature 2019, 576, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Sun, X.; Oh, S.F.; Wu, M.; Zhang, Y.; Zheng, W.; Geva-Zatorsky, N.; Jupp, R.; Mathis, D.; Benoist, C.; et al. Microbial Bile Acid Metabolites Modulate Gut RORγ+ Regulatory T Cell Homeostasis. Nature 2020, 577, 410–415. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Oxo-BA | Common Name | Retention Time (min) | Quantifier/Qualifier (m/z) | Lipid Maps ID |
|---|---|---|---|---|
| 3,7,12-trioxo-5β-cholan-24-oic acid | trioxo-CA | 8.6 | [401.2]–[401.2] | LMST04010106 |
| 7α,12β-dihydroxy-3-oxo-5β-cholan-24-oic acid | 12β-3-oxo-CA | 12.5 | [405.3]–[405.3] | Not reported |
| 7β,12α-dihydroxy-3-oxo-5β-cholan-24-oic acid | 3-oxo-UCA | 12.9 | [405.3]–[405.3] | Not reported |
| 3α,6α-dihydroxy-7-oxo-5β-cholan-24-oic acid | 7-oxo-HCA | 17.86 | [405.3]–[405.3] | LMST04010173 |
| 3α,12α-dihydroxy-7-oxo-5β-cholan-24-oic acid | 7-oxo-CA | 19.49 | [405.3]–[405.3] | LMST04010184 |
| 3α,7α-dihydroxy-12-oxo-5β-cholan-24-oic acid | 12oxo-CA | 20.85 | [405.3]–[405.3] | LMST04010176 |
| 3,7-dioxo-5β-cholan-24-oic acid | 3,7-dioxo-CDCA | 22.48 | [387.3]–[387.3] | LMST04010136 |
| 6α,7α-dihydroxy-3-oxo-5β-cholan-24-oic acid | 3-oxo-HCA | 23.18 | [405.3]–[405.3] | LMST04010145 |
| 3,12-dioxo-5β-cholan-24-oic acid | 3,12-dioxo-DCA | 23.18 | [387.3]–[387.3] | LMST04010138 |
| 7β-hydroxy-3-oxo-5β-cholan-24-oic acid | 3-oxo-UDCA | 24.44 | [389.3]–[389.3] | LMST04010162 |
| 3,6-dioxo-5β-cholan-24-oic acid | 3,6-dioxo-HDCA | 24.10 | [387.3]–[387.3] | LMST04010134 |
| 7α,12α-dihydroxy-3-oxo-5β-cholan-24-oic acid | 3-oxo-CA | 25.8 | [405.3]–[405.3] | LMST04010443 |
| 6α-hydroxy-3-oxo-5β-cholan-24-oic acid | 3-oxo-HDCA | 25.92 | [389.3]–[389.3] | LMST04010158 |
| 3α-hydroxy-6-oxo-5β-cholan-24-oic acid | 6-oxo-HDCA | 26.32 | [389.3]–[389.3] | LMST04010146 |
| 3α-hydroxy-7-oxo-5β-cholan-24-oic acid | 7-oxo-CDCA | 26.92 | [389.3]–[389.3] | LMST04010150 |
| 12β-hydroxy-3-oxo-5β-cholan-24-oic acid | 12β-3-oxo-DCA | 27.77 | [389.3]–[389.3] | LMST04010157 |
| 3α-hydroxy-12-oxo-5β-cholan-24-oic acid | 12-oxo-DCA | 28.67 | [389.3]–[389.3] | LMST04010155 |
| 3α-hydroxy-6,7-dioxo-5β-cholan-24-oic acid | 6,7-dioxo-CA | 28.88 | [403.2]–[403.2] | Not reported |
| 7α-hydroxy-3-oxo-5β-cholan-24-oic acid | 3oxo-CDCA | 31.83 | [389.3]–[389.3] | LMST04010161 |
| 12α-hydroxy-3-oxo-5β-cholan-24-oic acid | 3oxo-DCA | 32.58 | [389.3]–[389.3] | LMST04010168 |
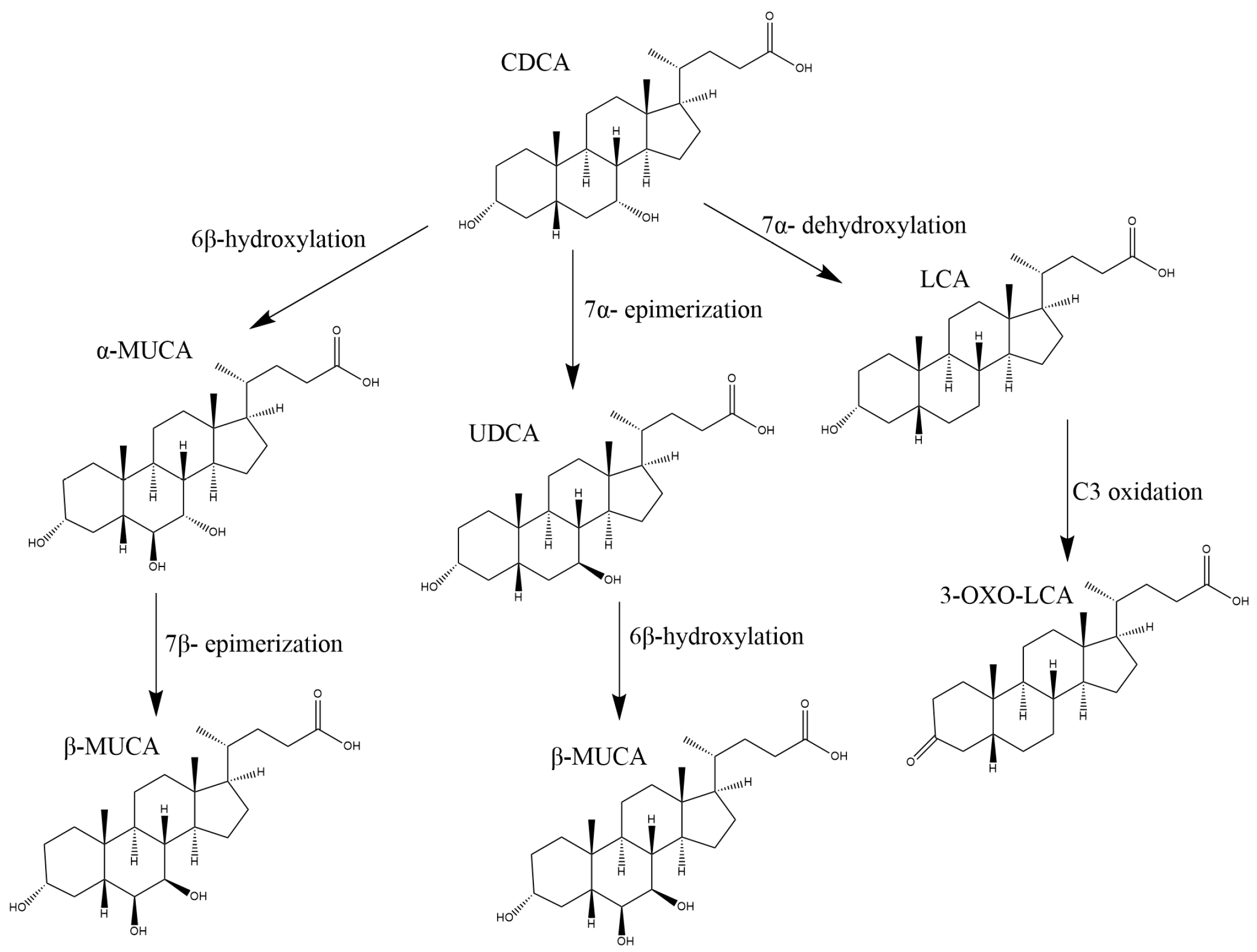
| 3-oxo-5β-cholan-24-oic acid | 3oxo-LCA | 38.22 | [373.2]–[373.2] | LMST04010127 |
| BA | ||||
| 3α,6α,7β-trihydroxy-5β-cholan-24-oic acid | (ωMUCA) | 20.27 | [407.2]–[407.2] | LMST04010065 |
| 3α,6β,7α-trihydroxy-5β-cholan-24-oic acid | (αMUCA) | 19.85 | [407.2]–[407.2] | LMST04010066 |
| 3α,6β,7β-trihydroxy-5β-cholan-24-oic acid | (βMUCA) | 20.87 | [407.2]–[407-2] | LMST04010067 |
| 3α,6α,7α-trihydroxy-5β-cholan-24-oic acid | HCA | 25.9 | [407.2]–[407.2] | LMST04010064 |
| 3α,7β-dihydroxy-5β-cholan-24-oic acid | UDCA | 26.28 | [391.2]–[391.2] | LMST04010033 |
| 3α,6α-dihydroxy-5β-cholan-24-oic acid | HDCA | 28.19 | [391.2]–[391.2] | LMST04010024 |
| 3α,7α,12α-trihydroxy-5β-cholan-24-oic acid | CA | 30.07 | [407.2]–[407.2] | LMST04010001 |
| 3α,7α-dihydroxy-5β-cholan-24-oic acid | CDCA | 35.15 | [391.2]–[391.2] | LMST04010032 |
| 3α,12α-dihydroxy-5β-cholan-24-oic acid | DCA | 35.91 | [391.2]–[391.2] | LMST04010040 |
| 3α-hydroxy-5β-cholan-24-oic acid | LCA | 39.56 | [375.2]–[375.2] | LMST04010003 |
| Pathological Subjects (n = 16) μg/g | Control Subjects (n = 16) μg/g | p-Value | |
|---|---|---|---|
| Primary BAs | |||
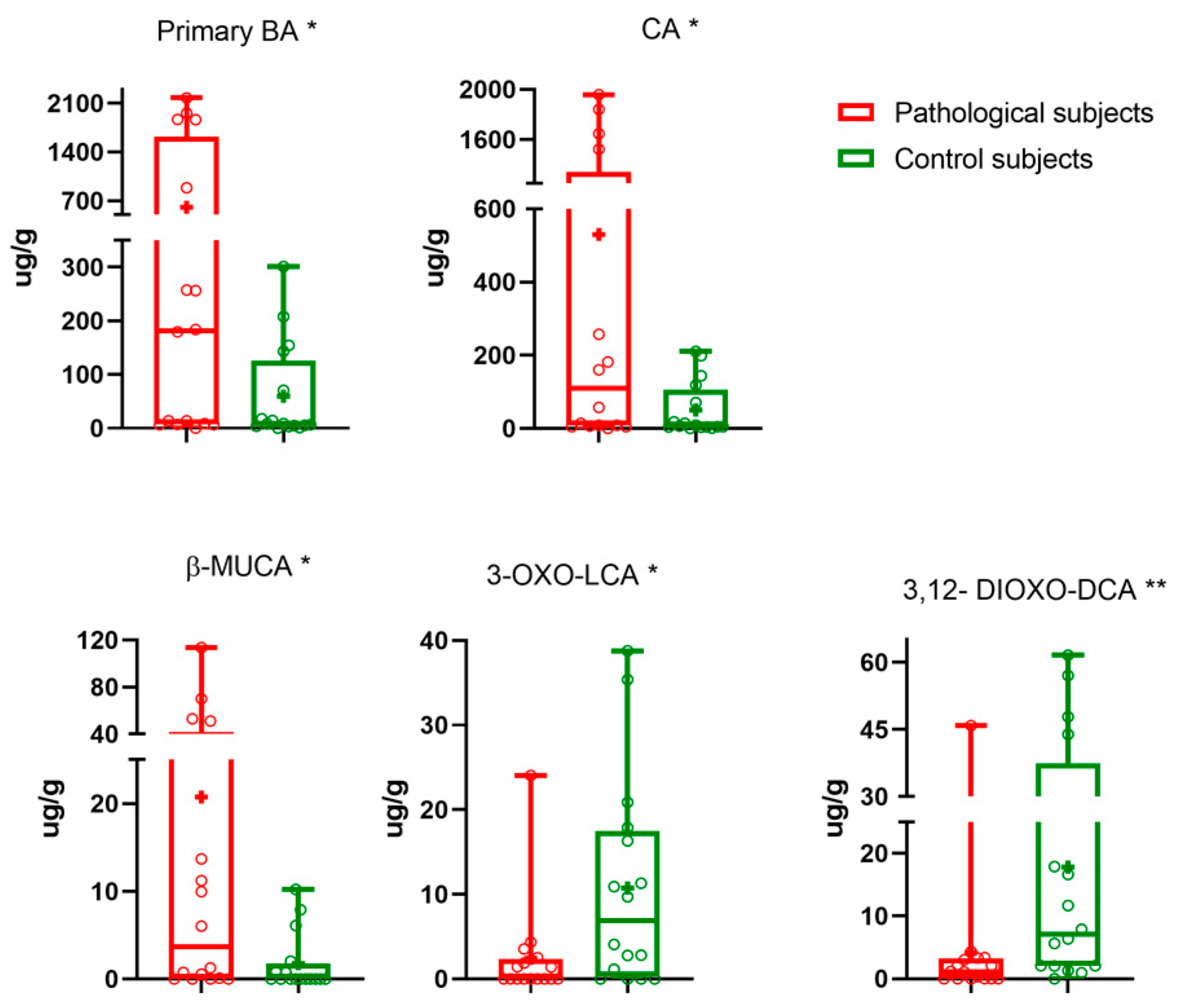
| CA + CDCA | 605 ± 211 | 8 [4–125] | 0.032 |
| CA | 530 ± 189 | 51 ± 19 | 0.017 |
| CDCA | 3.6 [0.01–111.8] | 0.01 [0.01–7.76] | 0.057 |
| Secondary BAs | |||
| DCA + LCA + UDCA + α-MUCA + β-MUCA | 594 [84–1106] | 1001 [190–1537] | 0.4230 |
| DCA | 496 [5–951] | 699 [7–1216] | 0.896 |
| LCA | 77.07 [0.09–203.2] | 171 ± 36 | 0.137 |
| UDCA | 0.63 [0.01–5.55] | 1.9 [0.01–4.49] | >0.999 |
| α-MUCA | 1.4 [0.8–3.5] | 0.30 [0.01–4.00] | 0.218 |
| β-MUCA | 3.65 [0.03–41.83] | 0.01 [0.01–1.74] | 0.021 |
| Oxo-BAs | 79 [35–411.4] | 216 ± 40 | 0.386 |
| 12-oxo-CA | 0.41 [0.01–4.06] | 0.56 [0.01–2.57] | 0.946 |
| 7-oxo-CA | 12 [1–128] | 27 ± 10 | 0.545 |
| 3-oxo-CDCA | 0.43 [0.01–2.11] | 0.01 [0.01–1.15] | 0.126 |
| 7-oxo-CDCA | 15 ± 5 | 0.01 [0.01–5.21] | 0.198 |
| 12-oxo-DCA | 16.02 [0.01–97.97] | 94 ± 25 | 0.233 |
| 3-oxo-DCA | 4.70 [0.01–40.25] | 48 ± 13 | 0.117 |
| 3,12-dioxo-DCA | 1.55 [0.01–3.24] | 7 [2–37] | 0.007 |
| 3-oxo-LCA | 0.01 [0.01–2.36] | 10± 3 | 0.017 |
| 3-oxo-UDCA | 0.01 [0.010–0.36] | 0.01 [0.01–0,01] | 0.434 |
| Non-oxo-BAs | 1027 [682–2221] | 879 [264–1426] | 0.361 |
| Total BAs | 1102 [733–3041] | 1076 [407–1723] | 0.539 |
| BA * | OR (95% CI) | p-Value | Cut-Off | Sensitivity-Specificity | AUC (95% CI) |
|---|---|---|---|---|---|
| β-MUCA | 3.67 (0.85–15.84) | 0.082 | −4.60 | 75–62.5% | 0.73 (0.56–0.90) |
| 3-oxo-LCA | 0.15 (0.03–0.71) | 0.017 | 1.03 | 81–69% | 0.74 (0.56–0.91) |
| 3,12-dioxo-DCA | 0.04 (0.004–0.384) | 0.005 | 1.73 | 93.75–62.50% | 0.78 (0.62–0.95) |
| CA | 3.86 (0.9–17) | 0.078 | 2.16 | 75–56.25% | 0.72 (0.54–0.90) |
| Primary BAs | 2.83 (0.67- 12) | 0.159 | 2.16 | 75–56.25% | 0.72 (0.54–0.90) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Comito, R.; Porru, E.; Interino, N.; Conti, M.; Terragni, R.; Gotti, R.; Candela, M.; Simoni, P.; Roda, A.; Fiori, J. Metabolic Bile Acid Profile Impairments in Dogs Affected by Chronic Inflammatory Enteropathy. Metabolites 2023, 13, 980. https://doi.org/10.3390/metabo13090980
Comito R, Porru E, Interino N, Conti M, Terragni R, Gotti R, Candela M, Simoni P, Roda A, Fiori J. Metabolic Bile Acid Profile Impairments in Dogs Affected by Chronic Inflammatory Enteropathy. Metabolites. 2023; 13(9):980. https://doi.org/10.3390/metabo13090980
Chicago/Turabian StyleComito, Rossana, Emanuele Porru, Nicolò Interino, Matteo Conti, Rossella Terragni, Roberto Gotti, Marco Candela, Patrizia Simoni, Aldo Roda, and Jessica Fiori. 2023. "Metabolic Bile Acid Profile Impairments in Dogs Affected by Chronic Inflammatory Enteropathy" Metabolites 13, no. 9: 980. https://doi.org/10.3390/metabo13090980
APA StyleComito, R., Porru, E., Interino, N., Conti, M., Terragni, R., Gotti, R., Candela, M., Simoni, P., Roda, A., & Fiori, J. (2023). Metabolic Bile Acid Profile Impairments in Dogs Affected by Chronic Inflammatory Enteropathy. Metabolites, 13(9), 980. https://doi.org/10.3390/metabo13090980