Impact of Soil Warming on the Plant Metabolome of Icelandic Grasslands



 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Material and Methods
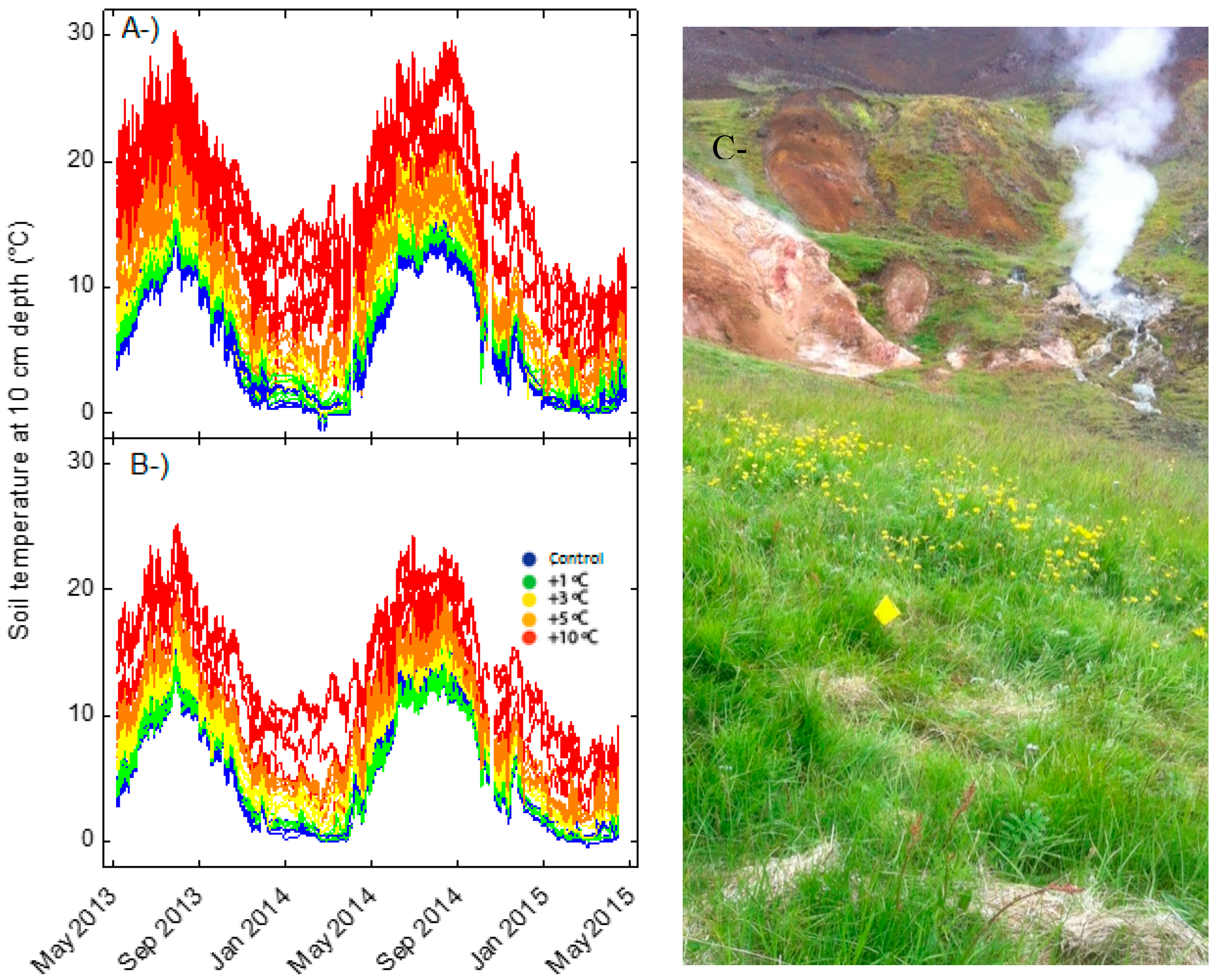
2.1. Description of the Study Area
Study Sites
2.2. Experimental Design
2.3. Collection and Preparation of Tissue Samples
2.4. Extraction of Metabolites for Analysis by Liquid Chromatography-Mass Spectrometry (LC-MS)
2.5. LC-MS Analysis
2.6. Processing of LC-MS Data
2.7. RNA:DNA Ratio
2.8. Statistical Analyses
3. Results
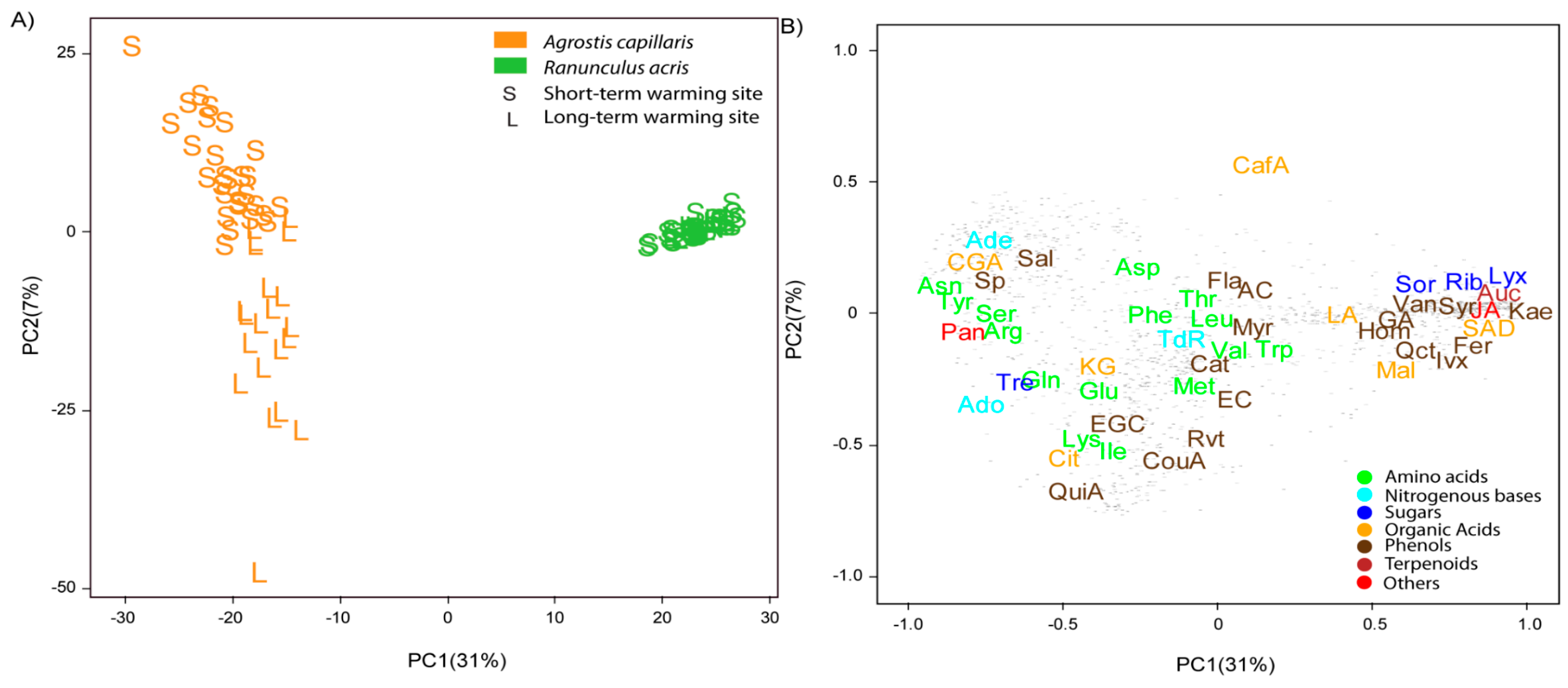
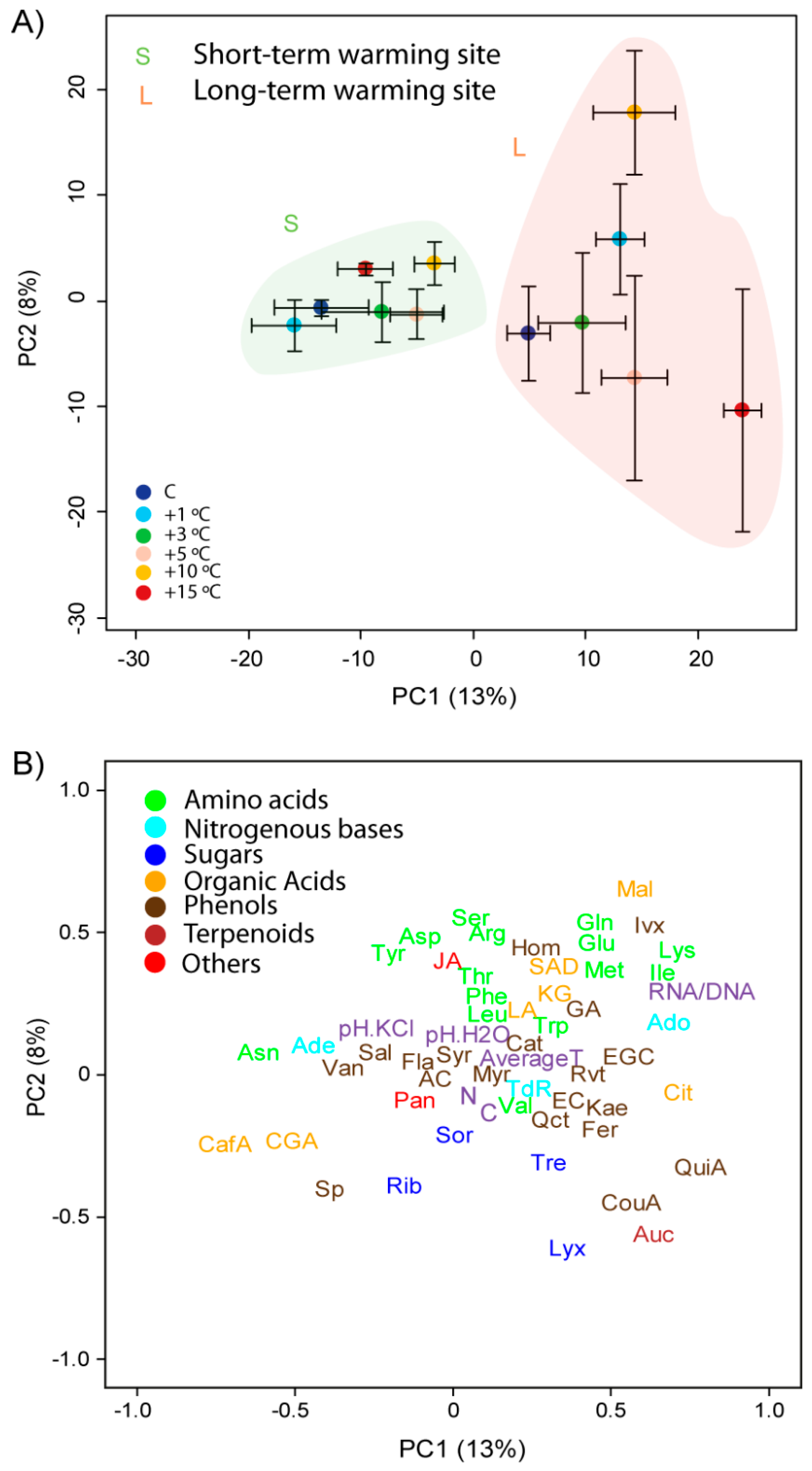
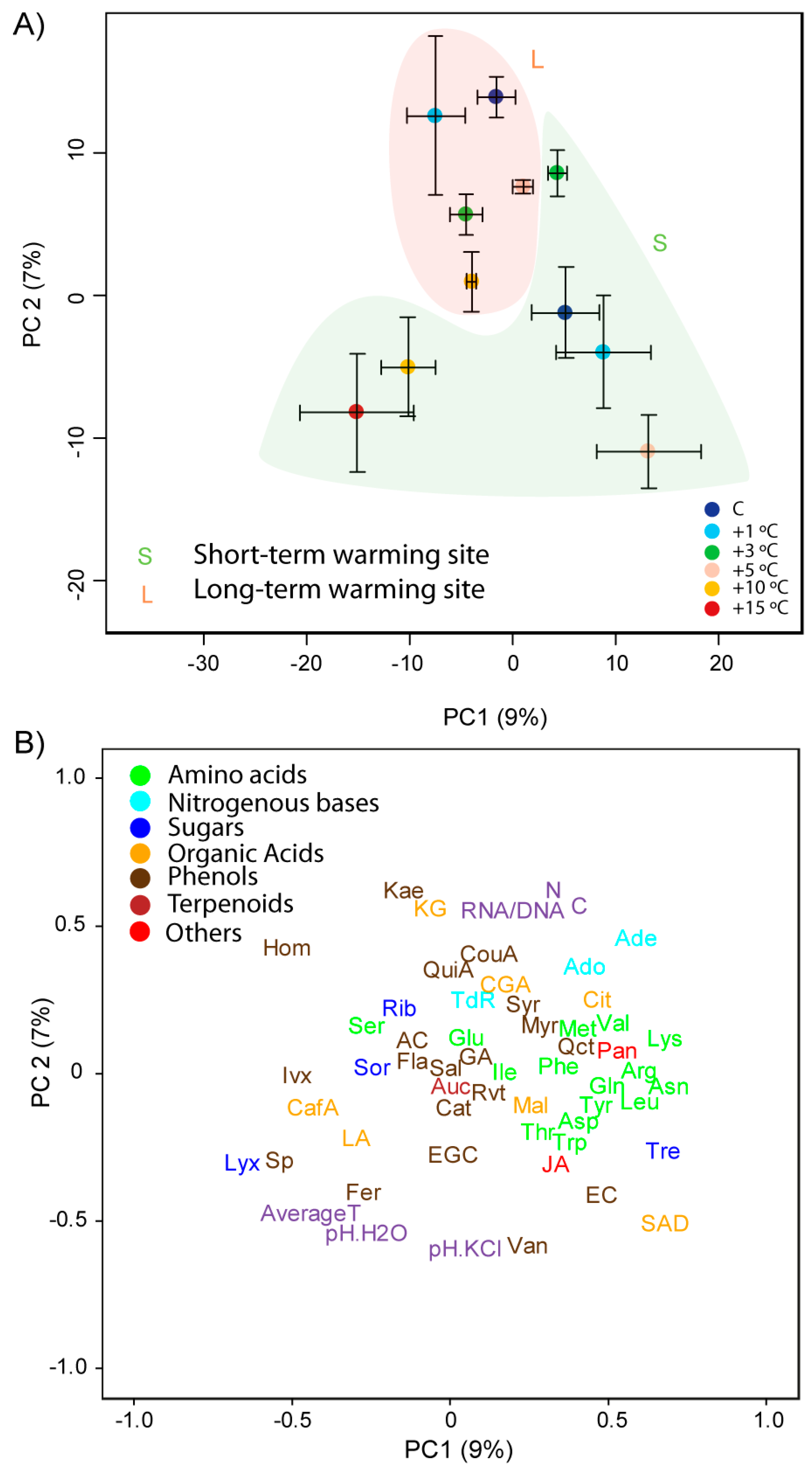
3.1. Differences between Species, Sites and Warming Levels
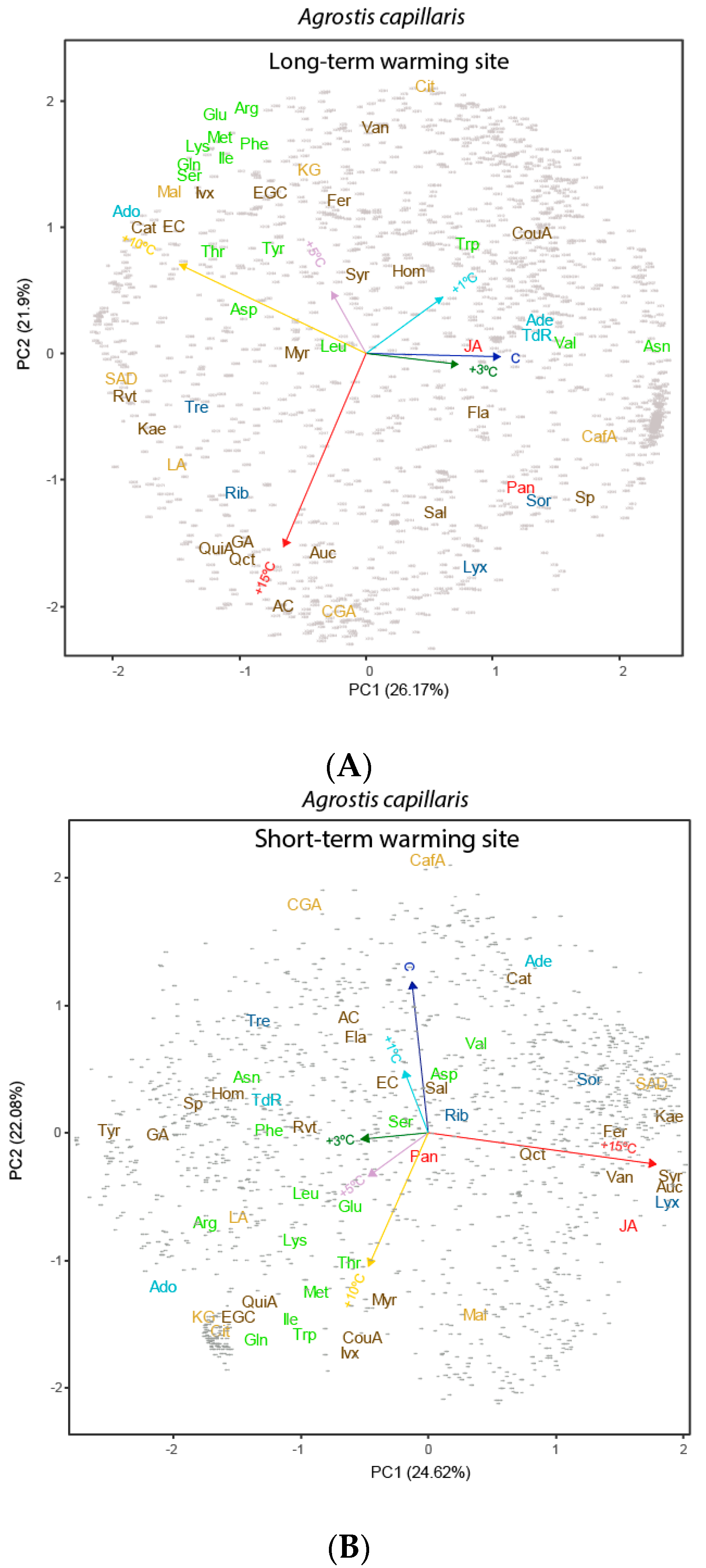
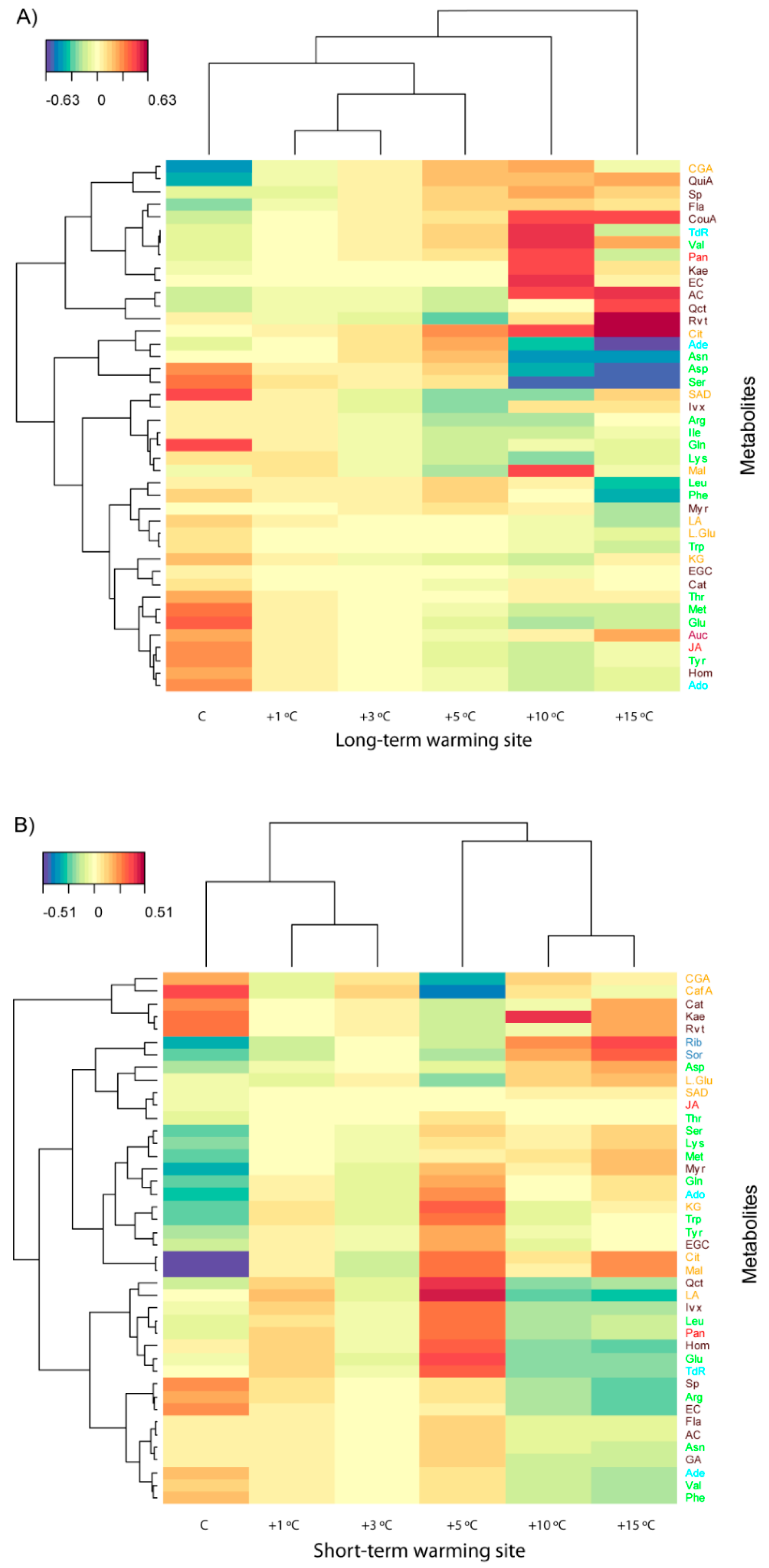
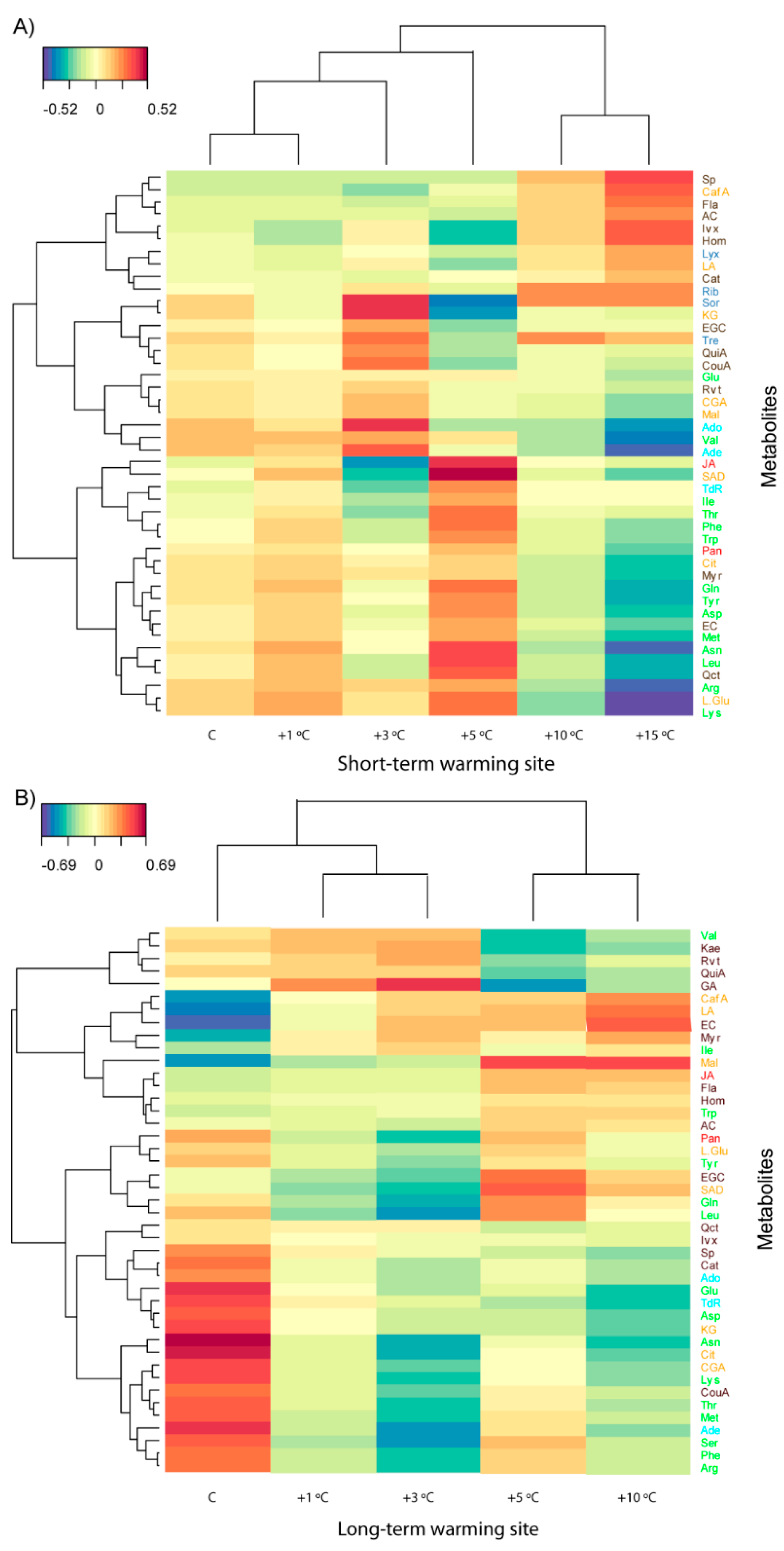
3.2. Effects of Length and Level of Warming on the Metabolome of A. capillaris
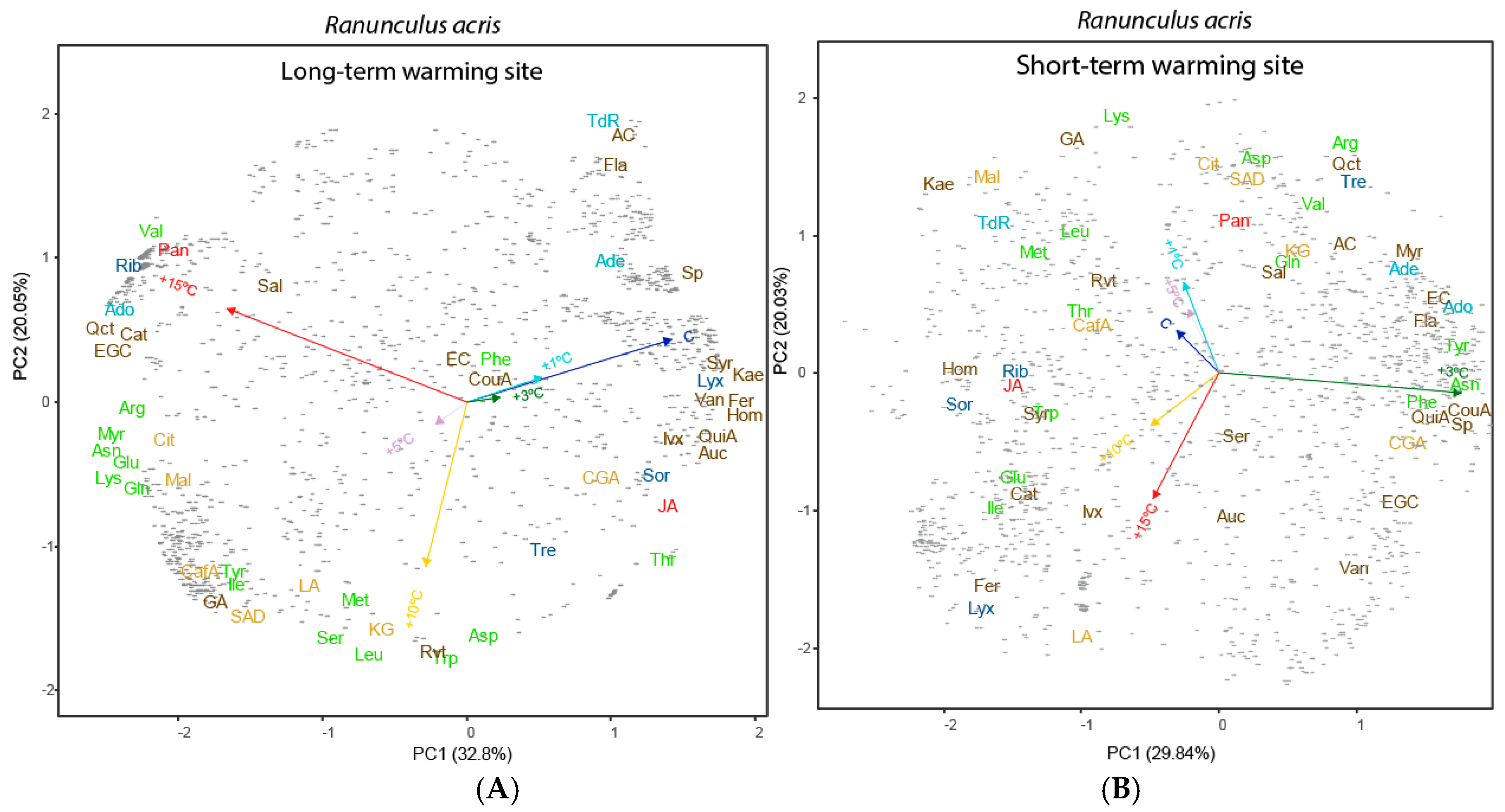
3.3. Effects of Length and Level of Warming on the Metabolome of R. acris
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Climate Change 2013: The Physical Science Basis. 2013. Available online: http://www.ipcc.ch/report/ar5/wg1/ (accessed on 19 August 2017).
- Moss, R.; Babiker, M.; Brinkman, S.; Calvo, E.; Carter, T.; Edmonds, J.; Elgizouli, I.; Emori, S.; Erda, L.; Hibbard, K.; et al. Towards new scenarios for analysis of emissions, climate change, impacts and response strategies. In The Netherlands: Technical Summary; Intergovernmental Panel on Climate Change Secretariat (IPCC): Geneva, Switzerland, 2008; ISBN 9789291691241. Available online: vuir.vu.edu.au/4819 (accessed on 19 August 2017).
- Parmesan, C. Ecological and Evolutionary Responses to Recent Climate Change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Estiarte, M.; Ogaya, R.; Carnicer, J.; Coll, M.; Barbeta, A.; Rivas-Ubach, A.; Llusià, J.; Garbulsky, M.; et al. Evidence of current impact of climate change on life: A walk from genes to the biosphere. Glob. Chang. Biol. 2013, 19, 2303–2338. [Google Scholar] [CrossRef] [PubMed]
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesank, C.; Beebee, T.J.C.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.; Tjoelker, M.; Vanderklein, D.; Buschena, C. Photosynthesis and respiration rates depend on leaf and root morphology and nitrogen concentration in nine boreal tree species differing in relative growth rate. Funct. Ecol. 1998, 12, 395–405. [Google Scholar] [CrossRef]
- Reich, P.B.; Wright, I.J.; Lusk, C.H. Predicting leaf physiology from simple plant and climate attributes: A global GLOPNET analysis. Ecol. Appl. 2007, 17, 1982–1988. [Google Scholar] [CrossRef] [PubMed]
- Carnicer, J.; Coll, M.; Ninyerola, M.; Pons, X.; Sánchez, G.; Peñuelas, J. Widespread crown condition decline, food web disruption, and amplified tree mortality with increased climate change-type drought. Proc. Natl. Acad. Sci. USA 2011, 108, 1474–1478. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Martínez, M.; Vicca, S.; Janssens, I.A.; Sardans, J.; Luyssaert, S.; Campioli, M.; Chapin, F.S., III; Ciais, P.; Malhi, Y.; Obersteiner, M.; et al. Nutrient availability as the key regulator of global forest carbon balance. Nat. Clim. Chang. 2014, 4, 471–476. [Google Scholar] [CrossRef]
- Peñuelas, J.; Poulter, B.; Sardans, J.; Ciais, P.; van der Velde, M.; Bopp, L.; Boucher, O.; Godderis, Y.; Hinsinger, P.; Llusia, J.; et al. Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe. Nat. Commun. 2013, 4, 2934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Euskirchen, E.S.; Mcguire, A.D.; Chapin, F.S.; Yi, S.; Thompson, C.C.; Thompson, C.C. Changes in Vegetation in Northern Alaska under Scenarios of Climate Change, 2003–2100: Implications for Climate Feedbacks Changes in vegetation in northern Alaska under scenarios of climate change, 2003–2100: Implications for climate feedbacks. Ecol. Appl. 2009, 19, 1022–1043. [Google Scholar] [CrossRef] [PubMed]
- Mack, M.C.; Schuur, E.A.; Bret-Harte, M.S.; Shaver, G.R.; Chapin, F.S., III. Ecosystem carbon storage in arctic tundra reduced by long-term nutrient fertilization. Nature 2004, 431, 440–443. [Google Scholar] [CrossRef] [PubMed]
- Hobbie, S.E.; Chapin, F.S. The Response of Tundra Plant Biomass, Aboveground Production, Nitrogen, and CO2 Flux to Experimental Warming. Ecology 1998, 79, 1526–1544. [Google Scholar]
- Hobbie, S.E.; Chapin, F.S. Winter regulation of tundra litter carbon and nitrogen dynamics. Biogeochemistry 1996, 35, 327–338. [Google Scholar] [CrossRef]
- Walker, M.D.; Wahren, C.H.; Hollister, R.D.; Henry, G.H.R.; Ahlquist, L.E.; Alatalo, J.M.; Bret-harte, M.S.; Calef, M.P.; Callaghan, T.V.; Carroll, A.B.; et al. Plant community responses to experimental warming across the tundra biome. Proc. Natl. Acad. Sci. USA 2006, 103, 1342–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmendorf, S.C.; Henry, G.H.R.; Hollister, R.D.; Björk, R.G.; Boulanger-Lapointe, N.; Cooper, E.J.; Cornelissen, J.H.C.; Day, T.A.; Dorrepaal, E.; Elumeeva, T.G.; et al. Plot-scale evidence of tundra vegetation change and links to recent summer warming. Nat. Clim. Chang. 2012, 2, 453–457. [Google Scholar] [CrossRef]
- Schär, C.; Jendritzky, G. Climate change: Hot news from summer 2003. Nature 2004, 432, 559–560. [Google Scholar] [CrossRef] [PubMed]
- Gutschick, V.P.; BassiriRad, H. Extreme events as shaping physiology, ecology, and evolution of plants: Toward a unified definition and evaluation of their consequences. New Phytol. 2003, 160, 21–42. [Google Scholar] [CrossRef]
- Reusch, T.B.H.; Ehlers, A.; Hämmerli, A.; Worm, B. Ecosystem recovery after climatic extremes enhanced by genotypic diversity. Proc. Natl. Acad. Sci. USA 2005, 102, 2826–2831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jentsch, A.; Beierkuhnlein, C. Research frontiers in climate change: Effects of extreme meteorological events on ecosystems. C. R. Geosci. 2008, 340, 621–628. [Google Scholar] [CrossRef]
- Knapp, A.K.; Beier, C.; Briske, D.D.; Classen, A.T.; Luo, Y.; Reichstein, M.; Smith, M.D.; Smith, S.D.; Bell, J.E.; Fay, P.A.; et al. Consequences of More Extreme Precipitation Regimes for Terrestrial Ecosystems. Bioscience 2008, 58, 811–821. [Google Scholar] [CrossRef]
- Jentsch, A.; Kreyling, J.; Elmer, M.; Gellesch, E.; Glaser, B.; Grant, K.; Hein, R.; Lara, M.; Mirzae, H.; Nadler, S.E.; et al. Climate extremes initiate ecosystem-regulating functions while maintaining productivity. J. Ecol. 2011, 99, 689–702. [Google Scholar] [CrossRef]
- Smith, M.D. The ecological role of climate extremes: Current understanding and future prospects. J. Ecol. 2011, 99, 651–655. [Google Scholar] [CrossRef]
- Fiehn, O.; Kopka, J.; Dörmann, P.; Altmann, T.; Trethewey, R.N.; Willmitzer, L. Metabolite profiling for plant functional genomics. Nat. Biotechnol. 2000, 18, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Sardans, J. Ecological metabolomics. Chem. Ecol. 2009, 25, 305–309. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J.; Rivas-Ubach, A. Ecological metabolomics: Overview of current developments and future challenges. Chemoecology 2011, 21, 191–225. [Google Scholar] [CrossRef]
- Charlton, A.J.; Donarski, J.A.; Harrison, M.; Jones, S.A.; Godward, J.; Oehlschlager, S.; Arques, J.L.; Ambrose, M.; Chinoy, C.; Mullineaux, P.M.; et al. Responses of the pea (Pisum sativum L.) leaf metabolome to drought stress assessed by nuclear magnetic resonance spectroscopy. Metabolomics 2008, 4, 312–327. [Google Scholar] [CrossRef]
- Fumagalli, E.; Baldoni, E.; Abbruscato, P.; Piffanelli, P.; Genga, A.; Lamanna, R.; Consonni, R. NMR Techniques Coupled with Multivariate Statistical Analysis: Tools to Analyse Oryza sativa Metabolic Content under Stress Conditions. J. Agron. Crop Sci. 2009, 195, 77–88. [Google Scholar] [CrossRef]
- Lugan, R.; Niogret, M.-F.; Kervazo, L.; Larher, F.R.; Kopka, J.; Bouchereau, A. Metabolome and water status phenotyping of Arabidopsis under abiotic stress cues reveals new insight into ESK1 function. Plant Cell Environ. 2009, 32, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Michaud, R.M.; Benoit, J.B.; Lopez-Martinez, G.; Elnitsky, M.A; Lee, R.E.; Denlinger, D.L. Metabolomics reveals unique and shared metabolic changes in response to heat shock, freezing and desiccation in the Antarctic midge, Belgica antarctica. J. Insect Physiol. 2008, 54, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, C.; Passarinho, J.A.; Ricardo, C.P. Effect of drought and rewatering on the metabolism of Lupinus albus organs. J. Plant Physiol. 2004, 161, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Michaud, M.R.; Denlinger, D.L. Shifts in the carbohydrate, polyol, and amino acid pools during rapid cold-hardening and diapause-associated cold-hardening in flesh flies (Sarcophaga crassipalpis): A metabolomic comparison. J. Comp. Physiol. B 2007, 177, 753–763. [Google Scholar] [CrossRef] [PubMed]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Oravec, M.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; Parella, T.; Peñuelas, J. Warming differentially influences the effects of drought on stoichiometry and metabolomics in shoots and roots. New Phytol. 2015, 207, 591–603. [Google Scholar] [CrossRef] [PubMed]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Rivas-Ubach, A.; Oravec, M.; Vecerova, K.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; et al. Opposite metabolic responses of shoots and roots to drought. Sci. Rep. 2014, 4, 6829. [Google Scholar] [CrossRef] [PubMed]
- Allakhverdiev, S.I.; Nishiyama, Y.; Suzuki, I.; Tasaka, Y.; Murata, N. Genetic engineering of the unsaturation of fatty acids in membrane lipids alters the tolerance of Synechocystis to salt stress. Proc. Natl. Acad. Sci. USA 1999, 96, 5862–5867. [Google Scholar] [CrossRef] [PubMed]
- Horváth, I.; Vigh, L.; Pali, T.; Thompson, G.A. Effect of catalytic hydrogenation of Tetrahymena ciliary phospholipid fatty acids on ciliary phospholipase A activity. Biochim. Biophys. Acta Lipids Lipid Metab. 1989, 1002, 409–412. [Google Scholar] [CrossRef]
- Vigh, L.; Gombos, Z.; Horváth, I.; Joó, F. Saturation of membrane lipids by hydrogenation induces thermal stability in chloroplast inhibiting the heat-dependent stimulation of Photosystem I-mediated electron transport. Biochim. Biophys. Acta Biomembr. 1989, 979, 361–364. [Google Scholar] [CrossRef]
- Vigh, L.; Los, D.A; Horváth, I.; Murata, N. The primary signal in the biological perception of temperature: Pd-catalyzed hydrogenation of membrane lipids stimulated the expression of the desA gene in Synechocystis PCC6803. Proc. Natl. Acad. Sci. USA 1993, 90, 9090–9094. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, F.; Kopka, J.; Haskell, D.W.; Zhao, W.; Schiller, K.C.; Gatzke, N.; Sung, D.Y.; Guy, C.L. Exploring the Temperature-Stress Metabolome. Plant Physiol. 2004, 136, 4159–4168. [Google Scholar] [CrossRef] [PubMed]
- Malmendal, A.; Overgaard, J.; Bundy, J.G.; Sørensen, J.G.; Nielsen, N.C.; Loeschcke, V.; Holmstrup, M. Metabolomic profiling of heat stress: Hardening and recovery of homeostasis in Drosophila. Am. J. Physiol. 2006, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Klimov, V.V.; Los, D.A.; Carpentier, R.; Mohanty, P. Heat stress: An overview of molecular responses in photosynthesis. Photosynth. Res. 2008, 98, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Blamey, M.; Fitter, S.R.; Alastair, F. Wild Flowers of Britain & Ireland; A. & C. Black: London, UK, 2003; Available online: http://agris.fao.org/agris-search/search.do?recordID=US201300085632? (accessed on 21 August 2017).
- Nicolini, G. Enciclopedia Botanica Motta. Volume Primo Milano Federico Motta Editore 1960, 198. [Google Scholar]
- Arnalds, Ó. Soil Survey and Databases in Iceland. Eur. Soil Bur. Res. Rep. 1999, 6, 91–96. [Google Scholar]
- Arnalds, O. Volcanic soils of Iceland. Catena 2004, 56, 3–20. [Google Scholar] [CrossRef]
- Geptner, A.R.; Ivanovskaya, T.A.; Pokrovskaya, E.V. Hydrothermally altered clayey sediments in the rift zone of Iceland (influence of microbiota on accumulation of minor elements). Lithol. Miner. Resour. 2006, 41, 332–343. [Google Scholar] [CrossRef]
- Zakharova, O.K.; Spichak, V.V. Geothermal fields of Hengill Volcano, Iceland. J. Volcanol. Seismol. 2012, 6, 1–14. [Google Scholar] [CrossRef]
- Magnússon, A. Jarðabók. Annað Bindi; 1708. [Google Scholar]
- Halldórsson, B.; Sigbjörnsson, R. The Mw 6.3 Ölfus earthquake at 15:45 UTC on 29 May 2008 in South Iceland: ICEARRAY strong-motion recordings. Soil Dyn. Earthq. Eng. 2009, 29, 1073–1083. [Google Scholar] [CrossRef]
- Sigurdsson, B.D.; Leblans, N.I.W.; Dauwe, S.; Guðmundsdóttir, E.; Gundersen, P.; Gunnarsdóttir, G.E.; Holmstrup, M.; Ilieva-Makulec, K.; Kätterer, T.; Marteinsdóttir, B.; et al. Geothermal ecosystems as natural climate change experiments: The ForHot research site in Iceland as a case study. Icelandic Agric. Sci. 2016, 29, 53–71. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Oresic, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Gorokhova, E.; Kyle, M. Analysis of nucleic acids in Daphnia: Development of methods and ontogenetic variations in RNA-DNA content. J. Plankton Res. 2002, 24, 511–522. [Google Scholar] [CrossRef]
- Wagner, M.; Durbin, E.; Buckley, L. RNA:DNA ratios as indicators of nutritional condition in the copepod Calanus finmarchicus. Mar. Ecol. Prog. Ser. 1998, 162, 173–181. [Google Scholar] [CrossRef]
- Bentle, L.A.; Dutta, S.; Metcoff, J. The sequential enzymatic determination of DNA and RNA. Anal. Biochem. 1981, 116, 5–16. [Google Scholar] [CrossRef]
- Bro, R.; Smilde, A.K. Centering and scaling in component analysis. J. Chemom. 2003, 17, 16–33. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRI-MER: Guide to Software and Statistical Methods; Primer-E.: Plymouth, UK, 2008. [Google Scholar]
- Smilde, A.K.; Jansen, J.J.; Hoefsloot, H.C.J.; Lamers, R.J.A.N.; van der Greef, J.; Timmerman, M.E. ANOVA-simultaneous component analysis (ASCA): A new tool for analyzing designed metabolomics data. Bioinformatics 2005, 21, 3043–3048. [Google Scholar] [CrossRef] [PubMed]
- Urbina, I.; Sardans, J.; Beierkuhnlein, C.; Jentsch, A.; Backhaus, S.; Grant, K.; Kreyling, J.; Peñuelas, J. Shifts in the elemental composition of plants during a very severe drought. Environ. Exp. Bot. 2015, 111, 63–73. [Google Scholar] [CrossRef]
- Sardans, J.; Bartrons, M.; Margalef, O.; Gargallo-Garriga, A.; Janssens, I.A.; Ciais, P.; Obersteiner, M.; Sigurdsson, B.D.; Chen, H.Y.H.; Penuelas, J. Plant invasion is associated with higher plant-soil nutrient concentrations in nutrient poor-environments. Glob. Chang. Biol. 2016, 23, 1282–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamant, S.; Eliahu, N.; Rosenthal, D.; Goloubinoff, P. Chemical Chaperones Regulate Molecular Chaperones in Vitro and in Cells under Combined Salt and Heat Stresses. J. Biol. Chem. 2001, 276, 39586–39591. [Google Scholar] [CrossRef] [PubMed]
- Lipka, V.; Dittgen, J.; Bednarek, P.; Bhat, R.; Wiermer, M.; Stein, M.; Landtag, J.; Brandt, W.; Rosahl, S.; Scheel, D.; et al. Pre- and postinvasion defenses both contribute to nonhost resistance in Arabidopsis. Science 2005, 310, 1180–1183. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive Oxygen Species: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When Defense Pathways Collide. The Response of Arabidopsis to a Combination of Drought and Heat Stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, H.; Hakata, M. Atlas of rice grain filling-related metabolism under high temperature: Joint analysis of metabolome and transcriptome demonstrated inhibition of starch accumulation and induction of amino acid accumulation. Plant Cell Physiol. 2010, 51, 795–809. [Google Scholar] [CrossRef] [PubMed]
- Lea, P.J.; Sodek, L.; Parry, M.A.J.; Shewry, P.R.; Halford, N.G. Asparagine in plants. Ann. Appl. Biol. 2007, 150, 1–26. [Google Scholar] [CrossRef]
- Vick, B.A.; Zimmerman, D.C. Biosynthesis of jasmonic Acid by several plant species. Plant Physiol. 1984, 75, 458–461. [Google Scholar] [CrossRef] [PubMed]
- Guy, C.; Kaplan, F.; Kopka, J.; Selbig, J.; Hincha, D.K. Metabolomics of temperature stress. Physiol. Plant. 2008, 132, 220–235. [Google Scholar] [CrossRef] [PubMed]
- Facchini, P.J.; De Luca, V. Phloem-Specific Expression of Tyrosine/Dopa Decarboxylase Genes and the Biosynthesis of Isoquinoline Alkaloids in Opium Poppy. Plant Cell 1995, 7, 1811–1821. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Estiarte, M. Can elevated CO2 affect secondary metabolism and ecosystem function? Trends Ecol. Evol. 1998, 13, 20–24. [Google Scholar] [CrossRef]
- Draths, K.M.; Knop, D.R.; Frost, J.W. Shikimic Acid and Quinic Acid: Replacing Isolation from Plant Sources with Recombinant Microbial Biocatalysis. J. Am. Chem. Soc. 1999, 127, 1603–1604. [Google Scholar] [CrossRef]
- Herrmann, K.M. The Shikimate Pathway: Early Steps in the Biosynthesis of Aromatic Compounds. Am. Soc. Plant Physiol. 1995, 7, 907–919. [Google Scholar]
- Moura, J.C.M.S.; Bonine, C.A.V.; de Oliveira Fernandes Viana, J.; Dornelas, M.C.; Mazzafera, P. Abiotic and biotic stresses and changes in the lignin content and composition in plants. J. Integr. Plant Biol. 2010, 52, 360–376. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Giraldo, L.; Jikumaru, Y.; Kamiya, Y.; Tang, Y.; Dixon, R.A. Selective lignin downregulation leads to constitutive defense response expression in alfalfa (Medicago sativa L.). New Phytol. 2011, 190, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Arbona, V.; Manzi, M.; de Ollas, C.; Gómez-Cadenas, A. Metabolomics as a tool to investigate abiotic stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 4885–4911. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Df | F | P | |
|---|---|---|---|
| Site | 1 | 13.3136 | 0.0005 |
| Temperature | 5 | 1.6354 | 0.0210 |
| Site × Temp | 5 | 1.4068 | 0.0815 |
| (A) | |||||
| Agrostis capillaris L. Long-Term Warmed Site | |||||
| Temperature | +1 °C | +3 °C | +5 °C | +10 °C | +15 °C |
| Control | 0.92 | 1.000 | 1.00 | 0.01 | 0.047 |
| +1 °C | 0.936 | 0.92 | 0.08 | 0.004 | |
| +3 °C | 1.00 | 0.04 | 0.01 | ||
| +5 °C | 0.05 | 0.05 | |||
| +10 °C | 0.03 | ||||
| (B) | |||||
| Agrostis capillaris L. Short-Term Warmed Site | |||||
| Temperature | +1 °C | +3 °C | +5 °C | +10 °C | +15 °C |
| Control | 1.00 | 0.70 | 0.23 | 0.02 | 0.08 |
| +1 °C | 0.93 | 0.49 | 0.04 | 0.07 | |
| +3 °C | 0.98 | 0.07 | 0.07 | ||
| +5 °C | 1.00 | 0.09 | |||
| +10 °C | 0.82 | ||||
| (A) | |||||
| Ranunculus acris L. Long-Term Warmed Site | |||||
| Temperature | +1 °C | +3 °C | +5 °C | +10 °C | |
| Control | 0.05 | 0.14 | 0.99 | 0.40 | |
| +1 °C | 0.92 | 0.05 | 0.53 | ||
| +3 °C | 0.12 | 0.91 | |||
| +5 °C | 0.31 | ||||
| (B) | |||||
| Ranunculus acris L. Short-Term Warmed Site | |||||
| Temperature | +1 °C | +3 °C | +5 °C | +10 °C | +15 °C |
| Control | 0.99 | 1.00 | 0.74 | 0.02 | 0.02 |
| +1 °C | 0.96 | 0.97 | 0.007 | 0.006 | |
| +3 °C | 0.62 | 0.03 | 0.03 | ||
| +5 °C | 0.01 | 0.001 | |||
| +10 °C | 0.89 | ||||
| Df | F | P | |
|---|---|---|---|
| Site | 1 | 2.9063 | 0.0005 |
| Temperature | 5 | 1.9687 | 0.0005 |
| Site × Temp | 4 | 1.0586 | 0.2334 |
| Site | Long-Term Warming Site | Short-Term Warming Site | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Spe | Agrostis Capillaris | Ranunculus Acris | Agrostis Capillaris | Ranunculus Acris | ||||||||||||||||||||
| Temp | A | B | C | D | E | F | A | B | C | D | E | F | A | B | C | D | E | F | A | B | C | D | E | F |
| Amino acid metabolism | ↑↑ | ↓ | - | - | ↑↑ | ↑↑ | - | - | ↑ | ↓ | ↓ | ↓ | - | - | - | - | ↑↑ | ↑↑↑ | - | - | - | ↓ | ↑↑↑ | ↑↑ |
| Biosynthesis of other secondary metabolites | - | ↓ | - | - | ↑↑↑ | ↑↑ | - | ↓↓ | ↑↑ | ↓ | - | ↑↑ | - | - | ↓↓ | ↑ | ↑ | ↑↑↑ | ↑↑ | - | - | - | ↑↑ | - |
| Carbohydrate metabolism | - | - | ↑ | - | ↑↑ | - | ↑ | ↓ | ↓↓ | - | ↓ | ↓↓↓ | - | - | ↓↓ | - | - | ↑ | ↓ | ↓ | ↓ | - | ↑↑↑ | ↑↑ |
| Energy metabolism | - | - | ↑ | - | ↓ | - | - | - | ↓↓↓ | - | ↓↓ | ↓↓↓ | ↑ | ↑↑ | - | ↑↑ | - | - | ↑ | ↑ | ↑↑ | ↓ | ↓↓ | - |
| Metabolism of cofactors and vitamins | ↑↑↑ | ↑↑↑ | ↑↑ | ↑↑ | ↑ | ↑↑ | ↓ | ↓ | ↓ | - | ↓↓ | ↓↓↓ | ↑ | ↑ | - | ↑↑ | ↑ | - | - | ↓ | - | ↓↓↓ | ↓↓↓ | ↓↓ |
| Metabolism of other amino acids | ↑↑↑ | - | ↑↑↑ | ↑↑ | ↑ | ↑↑↑ | ↓↓ | ↓ | ↓ | ↓↓↓ | ↓↓ | ↓ | ↑↑↑ | ↑↑ | ↑ | ↑↑ | ↑↑ | ↑↑ | ↓↓ | ↓↓ | ↓↓ | ↓↓↓ | ↓↓↓ | ↓↓ |
| Metabolism of terpenoids and polyketides | ↑↑ | ↓↓ | - | ↑ | ↑ | ↑↑ | ↓ | ↓↓↓ | - | ↓↓↓ | ↓↓ | ↓↓↓ | - | ↑↑ | - | ↑↑ | - | ↑↑ | ↑ | - | ↑ | - | - | - |
| Xenobiotics biodegradation and metabolism | - | ↑ | ↑ | - | - | - | ↓ | ↓↓↓ | ↓↓↓ | ↓↓↓ | ↓↓↓ | ↓↓↓ | ↓↓ | ↓↓ | ↓↓ | ↑ | ↑ | - | ↓↓ | ↓↓ | ↓↓ | ↑↑ | ↑↑↑ | ↑↑↑ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gargallo-Garriga, A.; Ayala-Roque, M.; Sardans, J.; Bartrons, M.; Granda, V.; Sigurdsson, B.D.; Leblans, N.I.W.; Oravec, M.; Urban, O.; Janssens, I.A.; et al. Impact of Soil Warming on the Plant Metabolome of Icelandic Grasslands. Metabolites 2017, 7, 44. https://doi.org/10.3390/metabo7030044
Gargallo-Garriga A, Ayala-Roque M, Sardans J, Bartrons M, Granda V, Sigurdsson BD, Leblans NIW, Oravec M, Urban O, Janssens IA, et al. Impact of Soil Warming on the Plant Metabolome of Icelandic Grasslands. Metabolites. 2017; 7(3):44. https://doi.org/10.3390/metabo7030044
Chicago/Turabian StyleGargallo-Garriga, Albert, Marta Ayala-Roque, Jordi Sardans, Mireia Bartrons, Victor Granda, Bjarni D. Sigurdsson, Niki I. W. Leblans, Michal Oravec, Otmar Urban, Ivan A. Janssens, and et al. 2017. "Impact of Soil Warming on the Plant Metabolome of Icelandic Grasslands" Metabolites 7, no. 3: 44. https://doi.org/10.3390/metabo7030044
APA StyleGargallo-Garriga, A., Ayala-Roque, M., Sardans, J., Bartrons, M., Granda, V., Sigurdsson, B. D., Leblans, N. I. W., Oravec, M., Urban, O., Janssens, I. A., & Peñuelas, J. (2017). Impact of Soil Warming on the Plant Metabolome of Icelandic Grasslands. Metabolites, 7(3), 44. https://doi.org/10.3390/metabo7030044