
The Effect of Season on the Metabolic Profile of the European Clam Ruditapes decussatus as Studied by 1H-NMR Spectroscopy

 ,
, 
Abstract
:1. Introduction
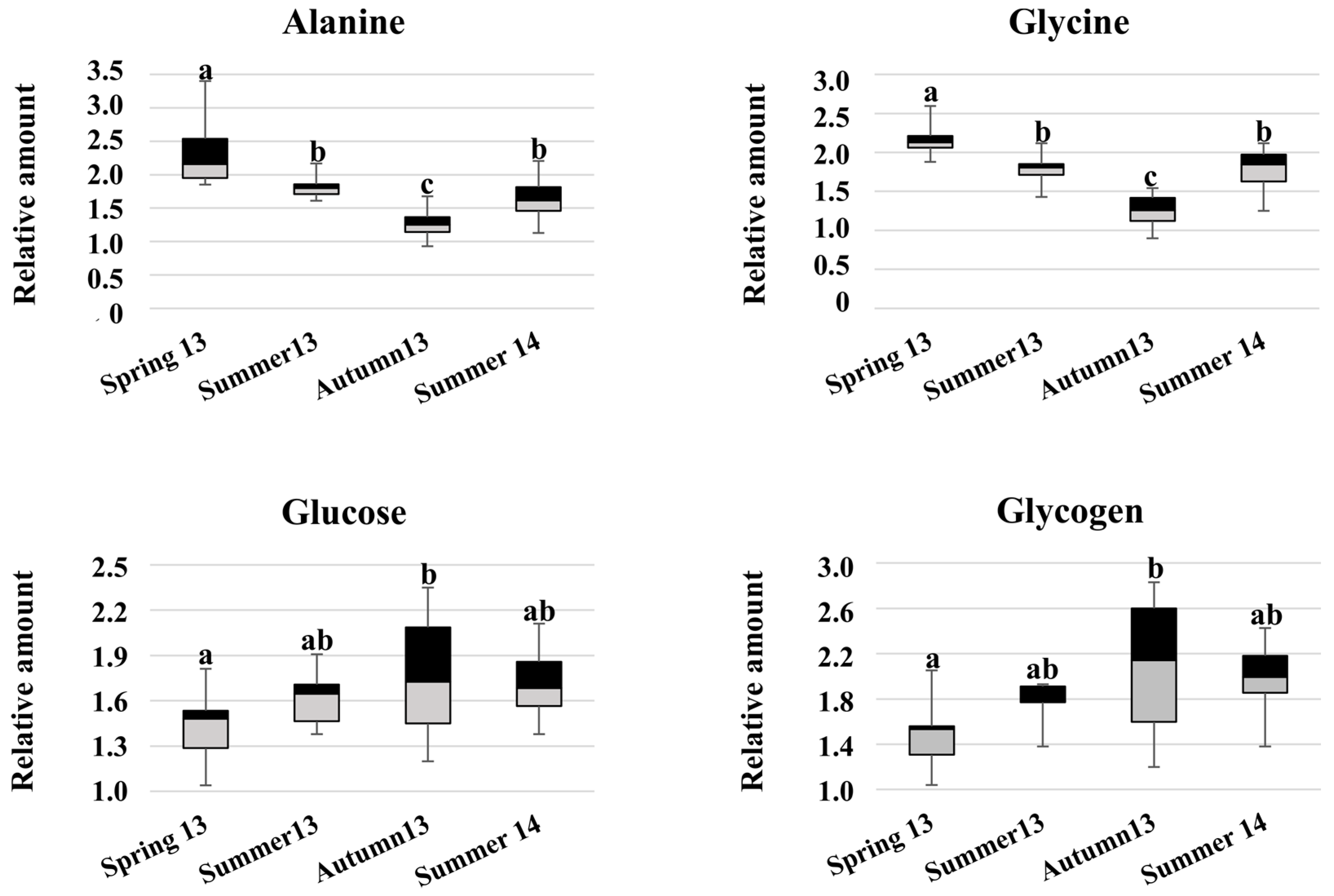
2. Results
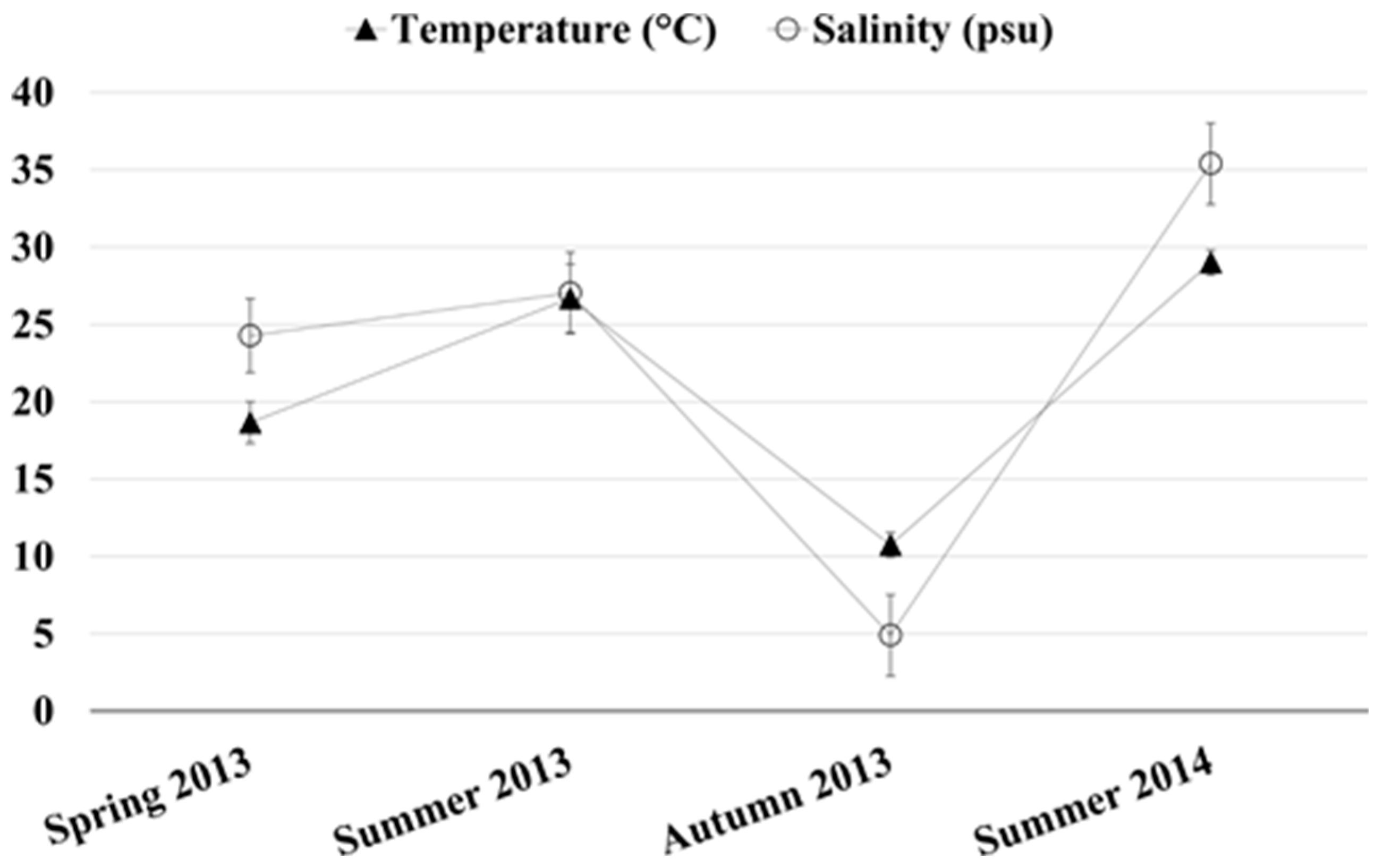
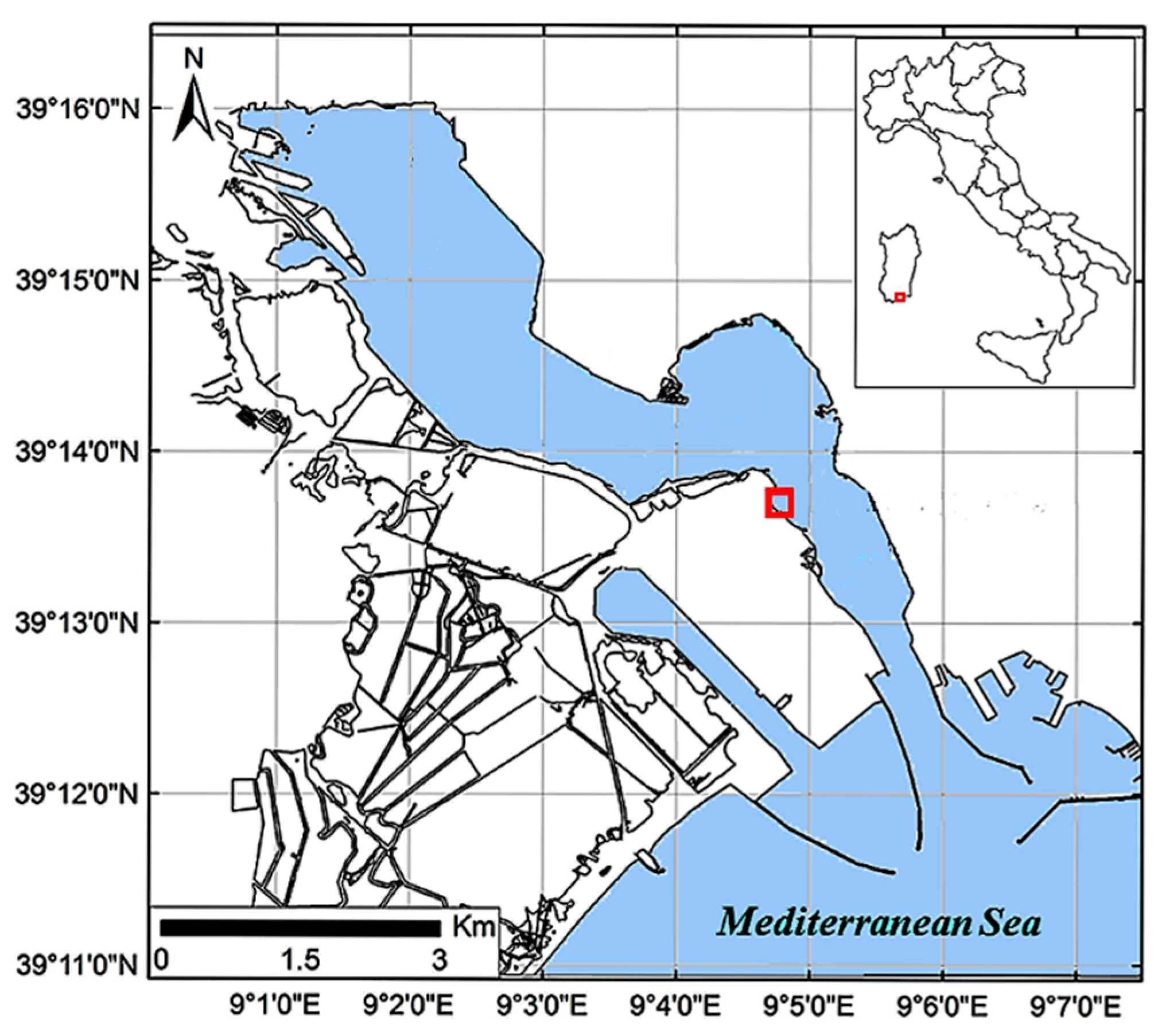
2.1. Chemical and Physical Parameters of the Monitored Site
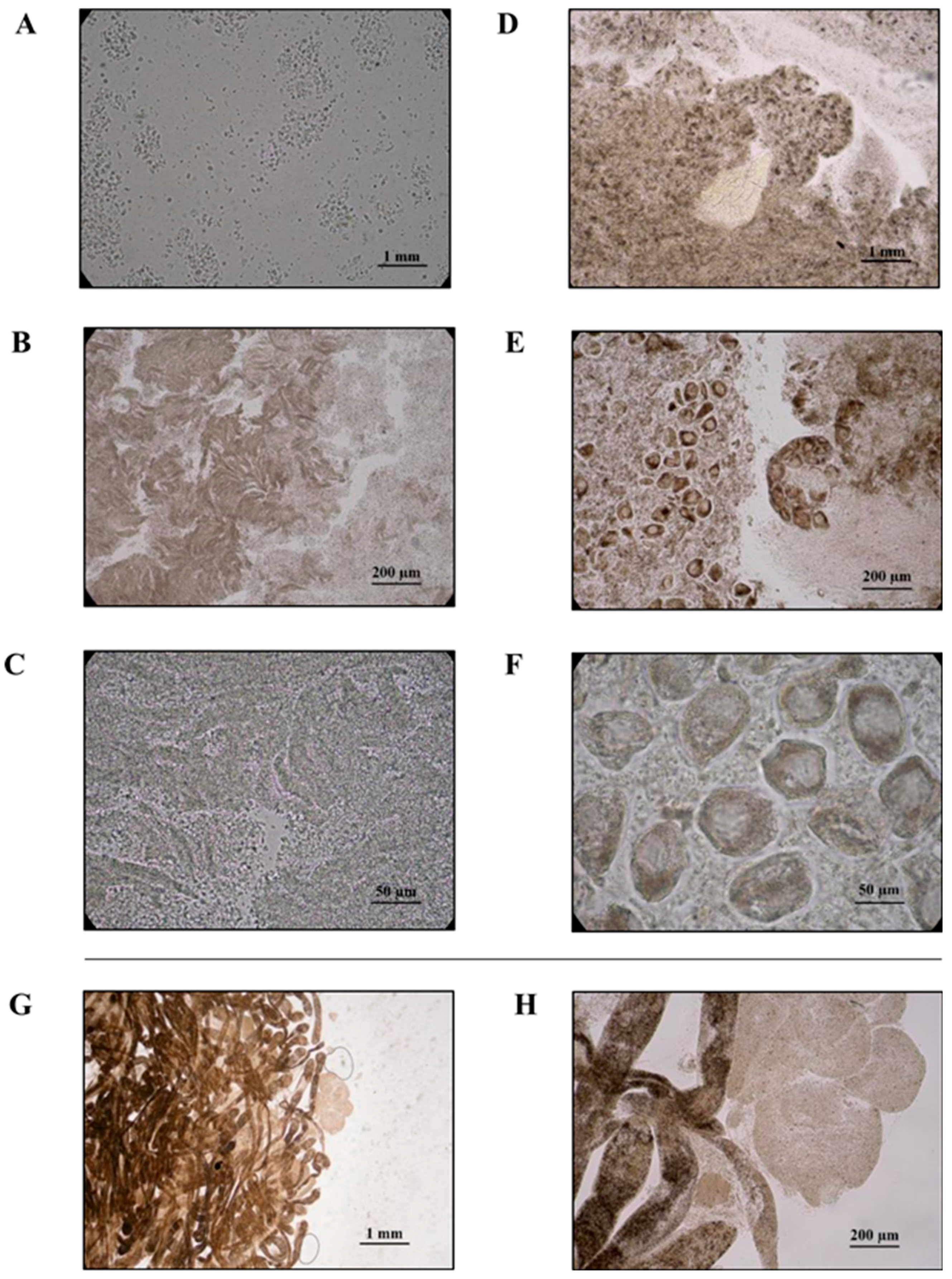
2.2. Sex and Parasitosis Determination
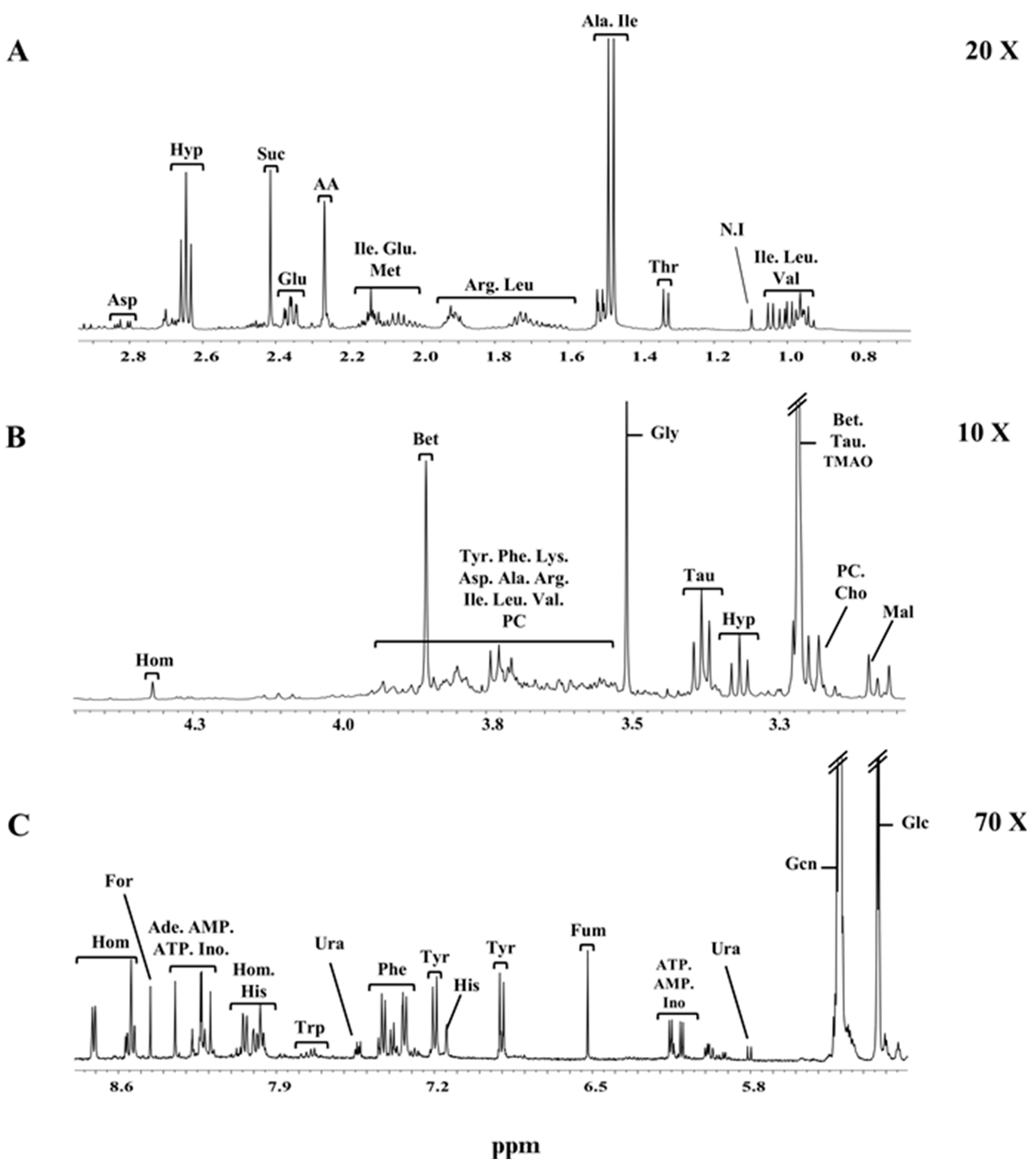
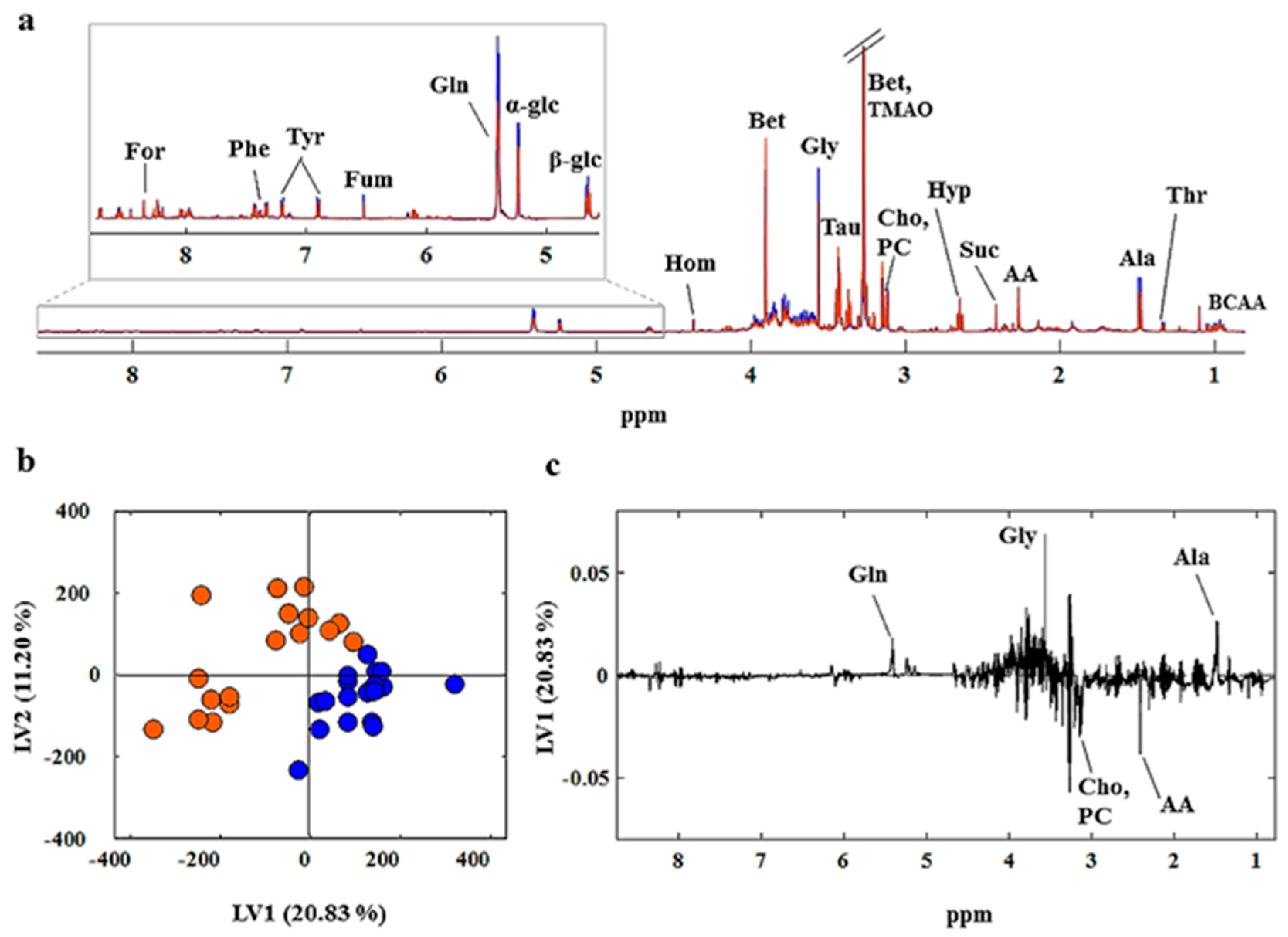
2.3. NMR Spectra of Clams
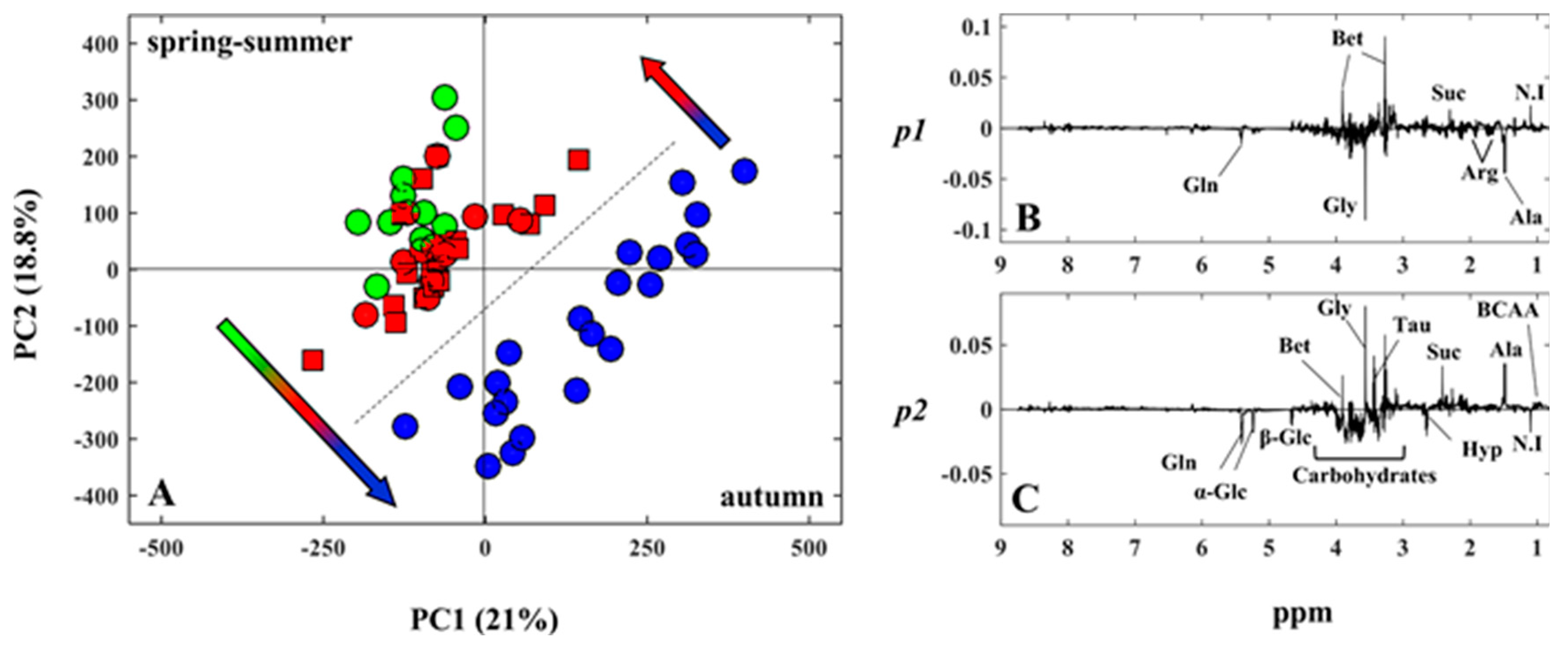
2.4. Chemometric Analysis
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Sample Collection
4.3. Water Chemical-Physical Parameters
4.4. Sex and Parasitosis Determination
4.5. NMR Analysis
Metabolite Extraction
4.6. NMR Measurements
4.7. Preprocessing of the NMR Spectral Data
4.8. Multivariate Analysis of the NMR Spectral Data
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dettmer, K.; Hammock, B.D. Metabolomics—A new exciting field within the “omics” Sciences. Environ. Health Perspect. 2004, 112, 396–397. [Google Scholar]
- Putri, S.P.; Nakayama, Y.; Matsuda, F.; Uchikata, T.; Kobayashi, S.; Matsubara, A.; Fukusaki, E. Current metabolomics: Practical applications. J. Biosci. Bioeng. 2013, 115, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Lankadurai, B.P.; Nagato, E.G.; Simpson, M.J. Environmental metabolomics: An emerging approach to study organisms responses to environmental stressors. Environ. Rev. 2013, 21, 180–205. [Google Scholar] [CrossRef]
- Viant, M.; Rosenblum, E.S.; Tjeerdema, R.S. NMR-based metabolomics: A powerful approach for characterizing the effects of environmental stressors on organism health. Environ. Sci. Technol. 2003, 37, 4982–4989. [Google Scholar] [CrossRef] [PubMed]
- Veldhoen, N.; Ikonomou, M.G.; Helbing, C.C. Molecular profiling of marine fauna: Integration of omics with environmental assessment of the world’s oceans. Ecotoxicol. Environ. Saf. 2012, 76, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Viant, M.R. Recent developments in environmental metabolomics. Mol. Biosyst. 2008, 4, 980. [Google Scholar] [CrossRef] [PubMed]
- Cappello, T.; Maisano, M.; Mauceri, A.; Fasulo, S. Contamination in posterior adductor muscles of caged mussel Mytilus. Ecotoxicol. Environ. Saf. 2017, 142, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Cappello, T.; Brandão, F.; Ana, M.; Maisano, M.; Mauceri, A.; Canário, J.; Pacheco, M.; Pereira, P. Science of the total environment insights into the mechanisms underlying mercury-induced oxidative stress in gills of wild fish (Liza aurata) combining 1H NMR metabolomics and conventional biochemical assays. Sci. Total Environ. 2016, 549, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Gosling, E. Bivalve Culture. In Bivalve Molluscs: Biol., Ecology and Culture; Blackwell Press: England, UK, 2003; ISBN 9780470995532. [Google Scholar]
- Alfaro, A.C.; Young, T. Showcasing metabolomic applications in aquaculture: A review. Rev. Aquac. 2016, 1–18. [Google Scholar] [CrossRef]
- Young, T.; Alfaro, A.C. Metabolomic strategies for aquaculture research: A primer. Rev. Aquac. 2016. [Google Scholar] [CrossRef]
- Karakoltsidis, P.A.; Zotos, A.; Constantinides, S.M. Composition of the commercially important mediterranean finfish, crustaceans, and molluscs. J. Food Compos. Anal. 1995, 8, 258–273. [Google Scholar] [CrossRef]
- Orban, E.; Di Lena, G.; Nevigato, T.; Casini, I.; Marzetti, A.; Caproni, R. Seasonal changes in meat content, condition index and chemical composition of mussels (Mytilus galloprovincialis) cultured in two different Italian sites. Food Chem. 2002, 77, 57–65. [Google Scholar] [CrossRef]
- Da Silva, J.G. The State of World Fisheries and Aquaculture 2016; Food and Agriculture Organization of the United Nations Press: Rome, Italy, 2016; pp. 1–204. ISBN 9789251091852. [Google Scholar]
- Ojea, J.; Pazos, A.J.; Martínez, D.; Novoa, S.; Sánchez, J.L.; Abad, M. Seasonal variation in weight and biochemical composition of the tissues of Ruditapes decussatus in relation to the gametogenic cycle. Aquaculture 2004, 238, 451–468. [Google Scholar] [CrossRef]
- Wold, S.; Esbensen, K.; Geladi, P. Principal component analysis. Chemom. Intell. Lab. Syst. 1987, 2, 37–52. [Google Scholar] [CrossRef]
- Barker, M.; Rayens, W. Partial least squares for discrimination. J. Chemom. 2003, 17, 166–173. [Google Scholar] [CrossRef]
- Culurgioni, J.; Figus, V.; Cabiddu, S.; De Murtas, R.; Cau, A.; Sabatini, A. Larval helminth parasites of fishes and shellfishes from Santa Gilla lagoon (sardinia, western mediterranean), and their use as bioecological indicators. Estuar. Coasts 2015, 38, 1505–1519. [Google Scholar] [CrossRef]
- Culurgioni, J.; Sabatini, A.; De Murtas, R.; Mattiucci, S.; Figus, V. Helminth parasites of fish and shellfish from the Santa Gilla Lagoon in southern Sardinia, Italy. J. Helminthol. 2014, 88, 489–498. [Google Scholar] [CrossRef] [PubMed]
- De Montaudouin, X.; Bazairi, H.; Mlik, K.; Gonzalez, P. Bacciger bacciger (Trematoda: Fellodistomidae) infection effects on wedge clam Donax trunculus condition. Dis. Aquat. Org. 2014, 111, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.H.; Claiborne, J.B.; Currie, S. The Physiology of Fishes, 4th ed.; CRC Press: Cleveland, OH, USA, 2006; pp. 177–230. ISBN 9780849320224. [Google Scholar]
- Gosling, E. Marine Bivalve Molluscs, 2nd ed.; Wiley Blackwell Press: New York, NY, USA; Hoboken, NJ, USA, 2015; pp. 1–513. [Google Scholar] [CrossRef]
- Yancey, P.H. Organic osmolytes as compatible, metabolic and counteracting cytoprotectants in high osmolarity and other stresses. J. Exp. Biol. 2005, 208, 2819–2830. [Google Scholar] [CrossRef] [PubMed]
- Pérez Camacho, A.; Delgado, M.; Fernández-Reiriz, M.; Labarta, U. Energy balance, gonad development and biochemical composition in the clam Ruditapes decussatus. Mar. Ecol. Prog. Ser. 2003, 258, 133–145. [Google Scholar] [CrossRef]
- Ellis, R.P.; Spicer, J.I.; Byrne, J.J.; Sommer, U.; Viant, M.R.; White, D.A.; Widdicombe, S. 1H NMR metabolomics reveals contrasting response by male and female mussels exposed to reduced seawater pH, increased temperature, and a pathogen. Environ. Sci. Technol. 2014, 48, 7044–7052. [Google Scholar] [CrossRef] [PubMed]
- Kasschau, R. Glycine concentration as a biochemical indicator of sex and maturation in the sea anemone Comp. Biochem. Physiol. A Physiol. 1982, 72, 595–597. [Google Scholar] [CrossRef]
- Cubero-Leon, E.; Minier, C.; Rotchell, J.M.; Hill, E.M. Metabolomic analysis of sex specific metabolites in gonads of the mussel, mytilus edulis. Comp. Biochem. Physiol. 2012, 7, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Stanley, S.G. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Savorani, F.; Tomasi, G.; Engelsen, S.B. Icoshift: A versatile tool for the rapid alignment of 1D NMR spectra. J. Magn. Reson. 2010, 202, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.; Cloarec, O.; Holmes, E.; Nicholson, J.K.; Lindon, J.C. Scaling and normalization effects in NMR spectroscopic metabonomic data sets. Anal. Chem. 2006, 78, 2262–2267. [Google Scholar] [CrossRef] [PubMed]
- Worley, B.; Powers, R. Multivariate analysis in metabolomics. Curr. Metabolomics 2012, 1, 92–107. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | ppm | Multiplicity * |
|---|---|---|
| α-Glucose (α-glc) | 5.23 | d |
| β-Glucose (β-glc) | 4.66 | d |
| Acetoacetate (AA) | 2.27 | s |
| Adenine (Ade) | 8.17/8.20 | s/s |
| Alanine (Ala) | 1.49 | d |
| Arginine (Arg) | 1.63/1.71/1.92 | m/m/m |
| Aspartate (Asp) | 2.66/2.80 | dd/dd |
| Adenosine Monophosphate (AMP) | 6.13/8.26/8.56 | d/s/s |
| Adenosine Triphosphate (ATP) | 6.13/8.25/8.51 | d/s/s |
| Betaine (Bet) | 3.26/3.89 | s/s |
| Choline (Cho) | 3.19 | s |
| Formate (Form) | 8.44 | s |
| Fumarate (Fum) | 6.5 | s |
| Glutamate (Glu) | 2.36 | t |
| Glycine (Gly) | 3.54 | s |
| Glycogen (Gcn) | 5.39 | bb |
| Histidine (His) | 7.13/7.98 | s/s |
| Homarine (Hom) | 4.35 | s |
| Hypotaurine (Hyp) | 2.64/3.34 | t/t |
| Inosine (Ino) | 6.09/8.22/8.33 | d/s/s |
| Isoleucine (Ile) | 0.93/0.99 | d/t |
| Leucine (Leu) | 0.96 | d |
| Malonate (Mal) | 3.11 | s |
| Methionine (Met) | 2.12 | s |
| N.I. | 1.08 | s |
| Phenylalanine (Phe) | 7.36/7.41/7.42 | m/m/m |
| Phosphoryl Choline (PC) | 3.22 | s |
| Succinate (Suc) | 2.4 | s |
| Taurine (Tau) | 3.25/3.41 | t/t |
| Trimethylamine oxide (TMAO) | 3.26 | s |
| Threonine (Thr) | 1.32 | d |
| Tryptophan (Trp) | 7.72 | d |
| Tyrosine (Tyr) | 6.89/7.18 | d/d |
| Uracil (Ura) | 5.79/7.52 | d/d |
| Valine (Val) | 0.98/1.03 | d/d |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aru, V.; Balling Engelsen, S.; Savorani, F.; Culurgioni, J.; Sarais, G.; Atzori, G.; Cabiddu, S.; Cesare Marincola, F. The Effect of Season on the Metabolic Profile of the European Clam Ruditapes decussatus as Studied by 1H-NMR Spectroscopy. Metabolites 2017, 7, 36. https://doi.org/10.3390/metabo7030036
Aru V, Balling Engelsen S, Savorani F, Culurgioni J, Sarais G, Atzori G, Cabiddu S, Cesare Marincola F. The Effect of Season on the Metabolic Profile of the European Clam Ruditapes decussatus as Studied by 1H-NMR Spectroscopy. Metabolites. 2017; 7(3):36. https://doi.org/10.3390/metabo7030036
Chicago/Turabian StyleAru, Violetta, Søren Balling Engelsen, Francesco Savorani, Jacopo Culurgioni, Giorgia Sarais, Giulia Atzori, Serenella Cabiddu, and Flaminia Cesare Marincola. 2017. "The Effect of Season on the Metabolic Profile of the European Clam Ruditapes decussatus as Studied by 1H-NMR Spectroscopy" Metabolites 7, no. 3: 36. https://doi.org/10.3390/metabo7030036
APA StyleAru, V., Balling Engelsen, S., Savorani, F., Culurgioni, J., Sarais, G., Atzori, G., Cabiddu, S., & Cesare Marincola, F. (2017). The Effect of Season on the Metabolic Profile of the European Clam Ruditapes decussatus as Studied by 1H-NMR Spectroscopy. Metabolites, 7(3), 36. https://doi.org/10.3390/metabo7030036