Back to GroEL-Assisted Protein Folding: GroES Binding-Induced Displacement of Denatured Proteins from GroEL to Bulk Solution



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
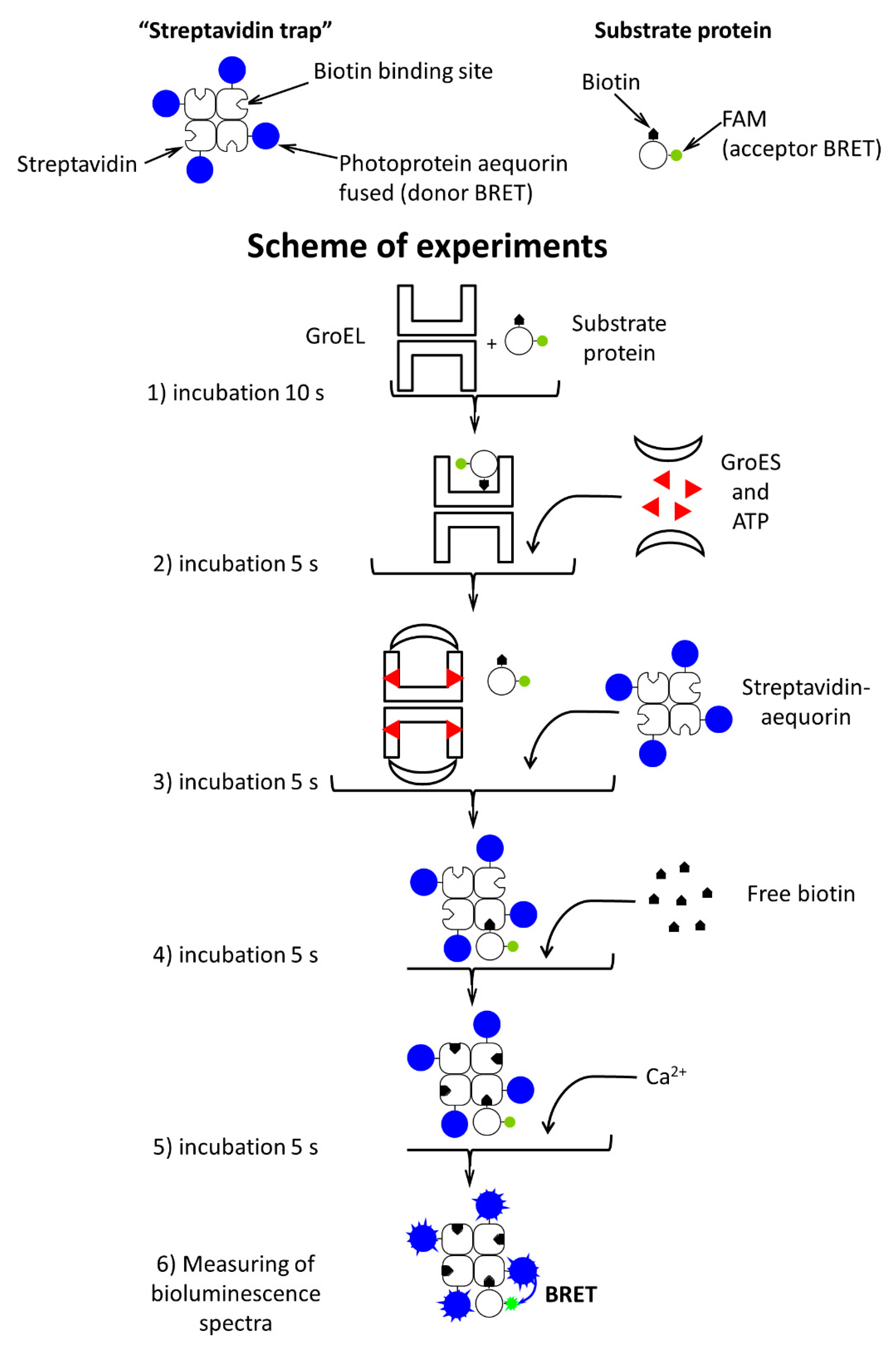
2. Materials and Methods
2.1. Solutions
2.2. Proteins
2.3. Methods
3. Results
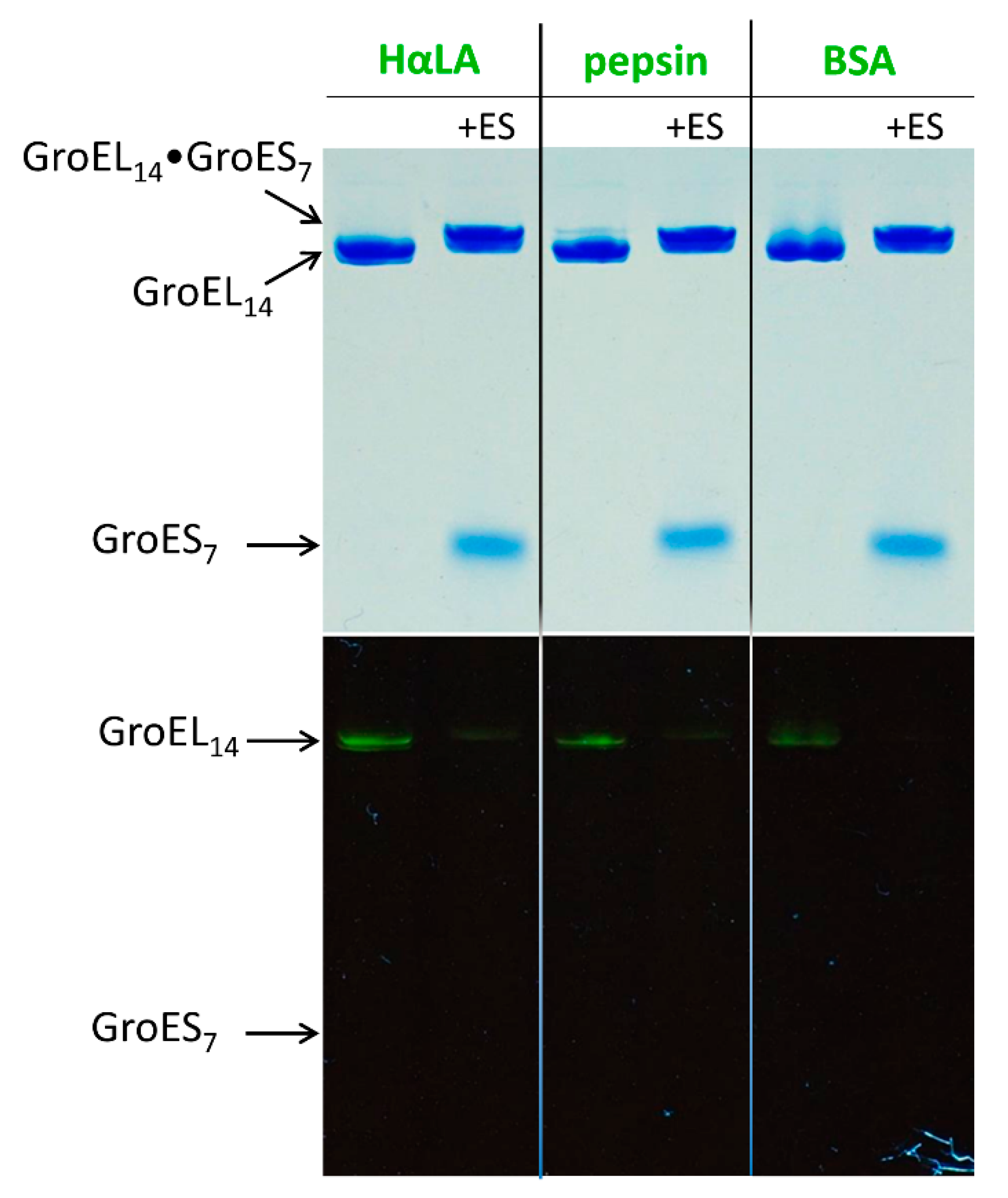
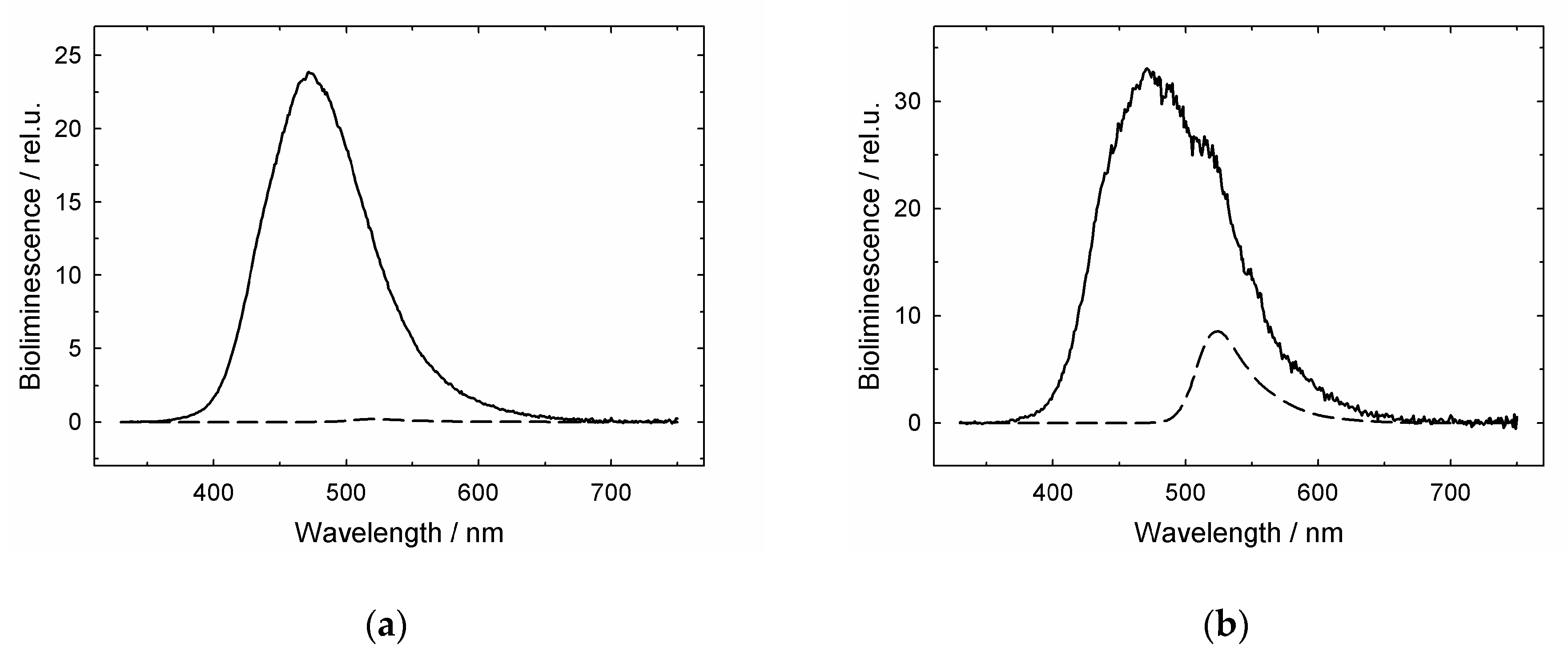
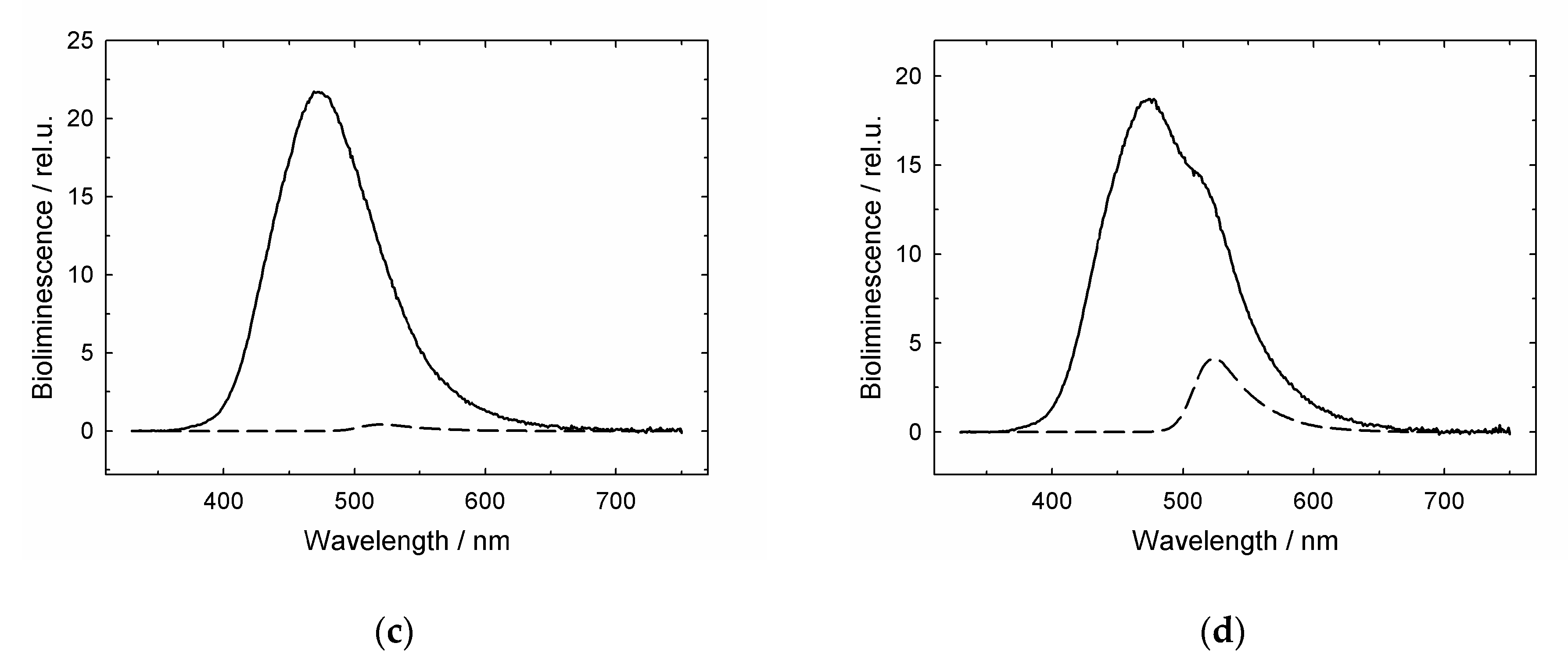
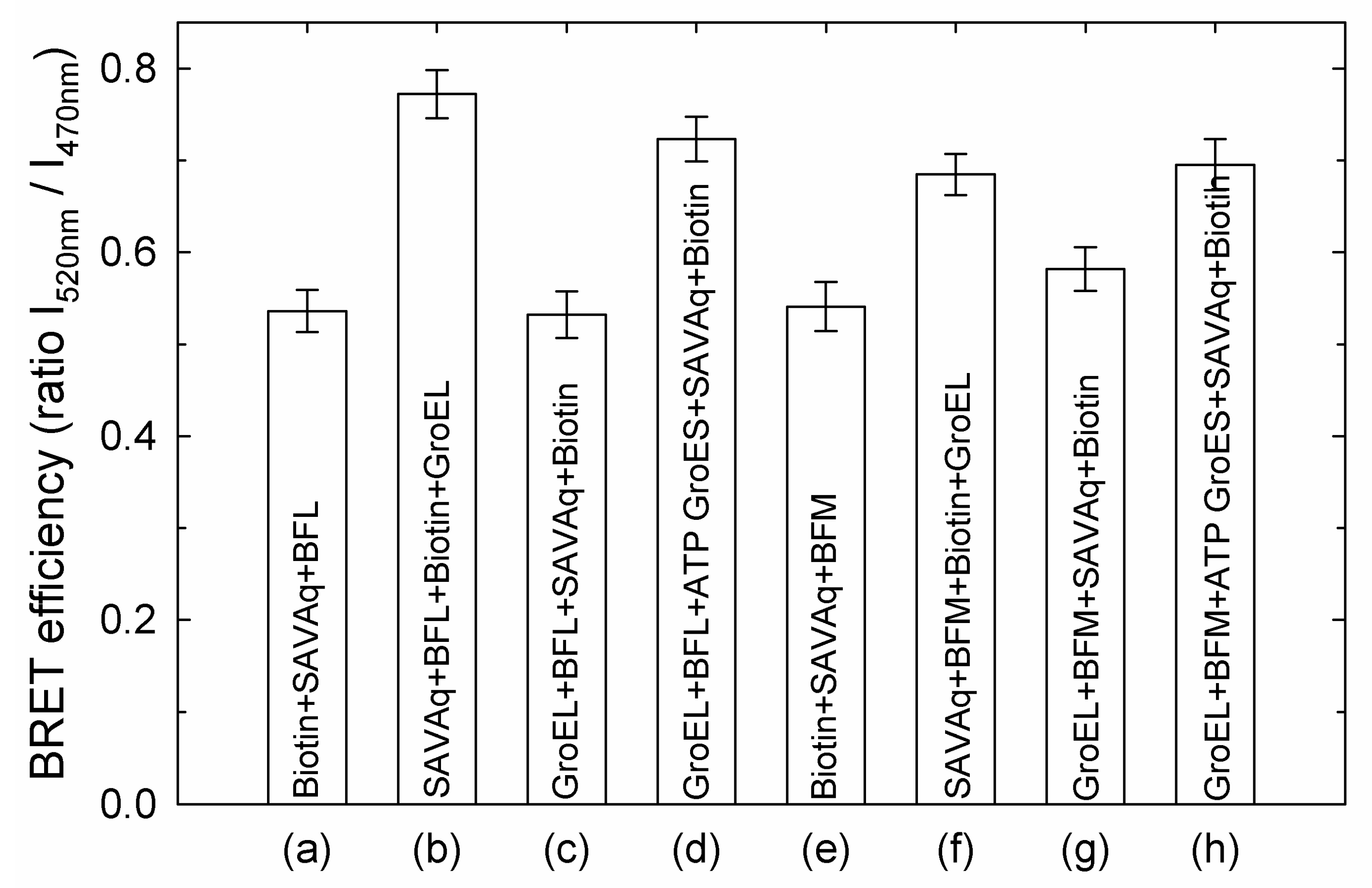
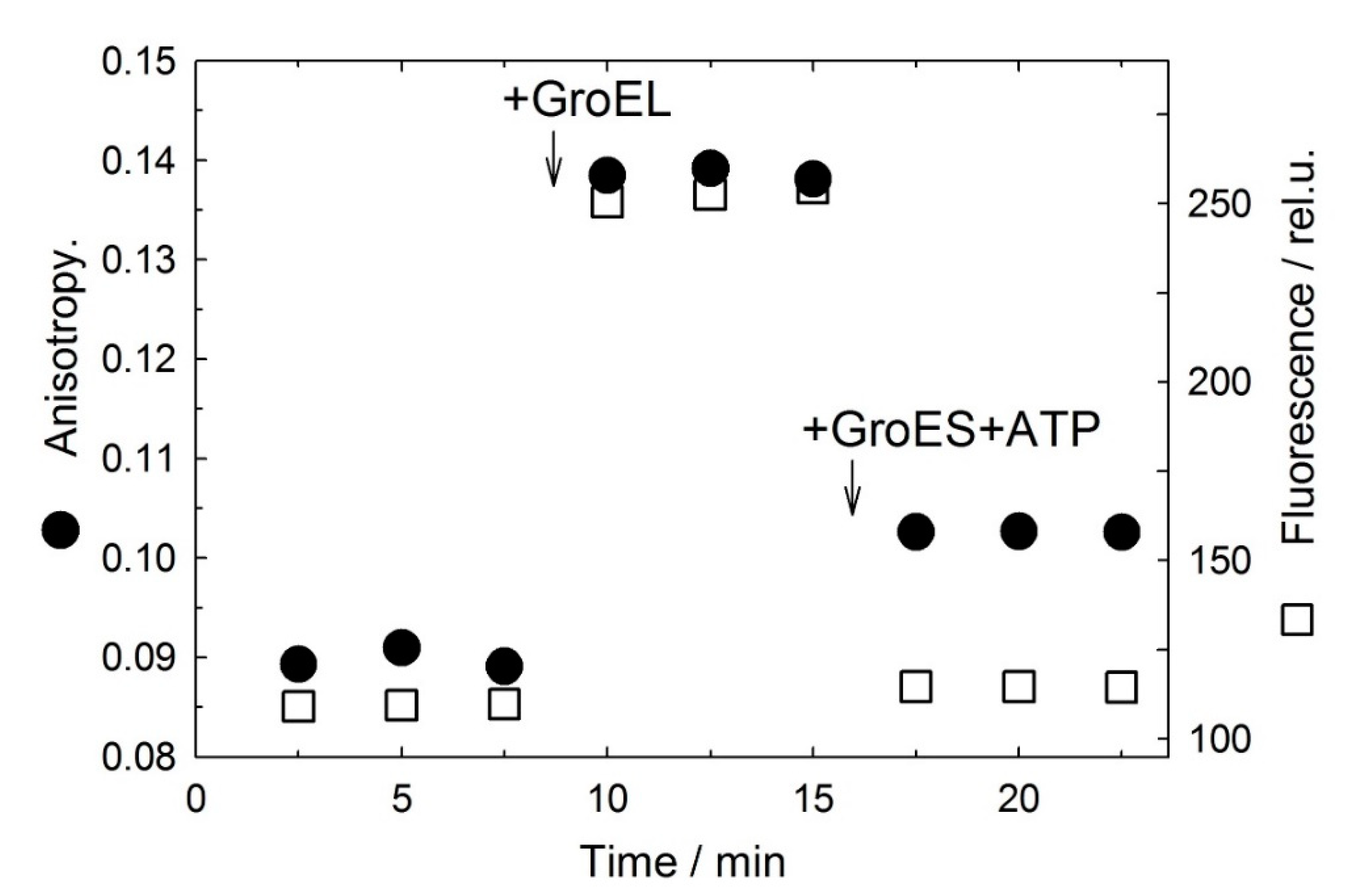
3.1. GroES Essentially Decreases the GroEL Affinity for Denatured Proteins
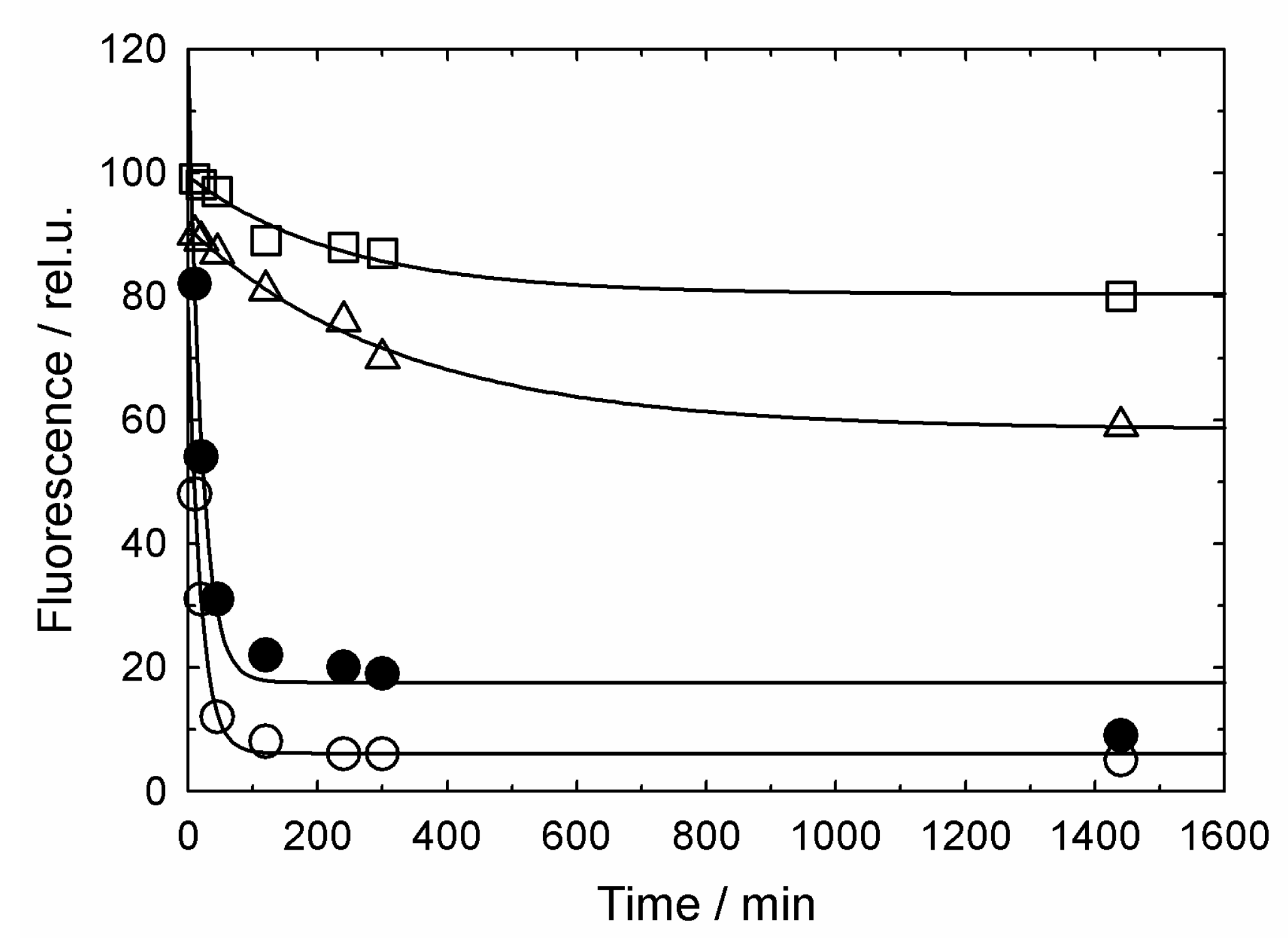
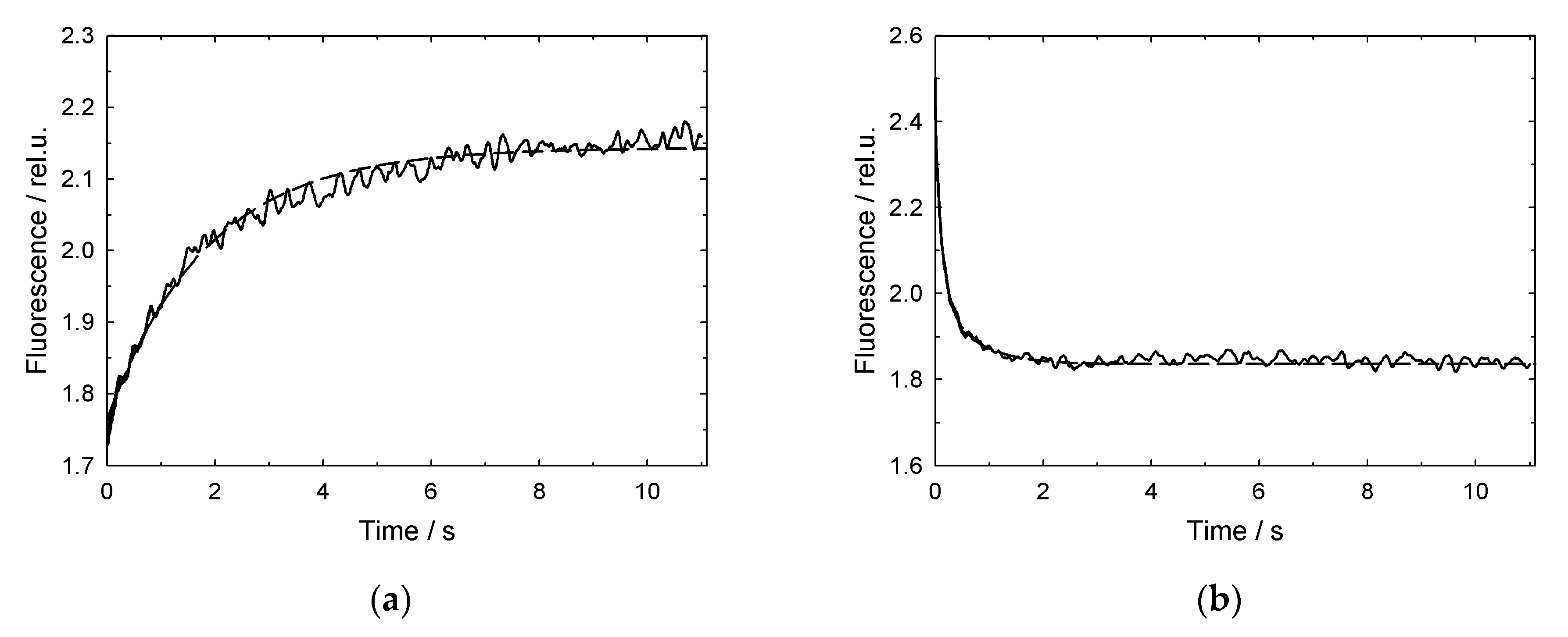
3.2. GroES-Assisted Dissociation of Denatured Proteins from the GroEL Surface Takes Much Less Time than the ATPase Cycle
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Anfinsen, C.B. Principles that govern the folding of protein chains. Science 1973, 181, 223–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ptitsyn, O.B.; Semisotnov, G.V. The mechanism of protein folding. In Conformations and Forces in Protein Folding; Nail, B.T., Dill, K.A., Eds.; AAAS: Washington, DC, USA, 1991; pp. 155–168. [Google Scholar]
- Seckler, R.; Jaenicke, R. Protein folding and protein refolding. FASEB J. 1992, 6, 2545–2552. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J. Proteins as molecular chaperones. Nature 1987, 328, 378–379. [Google Scholar] [CrossRef]
- Gething, M.J.; Sambrook, J. Protein folding in the cell. Nature 1992, 355, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, S.; Craig, E.A. The Heat-Shock Proteins. Annu. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef] [PubMed]
- Koike, H.; Katsuno, M. Ultrastructure in Transthyretin Amyloidosis: From Pathophysiology to Therapeutic Insights. Biomedicines 2019, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horwich, A.L.; Fenton, W.A.; Chapman, E.; Farr, G.W. Two families of chaperonin: Physiology and mechanism. Annu. Rev. Cell Dev. Biol. 2007, 23, 115–145. [Google Scholar] [CrossRef]
- Chen, S.; Roseman, A.M.; Hunter, A.S.; Wood, S.P.; Burston, S.G.; Ranson, N.A.; Clarke, A.R.; Saibil, H.R. Location of a folding protein and shape changes in GroEL-GroES complexes imaged by cryo-electron microscopy. Nature 1994, 371, 261–264. [Google Scholar] [CrossRef]
- Harris, J.R.; Plückthun, A.; Zahn, R. Transmission Electron Microscopy of GroEL, GroES, and the Symmetrical GroEL/ES Complex. J. Struct. Biol. 1994, 112, 216–230. [Google Scholar] [CrossRef]
- Braig, K.; Otwinowski, Z.; Hegde, R.; Boisvert, D.C.; Joachimiak, A.; Horwich, A.L.; Sigler, P.B. The crystal structure of the bacterial chaperonin GroEL at 2.8 A. Nature 1994, 371, 578–586. [Google Scholar] [CrossRef]
- Braig, K.; Adams, P.D.; Brünger, A.T. Conformational variability in the refined structure of the chaperonin groel at 2.8 Â resolution. Nat. Struct. Biol. 1995, 2, 1095–1101. [Google Scholar] [CrossRef] [PubMed]
- Boisvert, D.C.; Wang, J.; Otwinowski, Z.; Horwich, A.L.; Sigler, P.B. The 2.4 Å crystal structure of the bacterial chaperonin GroEL complexed with ATPγS. Nat. Struct. Biol. 1996, 3, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Horwich, A.L.; Sigler, P.B. The crystal structure of the asymmetric GroEL-GroES-(ADP)7 chaperonin complex. Nature 1997, 388, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J.F.; Weaver, A.J.; Landry, S.J.; Gierasch, L.; Deisenhofer, J. The crystal structure of the GroES co-chaperonin at 2.8 Å resolution. Nature 1996, 379, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Mayhew, M.; da Silva, A.C.R.; Martin, J.; Erdjument-Bromage, H.; Tempst, P.; Hartl, F.U. Protein folding in the central cavity of the GroEL–GroES chaperonin complex. Nature 1996, 379, 420–426. [Google Scholar] [CrossRef]
- Hayer-Hartl, M.; Bracher, A.; Hartl, F.U. The GroEL–GroES Chaperonin Machine: A Nano-Cage for Protein Folding. Trends Biochem. Sci. 2016, 41, 62–76. [Google Scholar] [CrossRef]
- Shtilerman, M.; Lorimer, G.H.; Englander, S.W. Chaperonin Function: Folding by Forced Unfolding. Science 1999, 284, 822. [Google Scholar] [CrossRef] [Green Version]
- Horwich, A.L.; Farr, G.W.; Fenton, W.A. GroEL−GroES-Mediated Protein Folding. Chem. Rev. 2006, 106, 1917–1930. [Google Scholar] [CrossRef]
- Lin, Z.; Madan, D.; Rye, H.S. GroEL stimulates protein folding through forced unfolding. Nat. Struct. Mol. Biol. 2008, 15, 303–311. [Google Scholar] [CrossRef]
- Priya, S.; Sharma, S.K.; Sood, V.; Mattoo, R.U.H.; Finka, A.; Azem, A.; De Los Rios, P.; Goloubinoff, P. GroEL and CCT are catalytic unfoldases mediating out-of-cage polypeptide refolding without ATP. Proc. Natl. Acad. Sci. USA 2013, 110, 7199–7204. [Google Scholar] [CrossRef] [Green Version]
- Horwich, A.L.; Fenton, W.A. Chaperonin-mediated protein folding: Using a central cavity to kinetically assist polypeptide chain folding. Q. Rev. Biophys. 2009, 42, 83–116. [Google Scholar] [CrossRef] [PubMed]
- Libich, D.S.; Tugarinov, V.; Clore, G.M. Intrinsic unfoldase/foldase activity of the chaperonin GroEL directly demonstrated using multinuclear relaxation-based NMR. Proc. Natl. Acad. Sci. USA 2015, 112, 8817–8823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchenko, N.Y.; Marchenkov, V.V.; Semisotnov, G.V.; Finkelstein, A.V. Strict experimental evidence that apo-chaperonin GroEL does not accelerate protein folding, although it does accelerate one of its steps. Proc. Natl. Acad. Sci. USA 2015, 112, E6831–E6832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahn, R.; Buckle, A.M.; Perrett, S.; Johnson, C.M.; Corrales, F.J.; Golbik, R.; Fersht, A.R. Chaperone activity and structure of monomeric polypeptide binding domains of GroEL. Proc. Natl. Acad. Sci. USA 1996, 93, 15024–15029. [Google Scholar] [CrossRef] [Green Version]
- Jain, N.; Knowles, T.J.; Lund, P.A.; Chaudhuri, T.K. Minichaperone (GroEL191-345) mediated folding of MalZ proceeds by binding and release of native and functional intermediates. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 941–951. [Google Scholar] [CrossRef]
- Chatellier, J.; Hill, F.; Lund, P.A.; Fersht, A.R. In vivo activities of GroEL minichaperones. Proc. Natl. Acad. Sci. USA 1998, 95, 9861–9866. [Google Scholar] [CrossRef] [Green Version]
- Marchenkov, V.V.; Semisotnov, G.V. GroEL-assisted protein folding: Does it occur within the chaperonin inner cavity? Int. J. Mol. Sci. 2009, 10, 2066–2083. [Google Scholar] [CrossRef] [Green Version]
- Marchenkov, V.V.; Sokolovskii, I.V.; Kotova, N.V.; Galzitskaya, O.V.; Bochkareva, E.S.; Girshovich, A.S.; Semisotnov, G.V. The interaction of the GroEL chaperone with early kinetic intermediates of renaturing proteins inhibits the formation of their native structure. Biofizika 2004, 49, 987–994. [Google Scholar]
- Staniforth, R.A.; Burston, S.G.; Atkinson, T.; Clarke, A.R. Affinity of chaperonin-60 for a protein substrate and its modulation by nucleotides and chaperonin-10. Biochem. J. 1994, 300, 651–658. [Google Scholar] [CrossRef] [Green Version]
- Makio, T.; Takasu-Ishikawa, E.; Kuwajima, K. Nucleotide-induced transition of GroEL from the high-affinity to the low-affinity state for a target protein: Effects of ATP and ADP on the GroEL-affected refolding of α-lactalbumin. J. Mol. Biol. 2001, 312, 555–567. [Google Scholar] [CrossRef]
- Tsurupa, G.P.; Ikura, T.; Makio, T.; Kuwajima, K. Refolding kinetics of staphylococcal nuclease and its mutants in the presence of the chaperonin GroEL. J. Mol. Biol. 1998, 277, 733–745. [Google Scholar] [CrossRef] [PubMed]
- Buchner, J.; Schmidt, M.; Fuchs, M.; Jaenicke, R.; Rudolph, R.; Schmid, F.X.; Kiefhaber, T. GroE facilitates refolding of citrate synthase by suppressing aggregation. Biochemistry 1991, 30, 1586–1591. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.E.; Gafni, A. Thermal switching between enhanced and arrested reactivation of bacterial glucose-6-phosphate dehydrogenase assisted by GroEL in the absence of ATP. J. Biol. Chem. 1993, 268, 21632–21636. [Google Scholar]
- Ayling, A.; Baneyx, F. Influence of the GroE molecular chaperone machine on the in vitro refolding of Escherichia coli beta-galactosidase. Protein Sci. 1996, 5, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Viitanen, P.V.; Gatenby, A.A.; Lorimer, G.H. Purified chaperonin 60 (groEL) interacts with the nonnative states of a multitude of Escherichia coli proteins. Protein Sci. 1992, 1, 363–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimm, R.; Donaldson, G.K.; Van Der Vies, S.M.; Schäfer, E.; Gatenby, A.A. Chaperonin-mediated reconstitution of the phytochrome photoreceptor. J. Biol. Chem. 1993, 268, 5220–5226. [Google Scholar]
- Lissin, N.M.; Venyaminov, S.Y.; Girshovich, A.S. (Mg-ATP)-dependent self-assembly of molecular chaperone GroEL. Nature 1990, 348, 339–342. [Google Scholar] [CrossRef]
- Ryabova, N.; Marchenkov, V.; Kotova, N.; Semisotnov, G. Chaperonin GroEL reassembly: An effect of protein ligands and solvent composition. Biomolecules 2014, 4, 458–473. [Google Scholar] [CrossRef] [Green Version]
- Hayer-Hartl, M.K.; Hartl, F.U. A comment on: “The aromatic amino acid content of the bacterial chaperone protein groEL (cpn60): Evidence for the presence of a single tryptophan”, by N.C. Price, S.M. Kelly, S. Wood and A. auf der Mauer (1991) FEBS Lett. 292, 9–12. FEBS Lett. 1993, 320, 83–84, discussion 85. [Google Scholar] [CrossRef] [Green Version]
- Gorokhovatsky, A.Y.; Rudenko, N.V.; Marchenkov, V.V.; Skosyrev, V.S.; Arzhanov, M.A.; Burkhardt, N.; Zakharov, M.V.; Semisotnov, G.V.; Vinokurov, L.M.; Alakhov, Y.B. Homogeneous assay for biotin based on Aequorea victoria bioluminescence resonance energy transfer system. Anal. Biochem. 2003, 313, 68–75. [Google Scholar] [CrossRef]
- Plotnikov, A.N.; Vasilenko, K.S.; Kirkitadze, M.D.; Kotova, N.V.; Motuz, L.P.; Korotkov, K.V.; Semisotnov, G.V.; Alakhov, I.B. Biosynthesis and conformational state of 17-kDa and 27-kDa N-terminal fragments of elongation factor EF-2 in solution. Bioorg. Khim. 1996, 22, 489–502. [Google Scholar] [PubMed]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchenkov, V.; Gorokhovatsky, A.; Marchenko, N.; Ivashina, T.; Semisotnov, G. Back to GroEL-Assisted Protein Folding: GroES Binding-Induced Displacement of Denatured Proteins from GroEL to Bulk Solution. Biomolecules 2020, 10, 162. https://doi.org/10.3390/biom10010162
Marchenkov V, Gorokhovatsky A, Marchenko N, Ivashina T, Semisotnov G. Back to GroEL-Assisted Protein Folding: GroES Binding-Induced Displacement of Denatured Proteins from GroEL to Bulk Solution. Biomolecules. 2020; 10(1):162. https://doi.org/10.3390/biom10010162
Chicago/Turabian StyleMarchenkov, Victor, Andrey Gorokhovatsky, Natalia Marchenko, Tanya Ivashina, and Gennady Semisotnov. 2020. "Back to GroEL-Assisted Protein Folding: GroES Binding-Induced Displacement of Denatured Proteins from GroEL to Bulk Solution" Biomolecules 10, no. 1: 162. https://doi.org/10.3390/biom10010162
APA StyleMarchenkov, V., Gorokhovatsky, A., Marchenko, N., Ivashina, T., & Semisotnov, G. (2020). Back to GroEL-Assisted Protein Folding: GroES Binding-Induced Displacement of Denatured Proteins from GroEL to Bulk Solution. Biomolecules, 10(1), 162. https://doi.org/10.3390/biom10010162