Metabolic Analysis of Four Cultivars of Liriope platyphylla


 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Analysis of Spicatoside A
2.2. Analysis of Phenolic Compounds
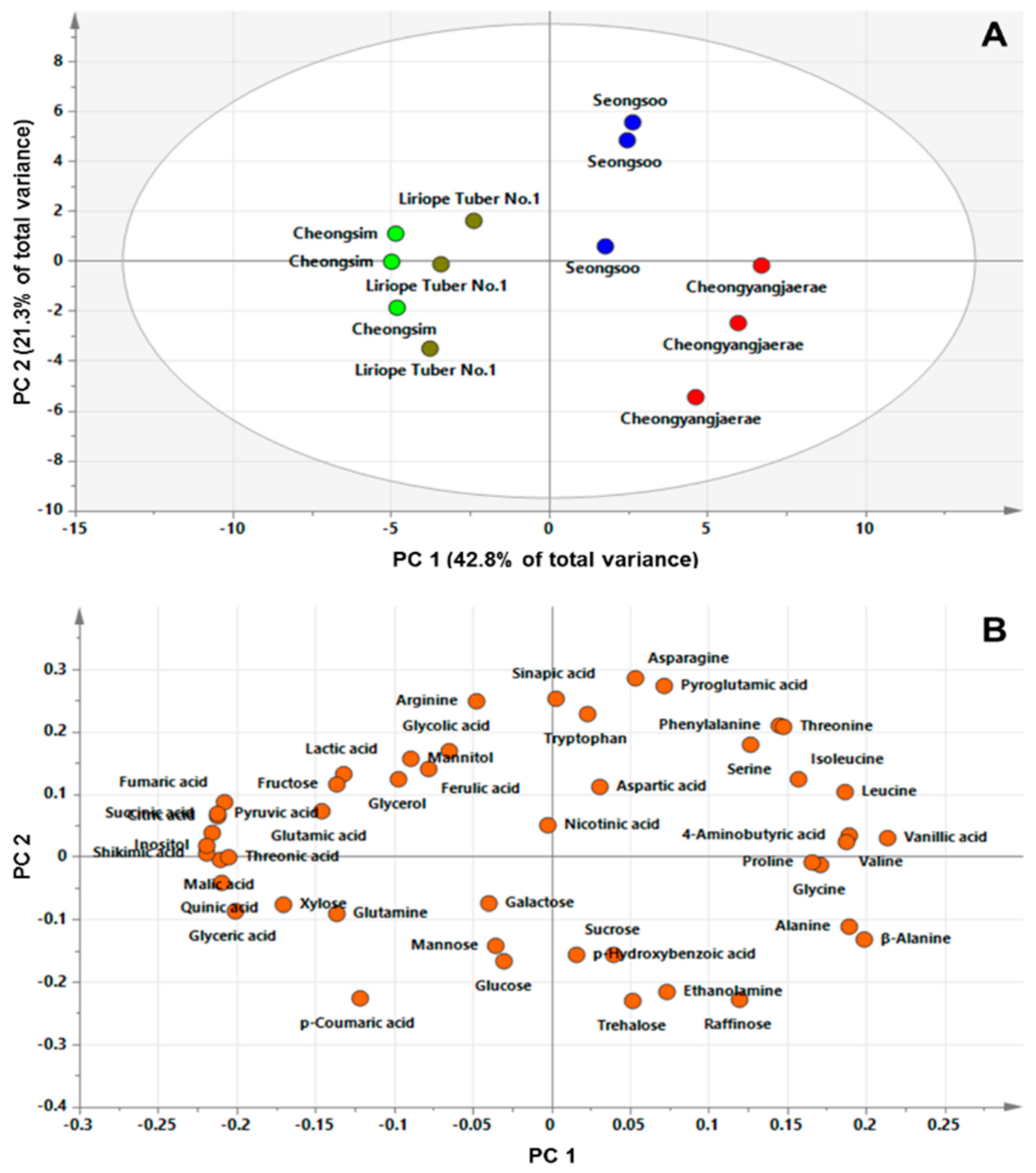
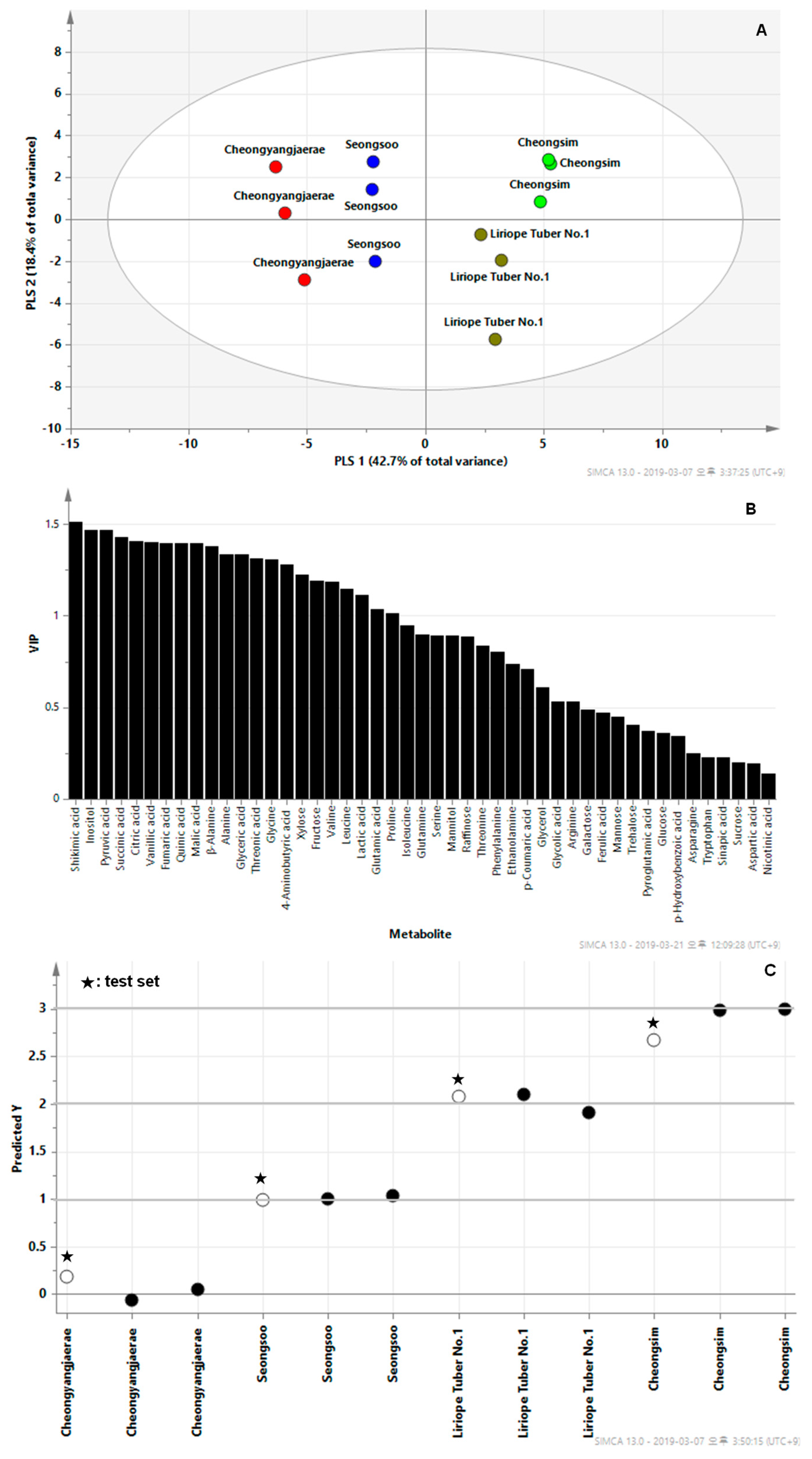
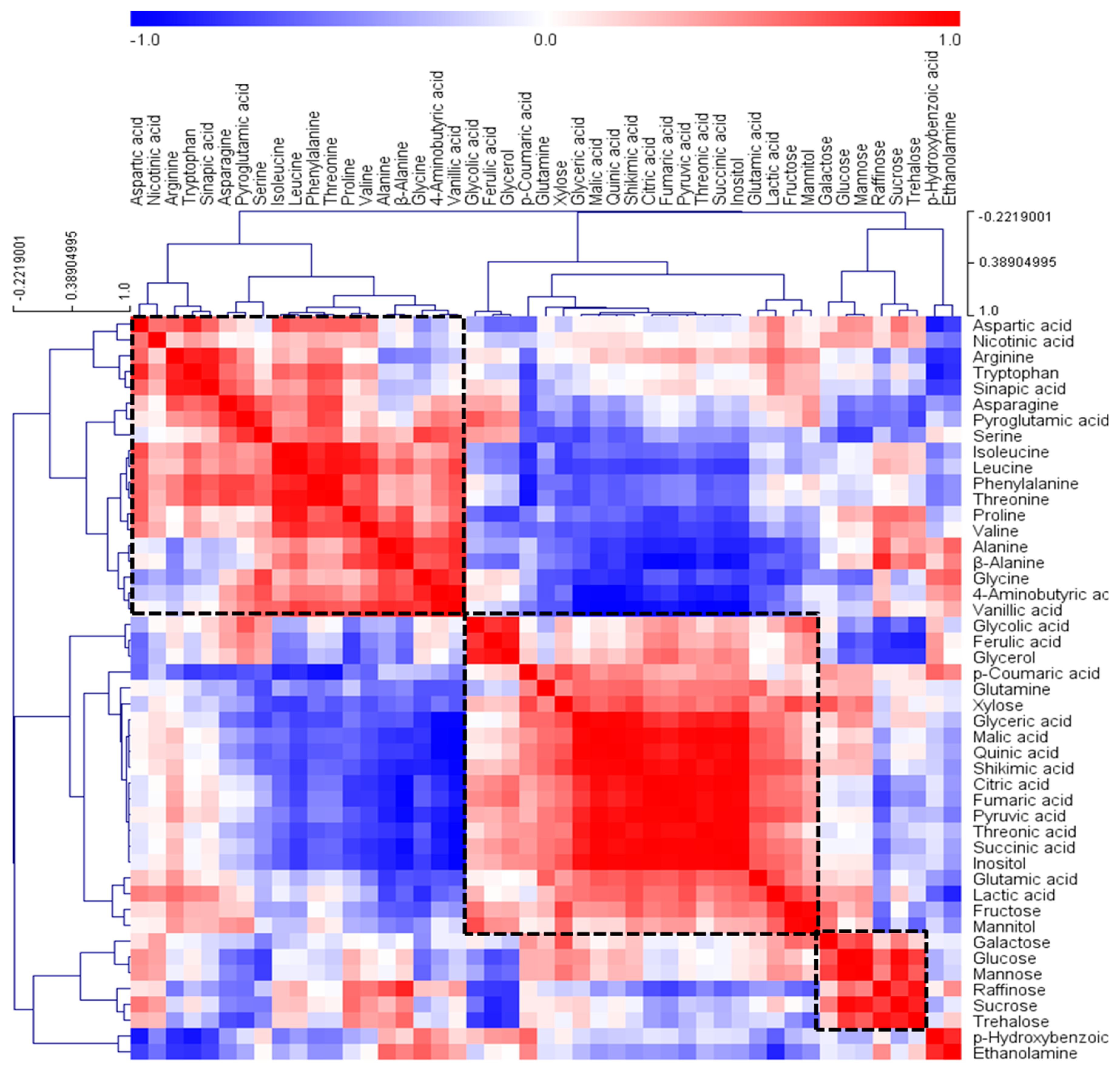
2.3. GC‒TOFMS‒Based Metabolic Profiling and Multivariate Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Spicatoside A Extraction and Quantification
4.3. Phenylpropanoid Extraction and Quantification
4.4. Metabolic Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Park, W.T.; Kim, Y.K.; Kim, Y.S.; Park, N.I.; Lee, S.Y.; Park, S.U. In vitro plant regeneration and micropropagation of Liriope platyphylla. Plant Omics 2011, 4, 199–203. [Google Scholar]
- Kim, W.K.; Pyee, Y.; Chung, H.-J.; Park, H.J.; Hong, J.-Y.; Son, K.H.; Lee, S.K. Antitumor activity of spicatoside A by modulation of autophagy and apoptosis in human colorectal cancer cells. J. Nat. Prod. 2016, 79, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Hur, J.; Lee, P.; Kim, J.; Kim, A.J.; Kim, H.; Kim, S.Y. Induction of nerve growth factor by butanol fraction of Liriope platyphylla in C6 and primary astrocyte cells. Biol. Pharm. Bull. 2004, 27, 1257–1260. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-C.; Wu, C.-C.; Cheng, T.-S.; Kuo, C.-Y.; Tsai, Y.-C.; Chiang, S.-Y.; Wong, T.-S.; Wu, Y.-C.; Chang, F.-R. Active constituents from Liriope platyphylla root against cancer growth in vitro. Evid. Based Complement. Altern. Med. 2013, 2013, 857929. [Google Scholar]
- Yeoman, M.; Yeoman, C. Tansley Review No. 90. Manipulating secondary metabolism in cultured plant cells. New Phytol. 1996, 134, 553–569. [Google Scholar] [CrossRef]
- Aharoni, A.; Galili, G. Metabolic engineering of the plant primary–secondary metabolism interface. Curr. Opin. Biotechnol. 2011, 22, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kim, H.K.; Yang, E.S.; Lee, K.Y.; Du Kim, S.; Kim, Y.C.; Sung, S.H. Optimization of pressurized liquid extraction for spicatoside A in Liriope platyphylla. Sep. Purif. Technol. 2010, 71, 168–172. [Google Scholar] [CrossRef]
- Bennett, R.N.; Wallsgrove, R.M. Secondary metabolites in plant defence mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef] [Green Version]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Kapoor, H.C. Anti-oxidant activity and total phenolic content of some Asian vegetables. Int. J. Food Sci. Tech. 2002, 37, 153–161. [Google Scholar] [CrossRef]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- McCann, M.; Gill, C.; O’Brien, G.; Rao, J.; McRoberts, W.; Hughes, P.; McEntee, R.; Rowland, I. Anti-cancer properties of phenolics from apple waste on colon carcinogenesis in vitro. Food Chem. Toxicol. 2007, 45, 1224–1230. [Google Scholar] [CrossRef]
- Villas-Boas, S.G.; Nielsen, J.; Smedsgaard, J.; Hansen, M.A.; Roessner-Tunali, U. Metabolome Analysis: An Introduction, 1st ed.; JohnWiley & Sons: Hoboken, NJ, USA, 2007; pp. 1–319. [Google Scholar]
- Roessner, U.; Luedemann, A.; Brust, D.; Fiehn, O.; Linke, T.; Willmitzer, L.; Fernie, A.R. Metabolic profiling allows comprehensive phenotyping of genetically or environmentally modified plant systems. Plant Cell 2001, 13, 11–29. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Liang, Y.; Dunn, W.B.; Shen, H.; Kell, D.B. Comparative evaluation of software for deconvolution of metabolomics data based on GC-TOF-MS. TrAC Trends Anal. Chem. 2008, 27, 215–227. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Ellis, D.I.; Brown, M.; Halsall, A.; O’hagan, S.; Spasic, I.; Tseng, A.; Kell, D.B. A GC-TOF-MS study of the stability of serum and urine metabolomes during the UK Biobank sample collection and preparation protocols. Int. J. Epidemiol. 2008, 37, i23–i30. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.; Chen, J.-H.; Wei, D.; Chen, F.; Chen, G. Global metabolic regulation of the snow alga Chlamydomonas nivalis in response to nitrate or phosphate deprivation by a metabolome profile analysis. Int. J. Mol. Sci. 2016, 17, 694. [Google Scholar] [CrossRef]
- Lei, Z.; Huhman, D.V.; Sumner, L.W. Mass spectrometry strategies in metabolomics. J. Biol. Chem. 2011, 286, 25435–25442. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Park, S.-Y.; Lim, S.-H.; Yeo, Y.; Cho, H.S.; Ha, S.-H. Comparative metabolic profiling of pigmented rice (Oryza sativa L.) cultivars reveals primary metabolites are correlated with secondary metabolites. J. Cereal Sci. 2013, 57, 14–20. [Google Scholar] [CrossRef]
- Park, C.H.; Baskar, T.B.; Park, S.-Y.; Kim, S.-J.; Valan Arasu, M.; Al-Dhabi, N.A.; Kim, J.K.; Park, S.U. Metabolic profiling and antioxidant assay of metabolites from three radish cultivars (Raphanus sativus). Molecules 2016, 21, 157. [Google Scholar] [CrossRef]
- Carreno-Quintero, N.; Acharjee, A.; Maliepaard, C.; Bachem, C.W.; Mumm, R.; Bouwmeester, H.; Visser, R.G.; Keurentjes, J.J. Untargeted metabolic quantitative trait loci analyses reveal a relationship between primary metabolism and potato tuber quality. Plant Physiol. 2012, 158, 1306–1318. [Google Scholar] [CrossRef] [PubMed]
- Yaseen, M.; Ahmad, T.; Sablok, G.; Standardi, A.; Hafiz, I.A. Role of carbon sources for in vitro plant growth and development. Mol. Biol. Rep. 2013, 40, 2837–2849. [Google Scholar] [CrossRef] [PubMed]
- Yeo, H.J.; Park, C.H.; Lee, K.B.; Kim, J.K.; Park, J.S.; Lee, J.W.; Park, S.U. Metabolic analysis of vigna unguiculata sprouts exposed to different light-emitting diodes. Nat. Prod. Commun. 2018, 13, 1349–1354. [Google Scholar] [CrossRef]
- Park, C.H.; Yeo, H.J.; Kim, N.S.; Park, Y.E.; Park, S.-Y.; Kim, J.K.; Park, S.U. Metabolomic profiling of the white, violet, and red flowers of rhododendron schlippenbachii maxim. Molecules 2018, 23, 827. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Yeo, H.J.; Baskar, T.B.; Kim, J.K.; Park, S.U. Metabolic profiling and chemical-based antioxidant assays of green and red lettuce (Lactuca sativa). Nat. Prod. Commun. 2018, 13, 315–322. [Google Scholar] [CrossRef]
- Zulak, K.G.; Weljie, A.M.; Vogel, H.J.; Facchini, P.J. Quantitative 1 H NMR metabolomics reveals extensive metabolic reprogramming of primary and secondary metabolism in elicitor-treated opium poppy cell cultures. BMC Plant Biol. 2008, 8, 5. [Google Scholar] [CrossRef]
- Muhlemann, J.K.; Maeda, H.; Chang, C.-Y.; San Miguel, P.; Baxter, I.; Cooper, B.; Perera, M.A.; Nikolau, B.J.; Vitek, O.; Morgan, J.A. Developmental changes in the metabolic network of snapdragon flowers. PLoS ONE 2012, 7, e40381. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.H.; Rosa, E. Free amino acid composition in primary and secondary inflorescences of 11 broccoli (Brassica oleracea var italica) cultivars and its variation between growing seasons. J. Sci. Food Agric. 2001, 81, 295–299. [Google Scholar] [CrossRef]
- Kim, J.K.; Bong, S.J.; Park, S.U. Amino acid content in different cultivars of Liriope platyphylla. Asian J. Chem. 2016, 28, 1754–1756. [Google Scholar] [CrossRef]
- Cho, S.-J.; Bang, M.-H.; Lee, I.-J.; Park, C.-G.; Kim, M.-S.; Kim, K.-S.; Sung, J.-D.; Baek, N.-I. Cytotoxicity of steroid-saponins from the tuber of Liriope platyphylla WT. Appl. Biol. Chem. 1998, 41, 390–394. [Google Scholar]
- Shin, J. Saponin composition of Liriope platyphylla and Ophiopogon japonicus. Korean J. Crop Sci. 2002, 47, 236–239. [Google Scholar]
- Lee, K.-S.; Kim, G.-H.; Kim, H.-H.; Choi, J.-W.; Lee, H.-C.; Song, M.-R.; Kim, M.-R.; Lee, G.-H. Physicochemical characteristics of Liriope platyphylla tubers by drying process. J. Korean Soc. Food Sci. Nutr. 2009, 38, 1104–1110. [Google Scholar] [CrossRef]
- Choi, S.J.; Choi, J.; Jeon, H.; Bae, S.K.; Ko, J.; Kim, J.; Yoon, K.D. Application of high-performance countercurrent chromatography for the isolation of steroidal saponins from Liriope plathyphylla. J. Sep. Sci. 2015, 38, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Yeo, H.J.; Park, Y.J.; Morgan, A.; Valan Arasu, M.; Al-Dhabi, N.A.; Park, S.U. Influence of indole-3-acetic acid and gibberellic acid on phenylpropanoid accumulation in common buckwheat (Fagopyrum esculentum Moench) sprouts. Molecules 2017, 22, 374. [Google Scholar] [CrossRef]
- Park, C.H.; Park, S.Y.; Lee, S.Y.; Kim, J.K.; Park, S.U. Analysis of metabolites in white flowers of magnolia denudata desr. and violet flowers of Magnolia Liliiflora Desr. Molecules 2018, 23, 1588. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Yeo, H.J.; Kim, N.S.; Eun, P.Y.; Kim, S.-J.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.-Y.; Kim, J.K.; Park, S.U. Metabolic profiling of pale green and purple kohlrabi (Brassica oleracea var. gongylodes). Appl. Biol. Chem. 2017, 60, 249–257. [Google Scholar] [CrossRef]
- Viant, M.R.; Kurland, I.J.; Jones, M.R.; Dunn, W.B. How close are we to complete annotation of metabolomes? Curr. Opin. Chem. Biol. 2017, 36, 64–69. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| No. | Cultivars | Contents (mg/g) |
|---|---|---|
| 1 | Liriope Tuber No. 1 | 1.83 ± 0.13 a 1 |
| 2 | Cheongyangjaerae | 1.25 ± 0.01 b |
| 3 | Seongsoo | 1.01 ± 0.02 c |
| 4 | Cheongsim | 1.09 ± 0.04 c |
| Contents (mg/g). | Liriope Tuber No. 1 | Cheongyangjaerae | Seongsoo | Cheongsim |
|---|---|---|---|---|
| Gallic acid | 29.07 ± 0.43 d 1 | 30.79 ± 1.37 c | 33.54 ± 0.94 b | 47.21 ± 0.30 a |
| Catechin | 105.37 ± 0.43 a | 105.65 ± 0.58 a | 105.72 ± 0.12 a | 105.54 ± 1.11 a |
| Chlorogenic acid | N.D | N.D | N.D | 82.43 ± 1.39 a |
| Caffeic acid | 38.17 ± 0.10 d | 41.84 ± 0.54 c | 42.89 ± 0.39 b | 52.31 ± 0.76 a |
| Benzoic acid | 220.44 ± 7.39 a | 161.75 ± 5.12 d | 191.78 ± 1.57 b | 173.62 ± 2.51 c |
| Total | 393.05 ± 7.87 b | 340.03 ± 4.29 d | 373.93 ± 0.74 c | 461.11 ± 4.96 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, C.H.; Morgan, A.M.A.; Park, B.B.; Lee, S.Y.; Lee, S.; Kim, J.K.; Park, S.U. Metabolic Analysis of Four Cultivars of Liriope platyphylla. Metabolites 2019, 9, 59. https://doi.org/10.3390/metabo9030059
Park CH, Morgan AMA, Park BB, Lee SY, Lee S, Kim JK, Park SU. Metabolic Analysis of Four Cultivars of Liriope platyphylla. Metabolites. 2019; 9(3):59. https://doi.org/10.3390/metabo9030059
Chicago/Turabian StylePark, Chang Ha, Abubaker Mohammed Awad Morgan, Byung Bae Park, Sook Young Lee, Sanghyun Lee, Jae Kwang Kim, and Sang Un Park. 2019. "Metabolic Analysis of Four Cultivars of Liriope platyphylla" Metabolites 9, no. 3: 59. https://doi.org/10.3390/metabo9030059
APA StylePark, C. H., Morgan, A. M. A., Park, B. B., Lee, S. Y., Lee, S., Kim, J. K., & Park, S. U. (2019). Metabolic Analysis of Four Cultivars of Liriope platyphylla. Metabolites, 9(3), 59. https://doi.org/10.3390/metabo9030059