Polyamines under Abiotic Stress: Metabolic Crossroads and Hormonal Crosstalks in Plants

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
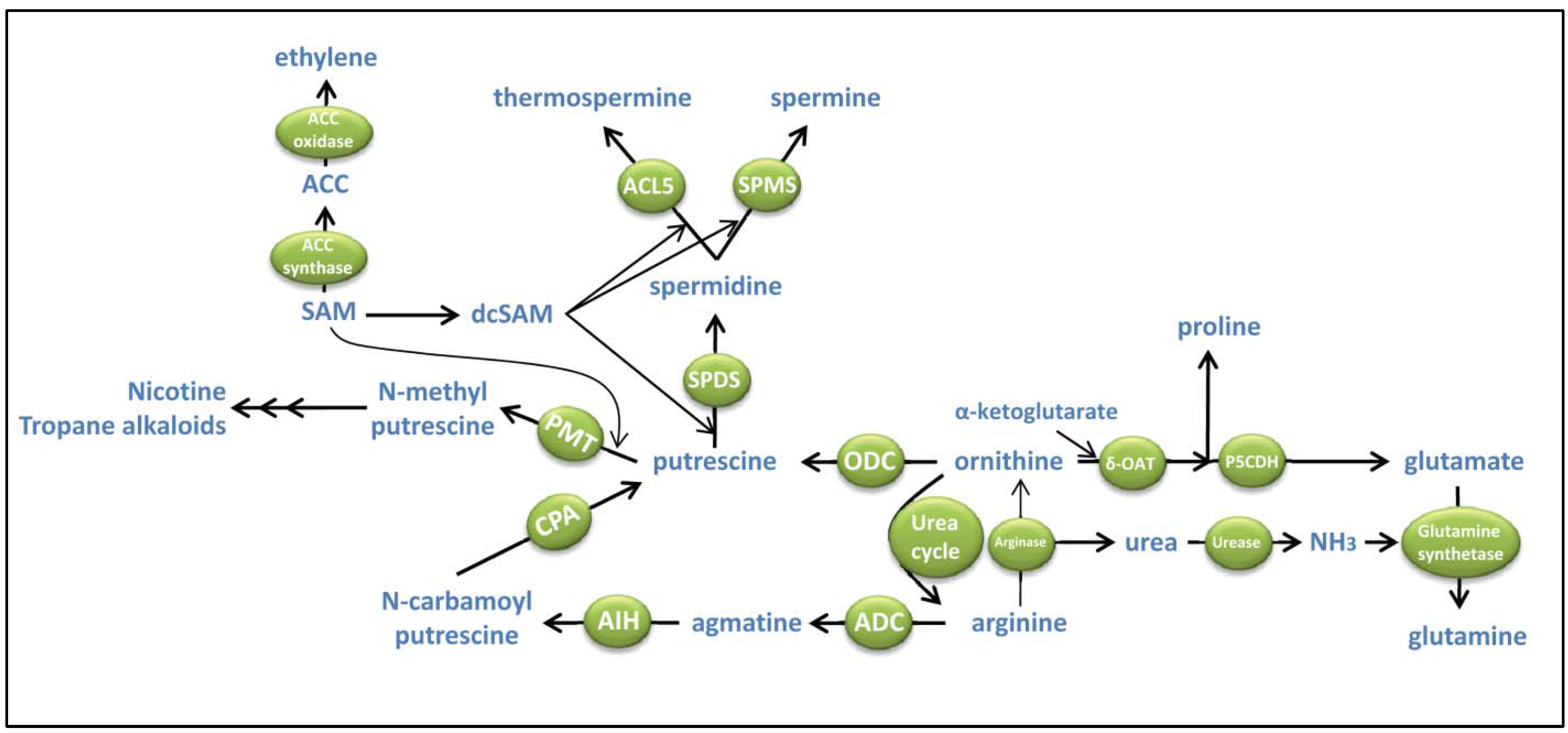
2. Polyamine Biosynthesis and Its Interaction with Other Metabolic Pathways

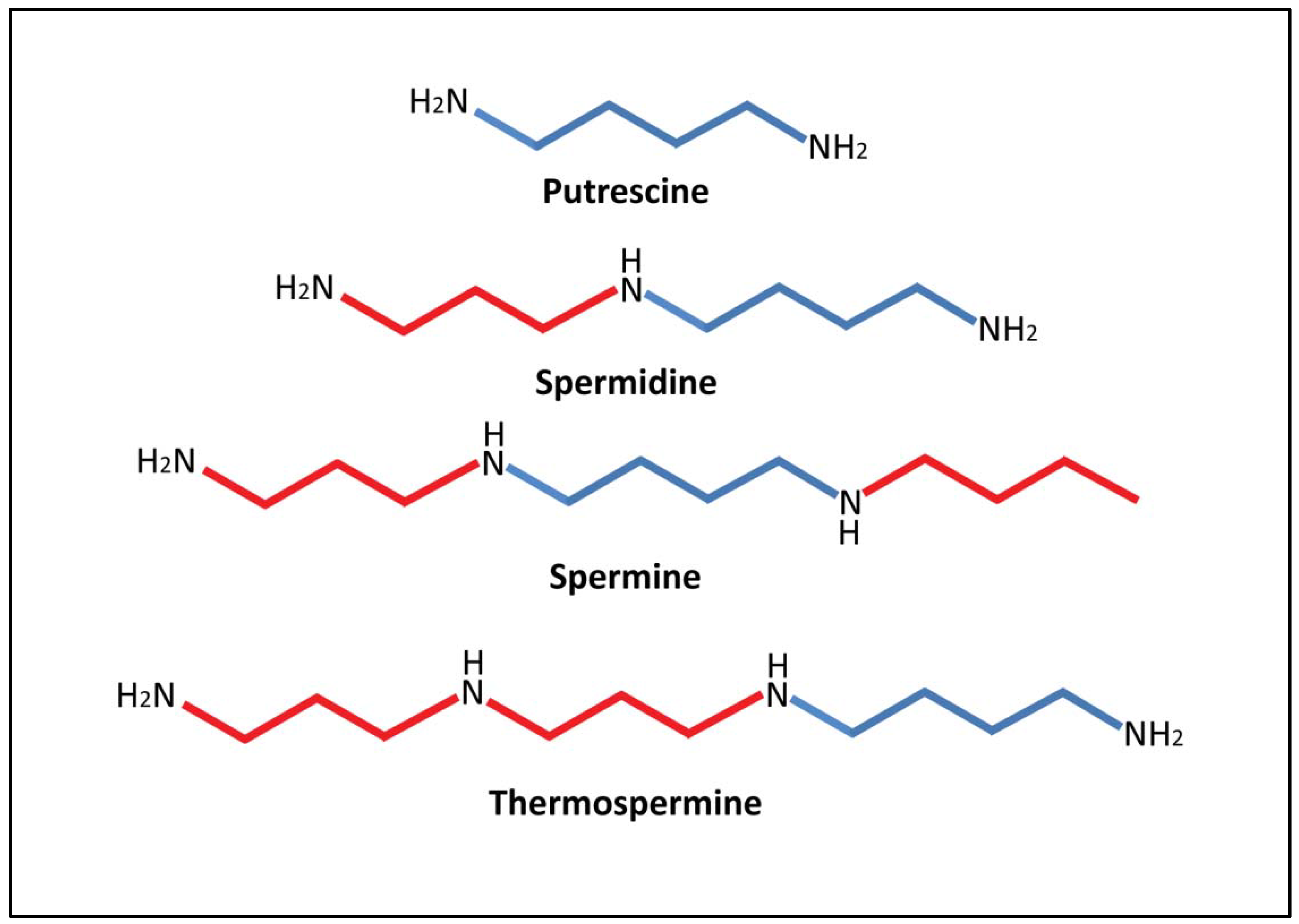
2.1. Biosynthesis of Putrescine. Interactions with Proline, Urea, NO and Alkaloid Biosynthesis

2.2. Biosynthesis of Spermidine, Spermine and Thermospermine. Interactions with S-adenosylmethionine (SAM) and ethylene
2.3. Put to Spm Canalization and Its Contribution to Stress Protection
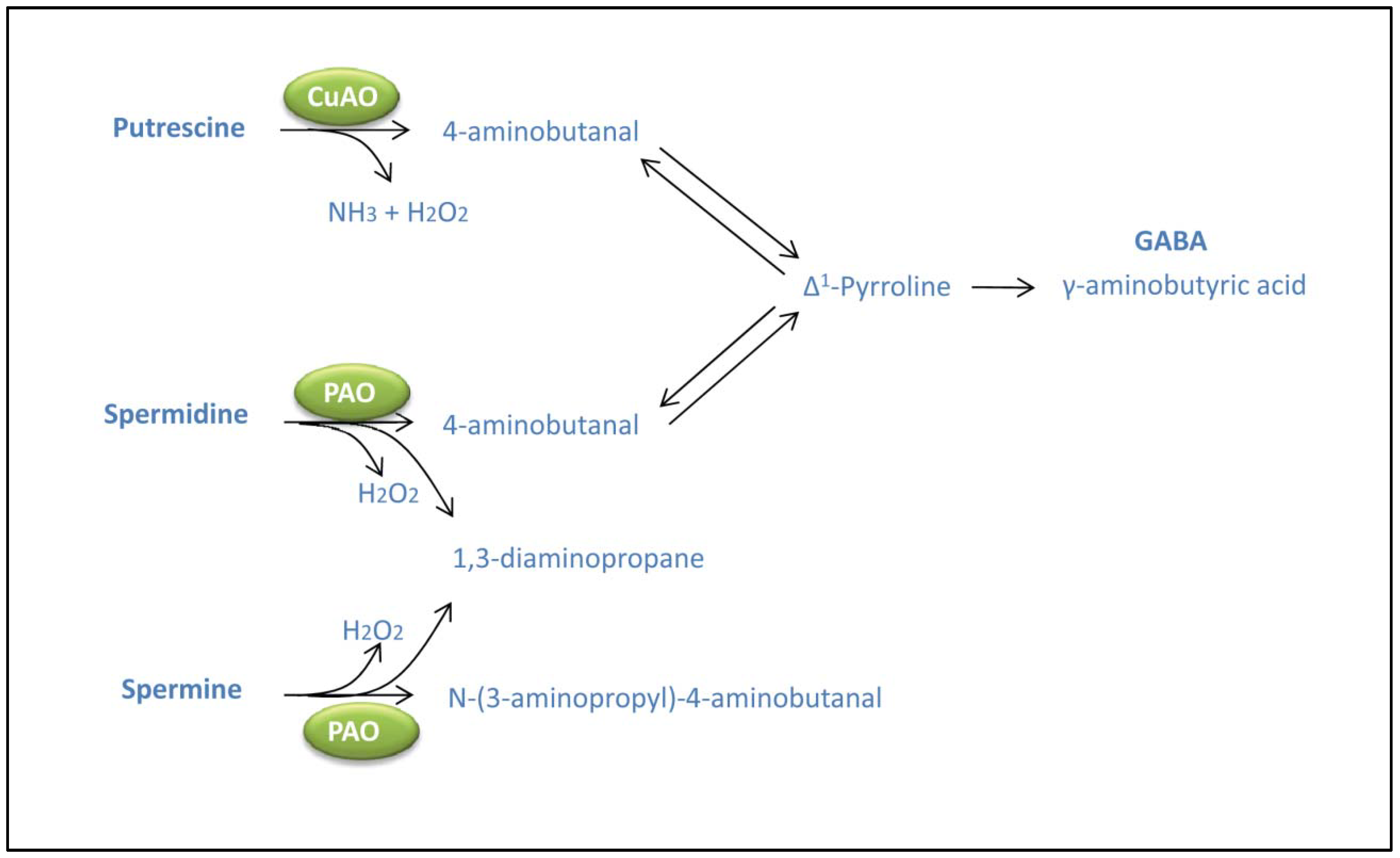
3. Polyamine Catabolism. Interactions with GABA

4. ABA Dependence of PA-Responses under Drought
5. Future Perspectives
Acknowledgments
Conflict of Interest
References
- Porter, J.R. Rising temperatures are likely to reduce crop yields. Nature 2005, 436, 174–174. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araujo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar]
- Bhatnagar-Mathur, P.; Vadez, V.; Sharma, K.K. Transgenic approaches for abiotic stress tolerance in plants: retrospect and prospects. Plant Cell Rep. 2008, 27, 411–424. [Google Scholar] [CrossRef]
- Alcázar, R.; Altabella, T.; Marco, F.; Bortolotti, C.; Reymond, M.; Koncz, C.; Carrasco, P.; Tiburcio, A.F. Polyamines: Molecules with regulatory functions in plant abiotic stress tolerance. Planta 2010, 231, 1237–1249. [Google Scholar] [CrossRef]
- Martin-Tanguy, J. Conjugated polyamines and reproductive development: Biochemical, molecular and physiological approaches. Physiol. Plant. 1997, 100, 675–688. [Google Scholar] [CrossRef]
- Hanfrey, C.; Sommer, S.; Mayer, M.J.; Burtin, D.; Michael, A.J. Arabidopsis polyamine biosynthesis: absence of ornithine decarboxylase and the mechanism of arginine decarboxylase activity. Plant J. 2001, 27, 551–560. [Google Scholar] [CrossRef]
- Murakami, Y.; Matsufuji, S.; Kameji, T.; Hayashi, S.; Igarashi, K.; Tamura, T.; Tanaka, K.; Ichihara, A. Ornithine decarboxylase is degraded by the 26s-proteasome without ubiquitination. Nature 1992, 360, 597–599. [Google Scholar]
- Kahana, C. Ubiquitin dependent and independent protein degradation in the regulation of cellular polyamines. Amino Acids 2007, 33, 225–230. [Google Scholar] [CrossRef]
- Alcázar, R.; García-Martínez, J.L.; Cuevas, J.C.; Tiburcio, A.F.; Altabella, T. Overexpression of ADC2 in Arabidopsis induces dwarfism and late-flowering through GA deficiency. Plant J. 2005, 43, 425–436. [Google Scholar] [CrossRef]
- Alcázar, R.; Planas, J.; Saxena, T.; Zarza, X.; Bortolotti, C.; Cuevas, J.; Bitrián, M.; Tiburcio, A.; Altabella, T. Putrescine accumulation confers drought tolerance in transgenic Arabidopsis plants over-expressing the homologous Arginine decarboxylase 2 gene. Plant Physiol. Biochem. 2010, 48, 547–522. [Google Scholar] [CrossRef]
- Krumpelman, P.M.; Freyermuth, S.K.; Cannon, J.F.; Fink, G.R.; Polacco, J.C. Nucleotide sequence of Arabidopsis thaliana arginase expressed in yeast. Plant Physiol. 1995, 107, 1479–1480. [Google Scholar]
- Brownfield, D.L.; Todd, C.D.; Deyholos, M.K. Analysis of Arabidopsis arginase gene transcription patterns indicates specific biological functions for recently diverged paralogs. Plant Mol. Biol. 2008, 67, 429–440. [Google Scholar] [CrossRef]
- Flores, T.; Todd, C.D.; Tovar-Mendez, A.; Dhanoa, P.K.; Correa-Aragunde, N.; Hoyos, M.E.; Brownfield, D.M.; Mullen, R.T.; Lamattina, L.; Polacco, J.C. Arginase-negative mutants of Arabidopsis exhibit increased nitric oxide signaling in root development. Plant Physiol. 2008, 147, 1936–1946. [Google Scholar] [CrossRef]
- Brauc, S.; De Vooght, E.; Claeys, M.; Geuns, J.M.C.; Hofte, M.; Angenon, G. Overexpression of arginase in Arabidopsis thaliana influences defence responses against Botrytis cinerea. Plant Biol. 2012, 14, 39–45. [Google Scholar]
- Witte, C.-P. Urea metabolism in plants. Plant Sci. 2011, 180, 431–438. [Google Scholar]
- Funck, D.; Stadelhofer, B.; Koch, W. Ornithine-delta-aminotransferase is essential for arginine catabolism but not for proline biosynthesis. BMC Plant Biol. 2008, 8. [Google Scholar]
- Hancock, J.T.; Neill, S.J.; Wilson, I.D. Nitric oxide and ABA in the control of plant function. Plant Sci. 2011, 181, 555–559. [Google Scholar]
- Crawford, N.M.; Galli, M.; Tischner, R.; Heimer, Y.M.; Okamoto, M.; Mack, A. Plant nitric oxide synthase: back to square one—Response. Trends Plant Sci. 2006, 11, 526–527. [Google Scholar] [CrossRef]
- Tun, N.N.; Santa-Catarina, C.; Begum, T.; Silveira, V.; Handro, W.; Floh, E.I.S.; Scherer, G.F.E. Polyamines induce rapid biosynthesis of nitric oxide (NO) in Arabidopsis thaliana seedlings. Plant Cell Physiol. 2006, 47, 346–354. [Google Scholar] [CrossRef]
- Yamasaki, H.; Cohen, M.F. NO signal at the crossroads: polyamine-induced nitric oxide synthesis in plants? Trends Plant Sci. 2006, 11, 522–524. [Google Scholar] [CrossRef]
- Biastoff, S.; Brandt, W.; Draeger, B. Putrescine N-methyltransferase - The start for alkaloids. Phytochemistry 2009, 70, 1708–1718. [Google Scholar] [CrossRef]
- Ge, C.M.; Cui, X.; Wang, Y.H.; Hu, Y.X.; Fu, Z.M.; Zhang, D.F.; Cheng, Z.K.; Li, J.Y. BUD2, encoding an S-adenosylmethionine decarboxylase, is required for Arabidopsis growth and development. Cell Res. 2006, 16, 446–456. [Google Scholar] [CrossRef]
- Roje, S. S-Adenosyl-L-methionine: Beyond the universal methyl group donor. Phytochemistry 2006, 67, 1686–1698. [Google Scholar] [CrossRef]
- Pegg, A.E. S-Adenosylmethionine decarboxylase. In Essays in Biochemistry, Vol 46: The Polyamines: Small Molecules in the Omics Era; Wallace, H., Ed.; Portland Press Ltd: London, UK, 2009; Volume 46, pp. 25–45. [Google Scholar]
- Herbik, A.; Koch, G.; Mock, H.P.; Dushkov, D.; Czihal, A.; Thielmann, J.; Stephan, U.W.; Baumlein, H. Isolation, characterization and cDNA cloning of nicotianamine synthase from barley - A key enzyme for iron homeostasis in plants. Eur. J. Biochem. 1999, 265, 231–239. [Google Scholar] [CrossRef]
- Mizuno, D.; Higuchi, K.; Sakamoto, T.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Three nicotianamine synthase genes isolated from maize are differentially regulated by iron nutritional status. Plant Physiol. 2003, 132, 1989–1997. [Google Scholar] [CrossRef]
- Fluhr, R.; Mattoo, A.K. Ethylene - Biosynthesis and perception. Crc. Cr. Rev. Plant Sci. 1996, 15, 479–523. [Google Scholar]
- Yang, S.F.; Hoffman, N.E. Ethylene biosynthesis and its regulation in higher-plants. Ann. Rev. Plant Phys. 1984, 35, 155–189. [Google Scholar] [CrossRef]
- Mattoo, A.K.; Suttle, J.C. The Plant Hormone Ethylene; CRC Press: Boca Raton: FL, USA, 1991; pp. 21–42. [Google Scholar]
- Evans, P.T.; Malmberg, R.L. Do polyamines have roles in plant development. Ann. Rev. Plant Phys. 1989, 40, 235–269. [Google Scholar] [CrossRef]
- Apelbaum, A.; Burgoon, A.C.; Anderson, J.D.; Lieberman, M.; Benarie, R.; Mattoo, A.K. Polyamines inhibit biosynthesis of ethylene in higher-plant tissue and fruit protoplasts. Plant Physiol. 1981, 68, 453–456. [Google Scholar] [CrossRef]
- Apelbaum, A.; Goldlust, A.; Icekson, I. Control by ethylene of arginine decarboxylase activity in pea-seedlings and its implication for hormonal-regulation of plant-growth. Plant Physiol. 1985, 79, 635–640. [Google Scholar] [CrossRef]
- Suttle, J.C. Effect of polyamines on ethylene production. Phytochemistry 1981, 20, 1477–1480. [Google Scholar] [CrossRef]
- Li, N.; Parsons, B.L.; Liu, D.R.; Mattoo, A.K. Accumulation of wound-inducible acc synthase transcript in tomato fruit is inhibited by salicylic-acid and polyamines. Plant Mol. Biol. 1992, 18, 477–487. [Google Scholar] [CrossRef]
- Roberts, D.R.; Walker, M.A.; Thompson, J.E.; Dumbroff, E.B. The effects of inhibitors of polyamine and ethylene biosynthesis on senescence, ethylene production and polyamine levels in cut carnation flowers. Plant Cell Physiol. 1984, 25, 315–322. [Google Scholar]
- Mehta, R.A.; Cassol, T.; Li, N.; Ali, N.; Handa, A.K.; Mattoo, A.K. Engineered polyamine accumulation in tomato enhances phytonutrient content, juice quality, and vine life. Nat. Biotechnol. 2002, 20, 613–618. [Google Scholar] [CrossRef]
- Nambeesan, S.; AbuQamar, S.; Laluk, K.; Mattoo, A.K.; Mickelbart, M.V.; Ferruzzi, M.G.; Mengiste, T.; Handa, A.K. Polyamines attenuate ethylene-mediated defense responses to abrogate resistance to Botrytis cinerea in tomato. Plant Physiol. 2012, 158, 1034–1045. [Google Scholar] [CrossRef]
- Knott, J.M.; Romer, P.; Sumper, M. Putative spermine synthases from Thalassiosira pseudonana and Arabidopsis thaliana synthesize thermospermine rather than spermine. FEBS Lett. 2007, 581, 3081–3086. [Google Scholar] [CrossRef]
- Hanzawa, Y.; Takahashi, T.; Michael, A.J.; Burtin, D.; Long, D.; Pineiro, M.; Coupland, G.; Komeda, Y. ACAULIS5, an Arabidopsis gene required for stem elongation, encodes a spermine synthase. EMBO J. 2000, 19, 4248–4256. [Google Scholar] [CrossRef]
- Imai, A.; Akiyama, T.; Kato, T.; Sato, S.; Tabata, S.; Yamamoto, K.T.; Takahashi, T. Spermine is not essential for survival of Arabidopsis. FEBS Lett. 2004, 556, 148–152. [Google Scholar] [CrossRef]
- Imai, A.; Matsuyama, T.; Hanzawa, Y.; Akiyama, T.; Tamaoki, M.; Saji, H.; Shirano, Y.; Kato, T.; Hayashi, H.; Shibata, D.; Tabata, S.; Komeda, Y.; Takahashi, T. Spermidine synthase genes are essential for survival of Arabidopsis. Plant Physiol. 2004, 135, 1565–1573. [Google Scholar]
- Urano, K.; Hobo, T.; Shinozaki, K. Arabidopsis ADC genes involved in polyamine biosynthesis are essential for seed development. FEBS Lett. 2005, 579, 1557–1564. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Takahashi, Y.; Berberich, T.; Imai, A.; Miyazaki, A.; Takahashi, T.; Michael, A.; Kusano, T. The polyamine spermine protects against high salt stress in Arabidopsis thaliana. FEBS Lett. 2006, 580, 6783–6788. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Takahashi, Y.; Berberich, T.; Imai, A.; Takahashi, T.; Michael, A.J.; Kusano, T. A protective role for the polyamine spermine against drought stress in Arabidopsis. Biochem. Biophys. Res. Commun. 2007, 352, 486–490. [Google Scholar] [CrossRef]
- Sagor, G.H.M.; Takahashi, H.; Niitsu, M.; Takahashi, Y.; Berberich, T.; Kusano, T. Exogenous thermospermine has an activity to induce a subset of the defense genes and restrict cucumber mosaic virus multiplication in Arabidopsis thaliana. Plant Cell Rep. 2012, 31, 1227–1232. [Google Scholar] [CrossRef]
- Alcázar, R.; Bitrián, M.; Bartels, D.; Koncz, C.; Altabella, T.; Tiburcio, A.F. Polyamine metabolic canalization in response to drought stress in Arabidopsis and the resurrection plant Craterostigma plantagineum. Plant Signal. Behav. 2011, 6, 243–250. [Google Scholar] [CrossRef]
- Cona, A.; Rea, G.; Angelini, R.; Federico, R.; Tavladoraki, P. Functions of amine oxidases in plant development and defence. Trends Plant Sci. 2006, 11, 80–88. [Google Scholar] [CrossRef]
- Moschou, P.N.; Paschalidis, K.A.; Roubelakis-Angelakis, K.A. Plant polyamine catabolism: The state of the art. Plant Signal. Behav. 2008, 3, 1061–1066. [Google Scholar] [CrossRef]
- Roberts, M.R. Does GABA act as a signal in plants? Hints from molecular studies. Plant Signal. Behav. 2007, 2, 408–409. [Google Scholar] [CrossRef]
- Bouché, N.; Fromm, H. GABA in plants: just a metabolite? Trends Plant Sci. 2004, 9, 110–115. [Google Scholar] [CrossRef]
- Xing, S.G.; Jun, Y.B.; Hau, Z.W.; Liang, L.Y. Higher accumulation of γ-aminobutyric acid induced by salt stress through stimulating the activity of diamine oxidases in Glycine max (L.) Merr. roots. Plant Physiol. Biochem. 2007, 45, 560–566. [Google Scholar] [CrossRef]
- Yoda, H.; Yamaguchi, Y.; Sano, H. Induction of hypersensitive cell death by hydrogen peroxide produced through polyamine degradation in tobacco plants. Plant Physiol. 2003, 132, 1973–1981. [Google Scholar] [CrossRef]
- Yoda, H.; Fujimura, K.; Takahashi, H.; Munemura, I.; Uchimiya, H.; Sano, H. Polyamines as a common source of hydrogen peroxide in host- and nonhost hypersensitive response during pathogen infection. Plant Mol. Biol. 2009, 70, 103–112. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef]
- Alcázar, R.; Cuevas, J.C.; Patrón, M.; Altabella, T.; Tiburcio, A.F. Abscisic acid modulates polyamine metabolism under water stress in Arabidopsis thaliana. Physiol. Plant. 2006, 128, 448–455. [Google Scholar] [CrossRef]
- Alcázar, R.; Marco, F.; Cuevas, J.C.; Patrón, M.; Ferrando, A.; Carrasco, P.; Tiburcio, A.F.; Altabella, T. Involvement of polyamines in plant response to abiotic stress. Biotechnol Lett. 2006, 28, 1867–1876. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bitrián, M.; Zarza, X.; Altabella, T.; Tiburcio, A.F.; Alcázar, R. Polyamines under Abiotic Stress: Metabolic Crossroads and Hormonal Crosstalks in Plants. Metabolites 2012, 2, 516-528. https://doi.org/10.3390/metabo2030516
Bitrián M, Zarza X, Altabella T, Tiburcio AF, Alcázar R. Polyamines under Abiotic Stress: Metabolic Crossroads and Hormonal Crosstalks in Plants. Metabolites. 2012; 2(3):516-528. https://doi.org/10.3390/metabo2030516
Chicago/Turabian StyleBitrián, Marta, Xavier Zarza, Teresa Altabella, Antonio F. Tiburcio, and Rubén Alcázar. 2012. "Polyamines under Abiotic Stress: Metabolic Crossroads and Hormonal Crosstalks in Plants" Metabolites 2, no. 3: 516-528. https://doi.org/10.3390/metabo2030516
APA StyleBitrián, M., Zarza, X., Altabella, T., Tiburcio, A. F., & Alcázar, R. (2012). Polyamines under Abiotic Stress: Metabolic Crossroads and Hormonal Crosstalks in Plants. Metabolites, 2(3), 516-528. https://doi.org/10.3390/metabo2030516