
Effects of Storage Time on Glycolysis in Donated Human Blood Units

Abstract
:1. Introduction
2. Results
2.1. Time Courses of Metabolite Levels.
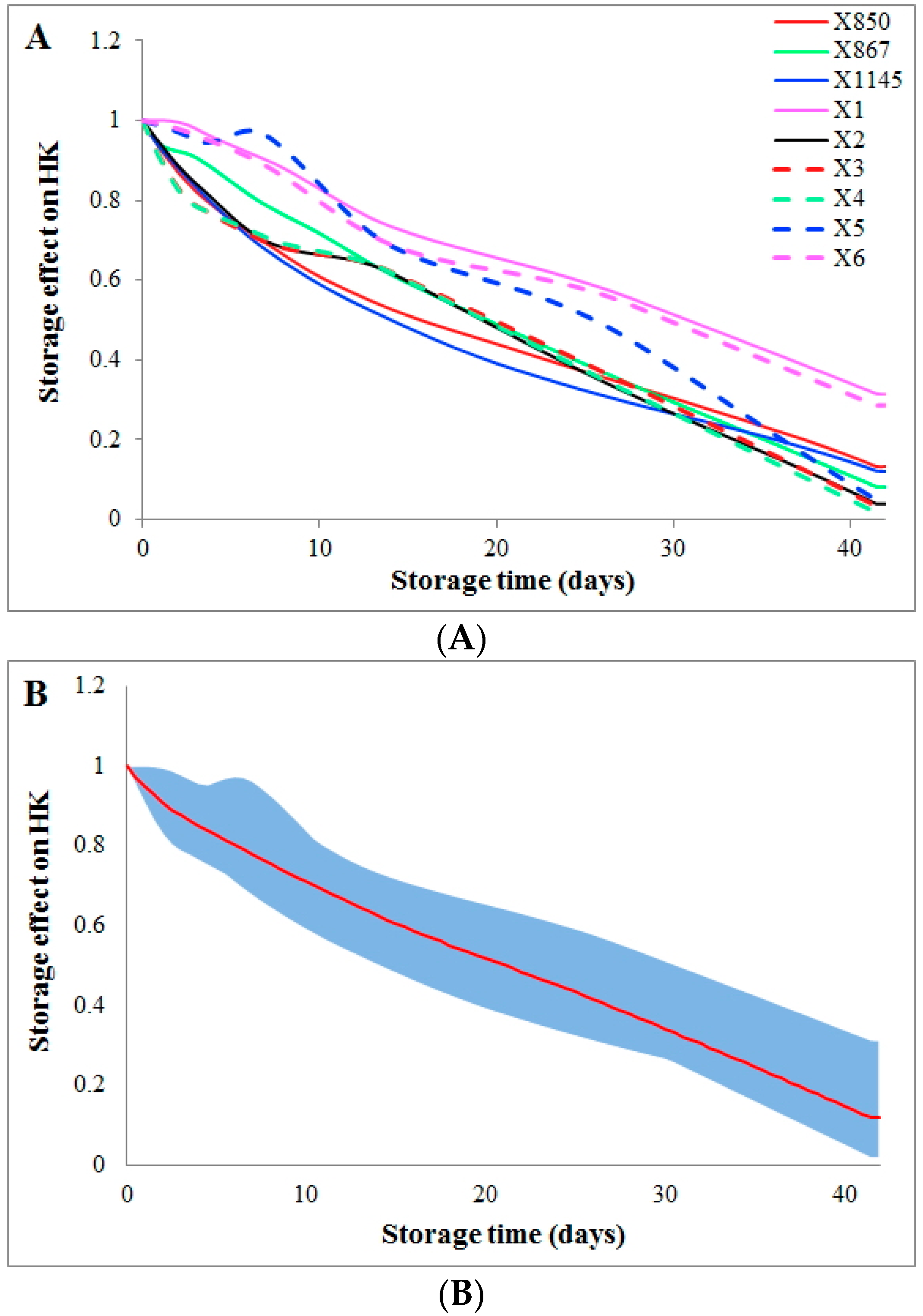
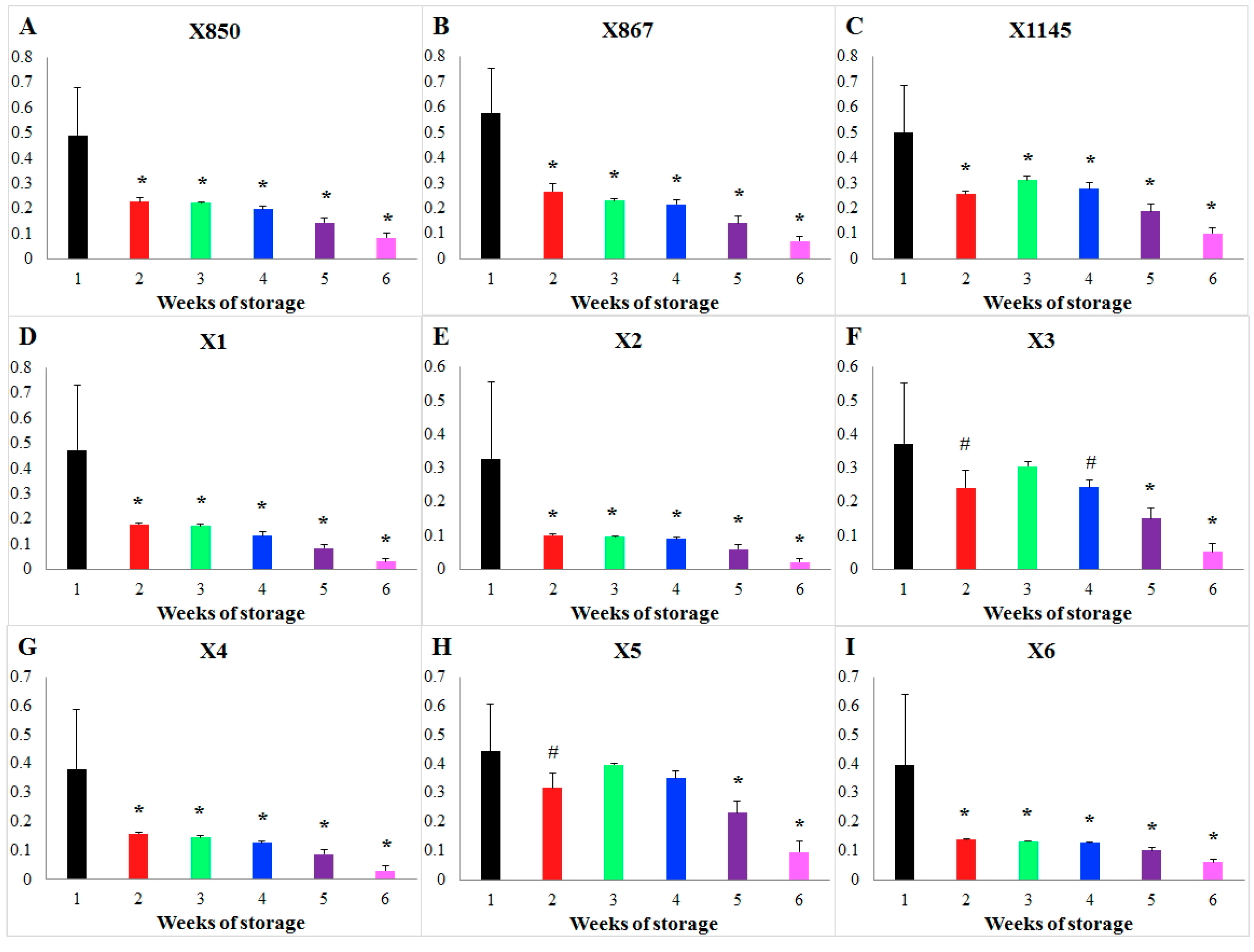
2.2. Effect of Storage Time
2.3. Influence of Donation Batch
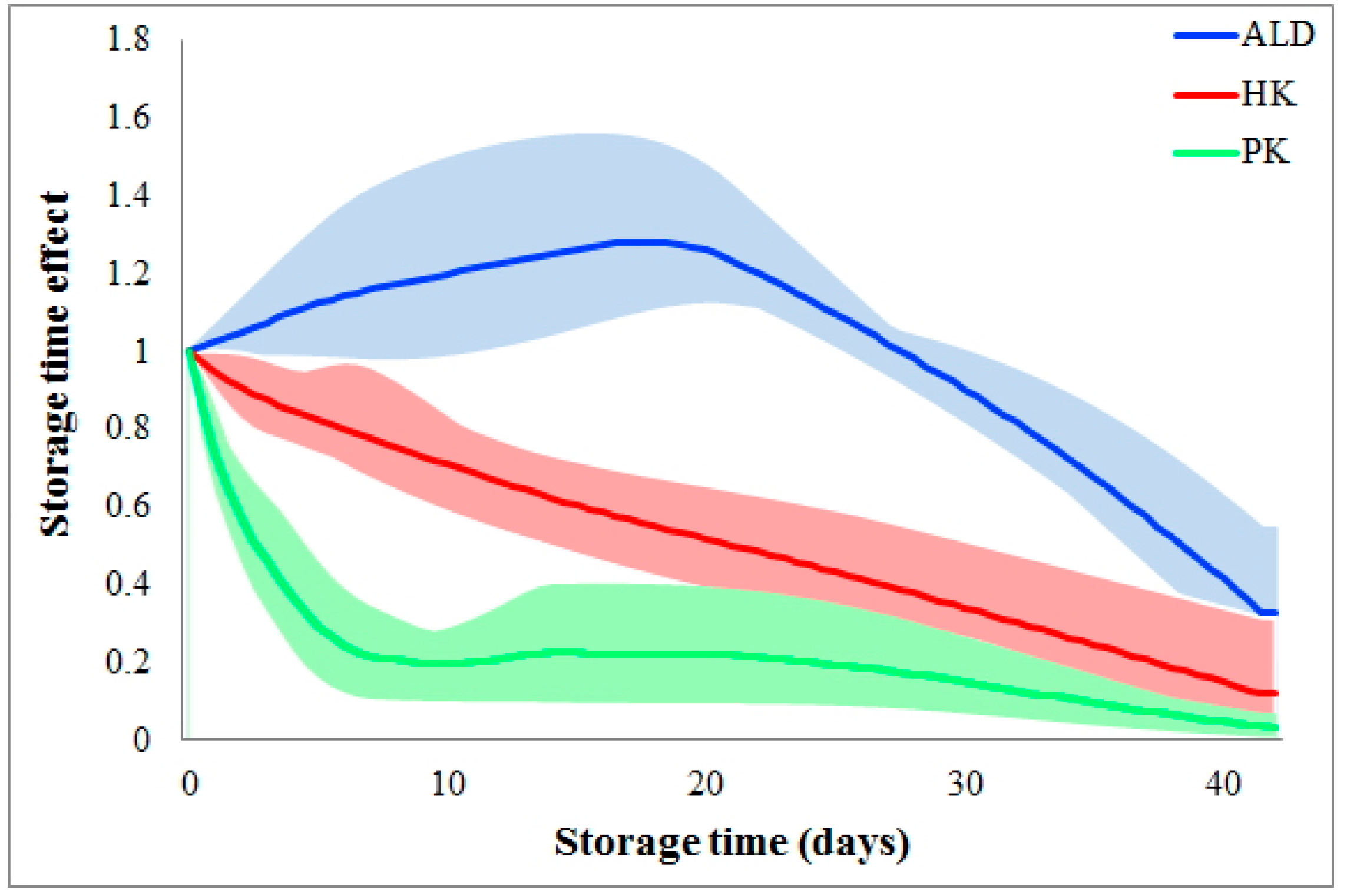
2.4. Comparison of Storage Time Effects among Various Reactions
2.5. Influence of Selecting Particular Kinetic Models
2.6. Summary of Storage Time Effects
3. Discussion
4. Materials and Methods
4.1. Metabolomics Data
4.2. Factors Influencing Metabolic Flux Dynamics during Storage
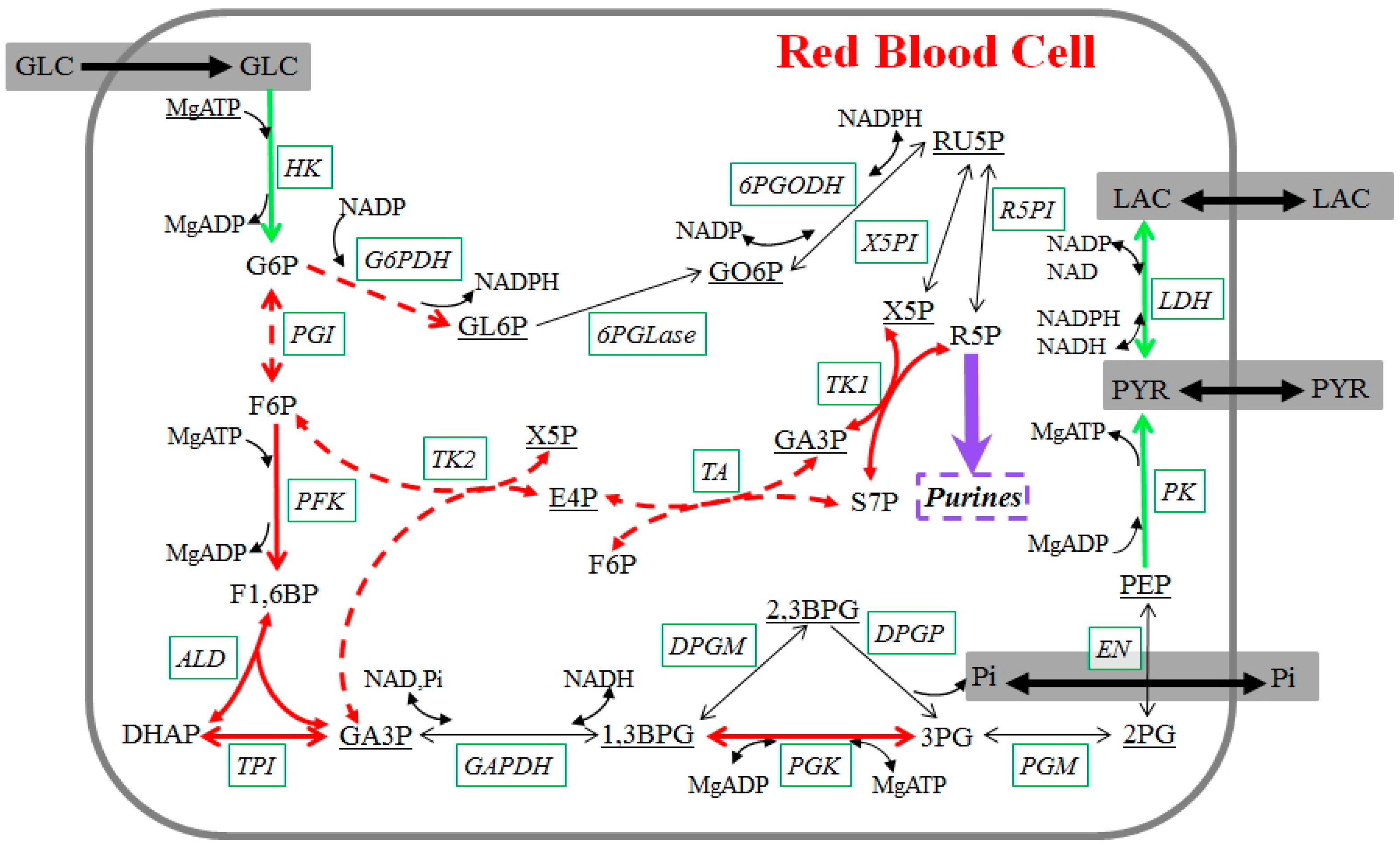
4.3. Pathways of Importance
4.4. Calibration of Metabolomics Data
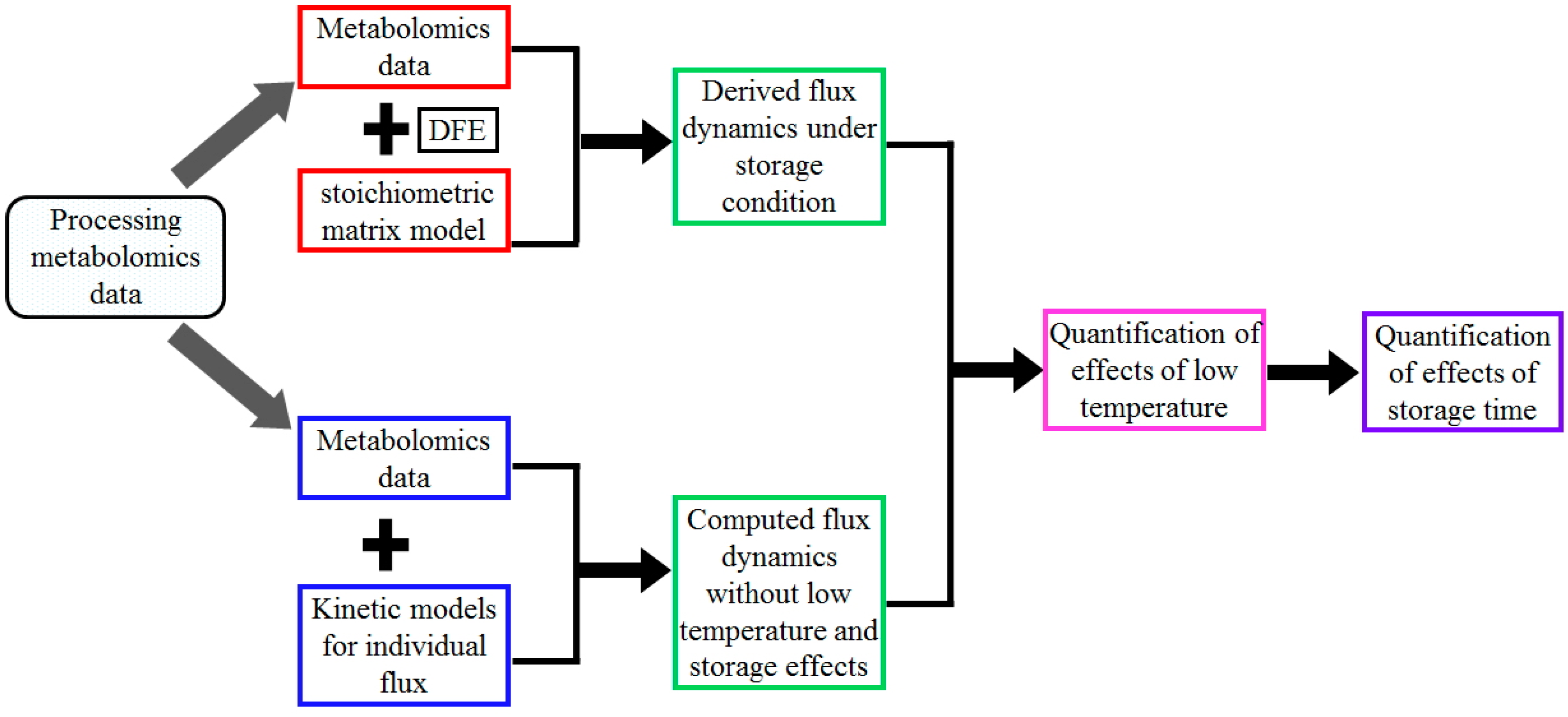
4.5. Strategy for the Quantification of Storage Time Effects on Flux Dynamics in Stored Blood
4.6. Inference of Flux Dynamics from Metabolomics Data under Storage Conditions
4.7. Quantification of Fluxes under Normal Physiological Conditions and Comparison with Storage Conditions
4.8. Effect of Storage Time on Flux Dynamics
4.9. Influence of Other Factors on the Dynamic Quantification of Storage Time Effects
5. Conclusions
Supplementary Materials
Acknowledgments
Authors Contributions
Conflicts of Interest
Abbreviations
References
- Levit, K.R.K.; Elixhauser, A.; Stranges, E.; Kassed, C.; Coffey, R. HCUP Facts and Figures: Statistics on Hospital-Based Care in the United States in 2005; Agency for Healthcare Research and Quality: Rockville, MD, USA, 2007. [Google Scholar]
- Dumont, L.J.; AuBuchon, J.P. Evaluation of proposed FDA criteria for the evaluation of radiolabeled red cell recovery trials. Transfusion 2008, 48, 1053–1060. [Google Scholar] [CrossRef] [PubMed]
- Van’t Erve, T.J.; Wagner, B.A.; Martin, S.M.; Knudson, C.M.; Blendowski, R.; Keaton, M.; Holt, T.; Hess, J.R.; Buettner, G.R.; Ryckman, K.K.; et al. The heritability of metabolite concentrations in stored human red blood cells. Transfusion 2014, 54, 2055–2063. [Google Scholar] [CrossRef] [PubMed]
- Van’t Erve, T.J.; Wagner, B.A.; Martin, S.M.; Knudson, C.M.; Blendowski, R.; Keaton, M.; Holt, T.; Hess, J.R.; Buettner, G.R.; Ryckman, K.K.; et al. The heritability of hemolysis in stored human red blood cells. Transfusion 2015, 55, 1178–1185. [Google Scholar] [CrossRef] [PubMed]
- Meryman, H.T.; Hornblower, M.L.; Syring, R.L. Prolonged storage of red cells at 4 degrees C. Transfusion 1986, 26, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Gerber, G.; Preissler, H.; Heinrich, R.; Rapoport, S.M. Hexokinase of human erythrocytes. Purification, kinetic model and its application to the conditions in the cell. Eur. J. Biochem. 1974, 45, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Prudent, M.; Rochat, B.; Goffaux, G.; Marvin, L.; Stauber, F.; Tissot, J.; Jolicoeur, M.; Lion, N. Dynamic Metabolic Flux Analysis of Stored Red Blood Cells. Transfusion 2016, 56 (Suppl. S4), 25A. [Google Scholar]
- Otto, M.; Heinrich, R.; Kuhn, B.; Jacobasch, G. A mathematical model for the influence of fructose 6-phosphate, ATP, potassium, ammonium and magnesium on the phosphofructokinase from rat erythrocytes. Eur. J. Biochem. 1974, 49, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Mulquiney, P.J.; Kuchel, P.W. Model of 2,3-bisphosphoglycerate metabolism in the human erythrocyte based on detailed enzyme kinetic equations: Equations and parameter refinement. Biochem. J. 1999, 342, 581–596. [Google Scholar] [CrossRef] [PubMed]
- Blood Products Advisory Committee. 2004. Available online: http://www.fda.gov/ohrms/dockets/ac/04/transcripts/2004-4057t1.htm (accessed on 1 March 2016).
- Roback, J.D.; Josephson, C.D.; Waller, E.K.; Newman, J.L.; Karatela, S.; Uppal, K.; Jones, D.P.; Zimring, J.C.; Dumont, L.J. Metabolomics of ADSOL (AS-1) red blood cell storage. Transfus. Med. Rev. 2014, 28, 41–55. [Google Scholar] [CrossRef] [PubMed]
- Bordbar, A.; Johansson, P.I.; Paglia, G.; Harrison, S.J.; Wichuk, K.; Magnusdottir, M.; Valgeirsdottir, S.; Gybel-Brask, M.; Ostrowski, S.R.; Palsson, S.; et al. Identified metabolic signature for assessing red blood cell unit quality is associated with endothelial damage markers and clinical outcomes. Transfusion 2016, 56, 852–862. [Google Scholar] [CrossRef] [PubMed]
- Paglia, G.; D’Alessandro, A.; Rolfsson, O.; Sigurjonsson, O.E.; Bordbar, A.; Palsson, S.; Nemkov, T.; Hansen, K.C.; Gudmundsson, S.; Palsson, B.O. Biomarkers defining the metabolic age of red blood cells during cold storage. Blood 2016, 128, e43–e50. [Google Scholar] [CrossRef] [PubMed]
- Gevi, F.; D’Alessandro, A.; Rinalducci, S.; Zolla, L. Alterations of red blood cell metabolome during cold liquid storage of erythrocyte concentrates in CPD-SAGM. J. Proteom. 2012, 76, 168–180. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.; D’Amici, G.M.; Vaglio, S.; Zolla, L. Time-course investigation of SAGM-stored leukocyte-filtered red bood cell concentrates: From metabolism to proteomics. Haematologica 2012, 97, 107–115. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.; Nemkov, T.; Hansen, K.C.; Szczepiorkowski, Z.M.; Dumont, L.J. Red blood cell storage in additive solution-7 preserves energy and redox metabolism: A metabolomics approach. Transfusion 2015, 55, 2955–2966. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.; Nemkov, T.; Yoshida, T.; Bordbar, A.; Palsson, B.O.; Hansen, K.C. Citrate metabolism in red blood cells stored in additive solution-3. Transfusion 2016. [Google Scholar] [CrossRef] [PubMed]
- Dumont, L.J.; D’Alessandro, A.; Szczepiorkowski, Z.M.; Yoshida, T. CO2-dependent metabolic modulation in red blood cells stored under anaerobic conditions. Transfusion 2016, 56, 392–403. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.; Kriebardis, A.G.; Rinalducci, S.; Antonelou, M.H.; Hansen, K.C.; Papassideri, I.S.; Zolla, L. An update on red blood cell storage lesions, as gleaned through biochemistry and omics technologies. Transfusion 2015, 55, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Zimring, J.C.; Smith, N.; Stowell, S.R.; Johnsen, J.M.; Bell, L.N.; Francis, R.O.; Hod, E.A.; Hendrickson, J.E.; Roback, J.D.; Spitalnik, S.L. Strain-specific red blood cell storage, metabolism, and eicosanoid generation in a mouse model. Transfusion 2014, 54, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Felcyn, J.R.; Odem-Davis, K.; Zimring, J.C. Bioactive lipids accumulate in stored red blood cells despite leukoreduction: A targeted metabolomics study. Transfusion 2016, 56, 2560–2570. [Google Scholar] [CrossRef] [PubMed]
- De Wolski, K.; Fu, X.; Dumont, L.J.; Roback, J.D.; Waterman, H.; Odem-Davis, K.; Howie, H.L.; Zimring, J.C. Metabolic pathways that correlate with post-transfusion circulation of stored murine red blood cells. Haematologica 2016, 101, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Delobel, J.; Prudent, M.; Rubin, O.; Crettaz, D.; Tissot, J.D.; Lion, N. Subcellular fractionation of stored red blood cells reveals a compartment-based protein carbonylation evolution. J. Proteom. 2012, 76, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Reisz, J.A.; Wither, M.J.; Dzieciatkowska, M.; Nemkov, T.; Issaian, A.; Yoshida, T.; Dunham, A.J.; Hill, R.C.; Hansen, K.C.; D'Alessandro, A. Oxidative modifications of glyceraldehyde 3-phosphate dehydrogenase regulate metabolic reprogramming of stored red blood cells. Blood 2016, 128, e32–e42. [Google Scholar] [CrossRef] [PubMed]
- Nishino, T.; Yachie-Kinoshita, A.; Hirayama, A.; Soga, T.; Suematsu, M.; Tomita, M. Dynamic simulation and metabolome analysis of long-term erythrocyte storage in adenine-guanosine solution. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Tzounakas, V.L.; Kriebardis, A.G.; Georgatzakou, H.T.; Foudoulaki-Paparizos, L.E.; Dzieciatkowska, M.; Wither, M.J.; Nemkov, T.; Hansen, K.C.; Papassideri, I.S.; D'Alessandro, A.; et al. Glucose 6-phosphate dehydrogenase deficient subjects may be better “storers” than donors of red blood cells. Free Radic. Biol. Med. 2016, 96, 152–165. [Google Scholar] [CrossRef] [PubMed]
- Tzounakas, V.L.; Kriebardis, A.G.; Georgatzakou, H.T.; Foudoulaki-Paparizos, L.E.; Dzieciatkowska, M.; Wither, M.J.; Nemkov, T.; Hansen, K.C.; Papassideri, I.S.; D'Alessandro, A.; et al. Data on how several physiological parameters of stored red blood cells are similar in glucose 6-phosphate dehydrogenase deficient and sufficient donors. Data Brief 2016, 8, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Peters, A.L.; van Bruggen, R.; de Korte, D.; Van Noorden, C.J.; Vlaar, A.P. Glucose-6-phosphate dehydrogenase activity decreases during storage of leukoreduced red blood cells. Transfusion 2016, 56, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Sriyudthsak, K.; Shiraishi, F.; Hirai, M.Y. Mathematical Modeling and Dynamic Simulation of Metabolic Reaction Systems Using Metabolome Time Series Data. Front. Mol. Biosci. 2016, 3. [Google Scholar] [CrossRef] [PubMed]
- Dolatshahi, S.; Voit, E.O. Identification of Metabolic Pathway Systems. Front. Genet. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Goel, G.; Chou, I.C.; Voit, E.O. System estimation from metabolic time-series data. Bioinformatics 2008, 24, 2505–2511. [Google Scholar] [CrossRef] [PubMed]
- Nishino, T.; Yachie-Kinoshita, A.; Hirayama, A.; Soga, T.; Suematsu, M.; Tomita, M. In silico modeling and metabolome analysis of long-stored erythrocytes to improve blood storage methods. J. Biotechnol. 2009, 144, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Holzhütter, H.G.; Jacobasch, G.; Bisdorff, A. Mathematical modelling of metabolic pathways affected by an enzyme deficiency. A mathematical model of glycolysis in normal and pyruvate-kinase-deficient red blood cells. Eur. J. Biochem. 1985, 149, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.; Palsson, B.O. Metabolic dynamics in the human red cell. Part II—Interactions with the environment. J. Theor. Biol. 1989, 141, 529–545. [Google Scholar] [CrossRef]
- Price, N.D.; Reed, J.L.; Papin, J.A.; Wiback, S.J.; Palsson, B.O. Network-based analysis of metabolic regulation in the human red blood cell. J. Theor. Biol. 2003, 225, 185–194. [Google Scholar] [CrossRef]
- Mulquiney, P.J.; Bubb, W.A.; Kuchel, P.W. Model of 2,3-bisphosphoglycerate metabolism in the human erythrocyte based on detailed enzyme kinetic equations: In vivo kinetic characterization of 2,3-bisphosphoglycerate synthase/phosphatase using 13C and 31P NMR. Biochem. J. 1999, 342, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, Y.; Kinoshita, A.; Tomita, M. Dynamic simulation of red blood cell metabolism and its application to the analysis of a pathological condition. Theor. Biol. Med. Model. 2005, 2. [Google Scholar] [CrossRef] [PubMed]
- D’Amici, G.M.; Rinalducci, S.; Zolla, L. Proteomic analysis of RBC membrane protein degradation during blood storage. J. Proteome Res. 2007, 6, 3242–3455. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Kim, J.W.; Jeon, S.Y.; Park, B.K.; Han, B.G. Proteomic analysis of the effect of storage temperature on human serum. Ann. Clin. Lab. Sci. 2010, 40, 61–70. [Google Scholar] [PubMed]
- Dinkla, S.; Novotny, V.M.; Joosten, I.; Bosman, G.J. Storage-induced changes in erythrocyte membrane proteins promote recognition by autoantibodies. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Kriebardis, A.G.; Antonelou, M.H.; Stamoulis, K.E.; Economou-Petersen, E.; Margaritis, L.H.; Papassideri, I.S. Progressive oxidation of cytoskeletal proteins and accumulation of denatured hemoglobin in stored red cells. J. Cell. Mol. Med. 2007, 11, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Meyer, E.K.; Dumont, D.F.; Baker, S.; Dumont, L.J. Rejuvenation capacity of red blood cells in additive solutions over long-term storage. Transfusion 2011, 51, 1574–1579. [Google Scholar] [CrossRef] [PubMed]
- Dumont, L.J.; Herschel, L.; Roback, J.D.; Zimring, J.C.; Gray, A.D. Changes in the metabolomic profiles of stored RBC following treatment with Rejuvesol® Solution. Transfusion 2012, 52, 78A. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect on Specific Reactions | Example |
|---|---|
| Storage quickly reduces the speed of a reaction during the 1st week and then retains a small flux during the following 5 weeks. | PK |
| The flux magnitude linearly decreases during the entire 6 weeks of storage. | HK |
| Storage does not slow down a flux until after 3 weeks. | ALD |
| Except for the two donors (X3 and X5), there are significant differences in the storage effect between the 1st, 2nd, 3rd, and 4th week. | PK |
| In all donors, storage significantly changes a flux every week in comparison with the 1st week. | HK |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, Z.; Roback, J.D.; Voit, E.O. Effects of Storage Time on Glycolysis in Donated Human Blood Units. Metabolites 2017, 7, 12. https://doi.org/10.3390/metabo7020012
Qi Z, Roback JD, Voit EO. Effects of Storage Time on Glycolysis in Donated Human Blood Units. Metabolites. 2017; 7(2):12. https://doi.org/10.3390/metabo7020012
Chicago/Turabian StyleQi, Zhen, John D. Roback, and Eberhard O. Voit. 2017. "Effects of Storage Time on Glycolysis in Donated Human Blood Units" Metabolites 7, no. 2: 12. https://doi.org/10.3390/metabo7020012
APA StyleQi, Z., Roback, J. D., & Voit, E. O. (2017). Effects of Storage Time on Glycolysis in Donated Human Blood Units. Metabolites, 7(2), 12. https://doi.org/10.3390/metabo7020012