The Morphology and Phenotype of Monocyte-Macrophages When Cultured on Bionanofilms Substrates with Different Surface Relief Profiles

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Selection
2.2. The Study of Biopolymer Films
2.2.1. PHA-Bionanofilm Samples
- Film 1: Copolymer of 3-hydroxybutyrate and of 4-hydroxybutyrate [P (3HB/4HB) 92.0/8.0].
- Film 2: Copolymer of 3-hydroxybutyrate, 3-hydroxyvalerate and 3-hydroxyhexanoate [P (3HB/3HV/3HHx) 66.4/23.4/10.2].
- Film 3: Copolymer of 3-hydroxybutyrate, 3-hydroxyvalerate, 4-hydroxybutyrate and 3-hydroxyhexanoate [P (3HB/3HV/4HB/3HHx) 63.5/19.4/12.3/4.8].
- Film 4: Copolymer of 3-hydroxybutyrate and 3-hydroxyvalerate [P (3HB/3HV) 85.0/15.0].
- Film 5: Poly-3-hydroxybutyrate (P3HB) 100.
2.2.2. Atomic Force Microscopy
2.3. Isolation and Cultivation of Monocytes
2.4. Antigenic Spectrum of the MN Population
2.5. Electron Microscopy
3. Results
3.1. 3-D Surface Topography of PHA Bionanofilm Samples
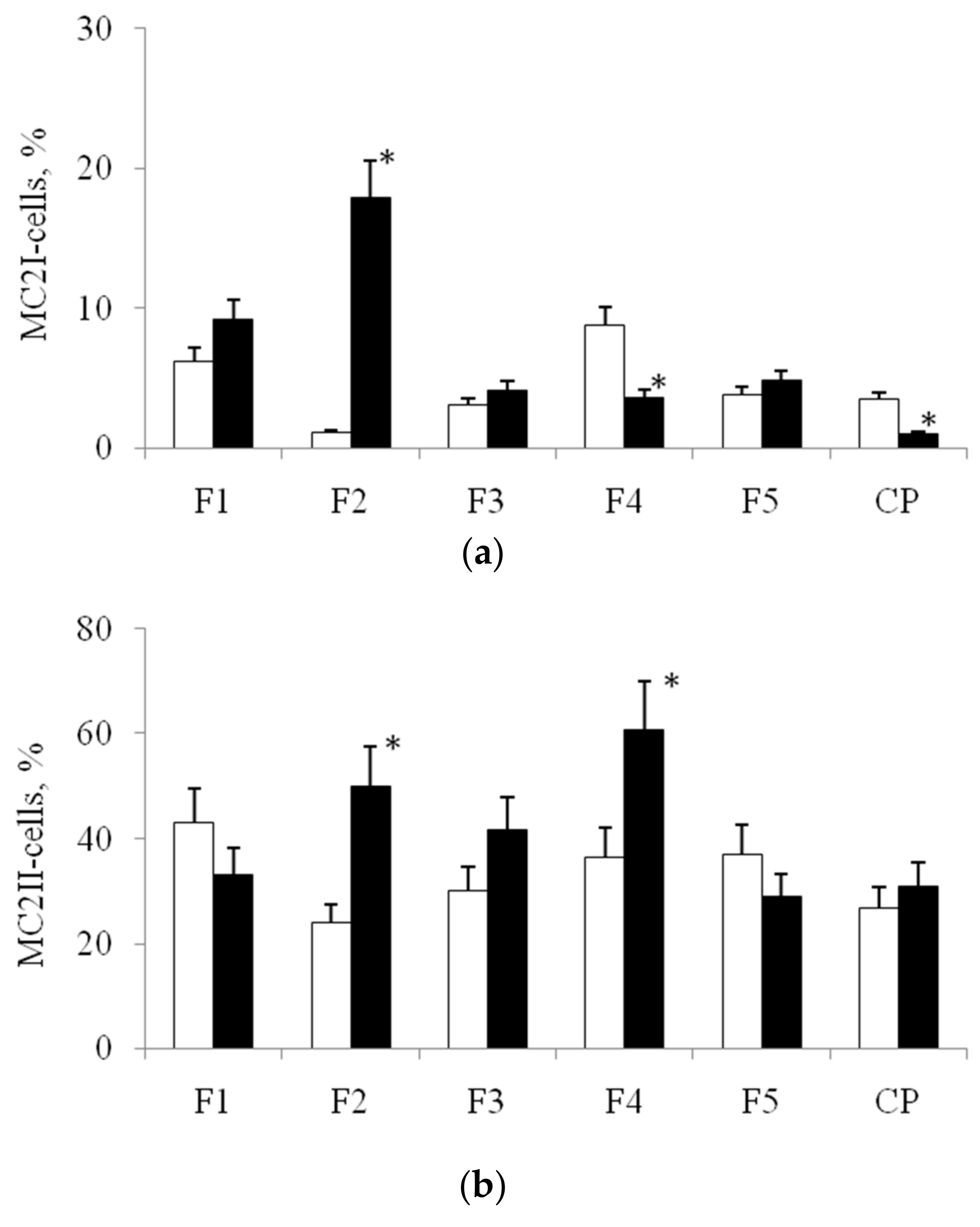
3.2. Expression of Marker CD Antigens on MNs, Isolated from Blood before and after Stenting
3.3. The Morphology of MN-MPhs on PHA Bionanofilm Samples with Different Surface Reliefs
3.4. The Concentration of IL-6 and IL-10 in the MPh Culture Filtrate
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Loye, A.M.; Kinser, E.R.; Bensouda, S.; Shayan, M.; Davis, R.; Wang, R.; Chen, Z.; Schwarz, U.D.; Schroers, J.; Kyriakides, T.R. Regulation of Mesenchymal Stem Cell Differentiation by Nanopatterning of Bulk Metallic Glass. Sci. Rep. 2018, 8, 8758. [Google Scholar] [CrossRef] [PubMed]
- Su, N.; Gao, P.-L.; Wang, K.; Wang, J.-Y.; Zhong, Y.; Luo, Y. Fibrous scaffolds potentiate the paracrine function of mesenchymal stem cells: A new dimension in cell-material interaction. Biomaterials 2017, 141, 74–85. [Google Scholar] [CrossRef] [PubMed]
- De Peppo, G.M.; Agheli, H.; Karlsson, C.; Ekström, K.; Brisby, H.; Lennerås, M.; Gustafsson, S.; Sjövall, P.; Johansson, A.; Olsson, E.; et al. Osteogenic response of human mesenchymal stem cells to well-defined nanoscale topography in vitro. Int. J. Nanomed. 2014, 9, 2499–2515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Bauer, S.; Schlegel, K.A.; Neukam, F.W.; Von Der Mark, K.; Schmuki, P. TiO2Nanotube Surfaces: 15 nm-An Optimal Length Scale of Surface Topography for Cell Adhesion and Differentiation. Small 2009, 5, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Kato, E.; Yamamoto, A.; Sakurai, K. A titanium surface with nano-ordered spikes and pores enhances human dermal fibroblastic extracellular matrix production and integration of collagen fibers. Biomed. Mater. 2016, 11, 15010. [Google Scholar] [CrossRef]
- Schvartzman, M.; Palma, M.; Sable, J.; Abramson, J.; Hu, X.; Sheetz, M.P.; Wind, S.J. Nanolithographic Control of the Spatial Organization of Cellular Adhesion Receptors at the Single-Molecule Level. Nano Lett. 2011, 11, 1306–1312. [Google Scholar] [CrossRef] [Green Version]
- Khang, N. Real time macrophage migration analysis and associated pro-inflammatory cytokine release on transparent carbon nanotube/polymer composite nano-film. Nanotechnology 2015, 26, 325101. [Google Scholar] [CrossRef]
- Huang, J.; Grater, S.V.; Corbellini, F.; Rinck, S.; Bock, E.; Kemkemer, R.; Kessler, H.; Ding, J.; Spatz, J.P.; Rinck-Jahnke, S. Impact of Order and Disorder in RGD Nanopatterns on Cell Adhesion. Nano Lett. 2009, 9, 1111–1116. [Google Scholar] [CrossRef] [Green Version]
- Hulshof, F.F.; Zhao, Y.; Vasilevich, A.; Beijer, N.R.; De Boer, M.; Papenburg, B.J.; Van Blitterswijk, C.; Stamatialis, D.; De Boer, J. NanoTopoChip: High-throughput nanotopographical cell instruction. Acta Biomater. 2017, 62, 188–198. [Google Scholar] [CrossRef]
- Treiser, M.D.; Yang, E.H.; Gordonov, S.; Cohen, D.M.; Androulakis, I.P.; Kohn, J.; Chen, C.S.; Moghe, P.V. Cytoskeleton-based forecasting of stem cell lineage fates. Proc. Natl. Acad. Sci. USA 2010, 107, 610–615. [Google Scholar] [CrossRef] [Green Version]
- Maniotis, A.J.; Chen, C.S.; Ingber, D.E. Demonstration of mechanical connections between integrins, cytoskeletal filaments, and nucleoplasm that stabilize nuclear structure. Proc. Natl. Acad. Sci. USA 1997, 94, 849–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, E.H.; Kim, Y.; Kshitiz; An, S.S.; Ahn, E.H.; Afzal, J.; Lee, S.; Kwak, M.; Suh, K.-Y.; Kim, D.-H.; et al. Spatial control of adult stem cell fate using nanotopographic cues. Biomaterials 2014, 35, 2401–2410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volova, T.G.; Vinogradova, O.N.; Zhila, N.O.; Kiselev, E.G.; Peterson, I.V.; Vasil’Ev, A.D.; Sukovatyi, A.G.; Shishatskaya, E.I. Physicochemical properties of multicomponent polyhydroxyalkanoates: Novel aspects. Polym. Sci. Ser. A 2017, 59, 98–106. [Google Scholar] [CrossRef]
- Berg, K.E.; Ljungcrantz, I.; Andersson, L.; Bryngelsson, C.; Hedblad, B.; Fredrikson, G.N.; Nilsson, J.; Björkbacka, H. CD14++CD16- monocytes predict cardiovascular events. Circ. Cardiovasc. Genet. 2012, 5, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Carlin, L.M.; Stamatiades, E.G.; Auffray, C.; Hanna, R.N.; Glover, L.; Vizcay-Barrena, G.; Hedrick, C.C.; Cook, H.T.; Diebold, S.; Geissmann, F. Nr4a1-dependent Ly6C(low) monocytes monitor endothelial cells and orchestrate their disposal. Cell 2013, 153, 362–375. [Google Scholar] [CrossRef] [Green Version]
- Cros, J.; Cagnard, N.; Woollard, K.; Patey, N.; Zhang, S.-Y.; Senechal, B.; Puel, A.; Biswas, S.K.; Moshous, D.; Picard, C.; et al. Human CD14dim monocytes patrol and sense nucleic acids and viruses via TLR7 and TLR8 receptors. Immunity 2010, 33, 375–386. [Google Scholar] [CrossRef] [Green Version]
- Hanna, R.N.; Cekic, C.; Sag, D.; Tacke, R.; Thomas, G.D.; Nowyhed, H.; Herrley, E.; Rasquinha, N.; McArdle, S.; Wu, R.; et al. Patrolling monocytes control tumor metastasis to the lung. Science 2015, 350, 985–990. [Google Scholar] [CrossRef] [Green Version]
- Rossol, M.; Kraus, S.; Pierer, M.; Baerwald, C.; Wagner, U. The CD14(bright) CD16+ monocyte subset is expanded in rheumatoid arthritis and promotes expansion of the Th17 cell population. Arthritis Rheum. 2012, 64, 671–677. [Google Scholar] [CrossRef]
- Rogacev, K.S.; Cremers, B.; Zawada, A.M.; Seiler, S.; Binder, N.; Ege, P.; Grobe-Dunker, G.; Heisel, I.; Hornof, F.; Jeken, J.; et al. CD14++CD16+ monocytes independently predict cardiovascular events: A cohort study of 951 patients referred for elective coronary angiography. J. Am. Coll. Cardiol. 2012, 60, 1512–1520. [Google Scholar] [CrossRef] [Green Version]
- Urra, X.; Villamor, N.; Amaro, S.; Gómez-Choco, M.; Obach, V.; Oleaga, L.; Planas, A.M.; Chamorro, A. Monocyte Subtypes Predict Clinical Course and Prognosis in Human Stroke. J. Pharmacol. 2009, 29, 994–1002. [Google Scholar] [CrossRef] [Green Version]
- Azeredo, E.L.; Neves-Souza, P.C.; Alvarenga, A.R.; Reis, S.R.N.I.; Torrentes-Carvalho, A.; Zagne, S.O.; Nogueira, R.M.R.; Oliveira-Pinto, L.M.; Kubelka, C.F. Differential regulation of toll-like receptor-2, toll-like receptor-4, CD16 and human leucocyte antigen-DR on peripheral blood monocytes during mild and severe dengue fever. Immunology 2010, 130, 202–216. [Google Scholar] [CrossRef] [PubMed]
- Hamers, A.A.; Dinh, H.Q.; Thomas, G.D.; Marcovecchio, P.; Blatchley, A.; Nakao, C.S.; Kim, C.; McSkimming, C.; Taylor, A.M.; Nguyen, A.T.; et al. Human Monocyte Heterogeneity as Revealed by High-Dimensional Mass Cytometry. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Abcam. Available online: https://docs.abcam.com/pdf/immunology/Guide-to-human-CD-antigens.pdf (accessed on 30 December 2019).
- Yang, J.; Zhang, L.; Yu, C.; Yang, X.-F.; Wang, H. Monocyte and macrophage differentiation: Circulation inflammatory monocyte as biomarker for inflammatory diseases. Biomark. Res. 2014, 2, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wildgruber, M.; Aschenbrenner, T.; Wendorff, H.; Czubba, M.; Glinzer, A.; Haller, B.; Schiemann, M.; Zimmermann, A.; Berger, H.; Eckstein, H.H.; et al. The “intermediate” CD14++CD16+ monocyte subset increases in severe peripheral artery disease in humans. Sci. Rep. 2016, 6, 39483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.S.; Stachelek, S.J.; Tomczyk, N.; Finley, M.J.; Composto, R.J.; Eckmann, D.M. Correlating macrophage morphology and cytokine pro¬duction resulting from biomaterial contact. J. Biomed. Mater. Res. A 2013, 101, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Hind, L.E.; Dembo, M.; Hammer, D.A. Macrophage motility is driven by frontal-towing with a force magnitude dependent on substrate stiffness. Integr. Boil. 2015, 7, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Webster, T.J. Reduced immune cell responses on nano and submicron rough titanium. Acta Biomater. 2015, 16, 223–231. [Google Scholar] [CrossRef]
- Collison, J.L.; Carlin, L.M.; Eichmann, M.; Geissmann, F.; Peakman, M. Heterogeneity in the Locomotory Behavior of Human Monocyte Subsets over Human Vascular Endothelium In Vitro. J. Immunol. 2015, 195, 1162–1170. [Google Scholar] [CrossRef]
- Van Goethem, E.; Poincloux, R.; Gauffre, F.; Maridonneau-Parini, I.; Le Cabec, V. Matrix architecture dictates three-dimensional migration modes of human macrophages: Differential involvement of proteases and podosome-like structures. J. Immunol. 2010, 184, 1049–1061. [Google Scholar] [CrossRef]
- Yokose, S.; Klokkevold, P.R.; Takei, H.H.; Kadokura, H.; Kikui, T.; Hibino, Y.; Shigeta, H.; Nakajima, H.; Kawazu, H. Effects of surface microtopography of titanium disks on cell proliferation and differentiation of osteoblast-like cells isolated from rat calvariae. Dent. Mater. J. 2018, 37, 272–277. [Google Scholar] [CrossRef] [Green Version]
- Czeisler, C.; Short, A.; Nelson, T.; Gygli, P.; Ortiz, C.; Catacutan, F.P.; Stocker, B.; Cronin, J.; Lannutti, J.; Winter, J.; et al. Surface topography during neural stem cell differentiation regulates cell migration and cell morphology. J. Comp. Neurol. 2016, 524, 3485–3502. [Google Scholar] [CrossRef] [PubMed]
- Hulsman, M.; Hulshof, F.; Unadkat, H.; Papenburg, B.J.; Stamatialis, D.F.; Truckenmüller, R.; Van Blitterswijk, C.; De Boer, J.; Reinders, M.J.; Stamatialis, D. Analysis of high-throughput screening reveals the effect of surface topographies on cellular morphology. Acta Biomater. 2015, 15, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Bruce, A.; Mezan, R.; Kadiyala, A.; Wang, L.; Dawson, J.; Rojanasakul, Y.; Yang, Y. Nanotopographical Modulation of Cell Function through Nuclear Deformation. ACS Appl. Mater. Interfaces 2016, 8, 5082–5092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, L.; Zhao, W.; Lou, H.-Y.; Lin, Z.C.; Lee, S.W.; Chowdary, P.; Cui, Y.; Cui, B. Vertical nanopillars for in situ probing of nuclear mechanics in adherent cells. Nat. Nanotechnol. 2015, 10, 554–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fais, S.; Burgio, V.L.; Silvestri, M.; Capobianchi, M.R.; Pacchiarotti, A.; Pallone, F. Multinucleated giant cells generation induced by interferon-gamma. Changes in the expression and distribution of the intercellular adhesion molecule-1 during macrophages fusion and multinucleated giant cell formation. Lab. Investig. 1994, 71, 737–744. [Google Scholar] [PubMed]
- Helming, L.; Winter, J.; Gordon, S. The scavenger receptor CD36 plays a role in cytokine-induced macrophage fusion. J. Cell Sci. 2009, 122, 453–459. [Google Scholar] [CrossRef] [Green Version]
- Miron, R.J.; Bosshardt, D.D. Multinucleated giant cells: Good guys or bad guys. Tissue Eng. Part B Rev. 2017, 24, 1–37. [Google Scholar]
- Rey-Giraud, F.; Hafner, M.; Ries, C.H. In Vitro Generation of Monocyte-Derived Macrophages under Serum-Free Conditions Improves Their Tumor Promoting Functions. PLoS ONE 2012, 7, 42656. [Google Scholar] [CrossRef] [Green Version]
- Heinrich, F.; Lehmbecker, A.; Raddatz, B.B.; Kegler, K.; Tipold, A.; Stein, V.M.; Kalkuhl, A.; Deschl, U.; Baumgärtner, W.; Ulrich, R.; et al. Morphologic, phenotypic, and transcriptomic characterization of classically and alternatively activated canine blood-derived macrophages in vitro. PLoS ONE 2017, 12, e0183572. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CDAG | Expression | Functions CDAG |
|---|---|---|
| CD14 | MNs, MPhs | Receptor for complex of lipopolysaccharides and lipopolysaccharide binding protein |
| CD16 | MPhs | Low affinity Fc receptor. Mediates phagocytosis and the antibody-dependent cellular cytotoxicity. |
| CD31 | MNs | Cell adhesion. Plays a key role in leukocyte trafficking across the endothelium. |
| CD36 | MNs, MPhs | Scavenger receptor preferentially found within lipid rafts. Mediatesadhesion and phagocytosis |
| CD62L | MNs | Leukocyte rolling and homing on activated endothelium. |
| CD68 | MNs, MPhs | Possibly has a role in phagocytic activities of tissue macrophages, both in intracellular lysosomal metabolism and extracellular cell-cell and cell-pathogen interactions. Binds to tissue- and organ-specific lectins or selectins, allowing homing of macrophage subsets to particular sites. May allow macrophages to crawl over selectin bearing substrates or other cells. |
| CD163 | MNs, MPhs | Clearance and endocytosis of hemoglobin/haptoglobin complexes by macrophages. |
| CD206 | MNs, MPhs | Endocytosis (phagocytosis and pinocytosis) of mannose-containing solutes. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menzyanova, N.G.; Pyatina, S.A.; Shabanov, A.V.; Nemtsev, I.V.; Stolyarov, D.P.; Dryganov, D.B.; Sakhnov, E.V.; Shishatskaya, E.I. The Morphology and Phenotype of Monocyte-Macrophages When Cultured on Bionanofilms Substrates with Different Surface Relief Profiles. Biomolecules 2020, 10, 65. https://doi.org/10.3390/biom10010065
Menzyanova NG, Pyatina SA, Shabanov AV, Nemtsev IV, Stolyarov DP, Dryganov DB, Sakhnov EV, Shishatskaya EI. The Morphology and Phenotype of Monocyte-Macrophages When Cultured on Bionanofilms Substrates with Different Surface Relief Profiles. Biomolecules. 2020; 10(1):65. https://doi.org/10.3390/biom10010065
Chicago/Turabian StyleMenzyanova, Natalia G., Svetlana A. Pyatina, Alexander V. Shabanov, Ivan V. Nemtsev, Dmitry P. Stolyarov, Dmitry B. Dryganov, Eugene V. Sakhnov, and Ekaterina I. Shishatskaya. 2020. "The Morphology and Phenotype of Monocyte-Macrophages When Cultured on Bionanofilms Substrates with Different Surface Relief Profiles" Biomolecules 10, no. 1: 65. https://doi.org/10.3390/biom10010065
APA StyleMenzyanova, N. G., Pyatina, S. A., Shabanov, A. V., Nemtsev, I. V., Stolyarov, D. P., Dryganov, D. B., Sakhnov, E. V., & Shishatskaya, E. I. (2020). The Morphology and Phenotype of Monocyte-Macrophages When Cultured on Bionanofilms Substrates with Different Surface Relief Profiles. Biomolecules, 10(1), 65. https://doi.org/10.3390/biom10010065