Molecular Dynamics Simulations Predict that rSNP Located in the HNF-1α Gene Promotor Region Linked with MODY3 and Hepatocellular Carcinoma Promotes Stronger Binding of the HNF-4α Transcription Factor




Abstract
:1. Introduction
2. Methods
2.1. Preparation of the Starting Structures for the MD Simulation
2.2. Molecular Dynamics Simulations
2.3. Analysis
2.3.1. Analysis of Intermolecular Interactions
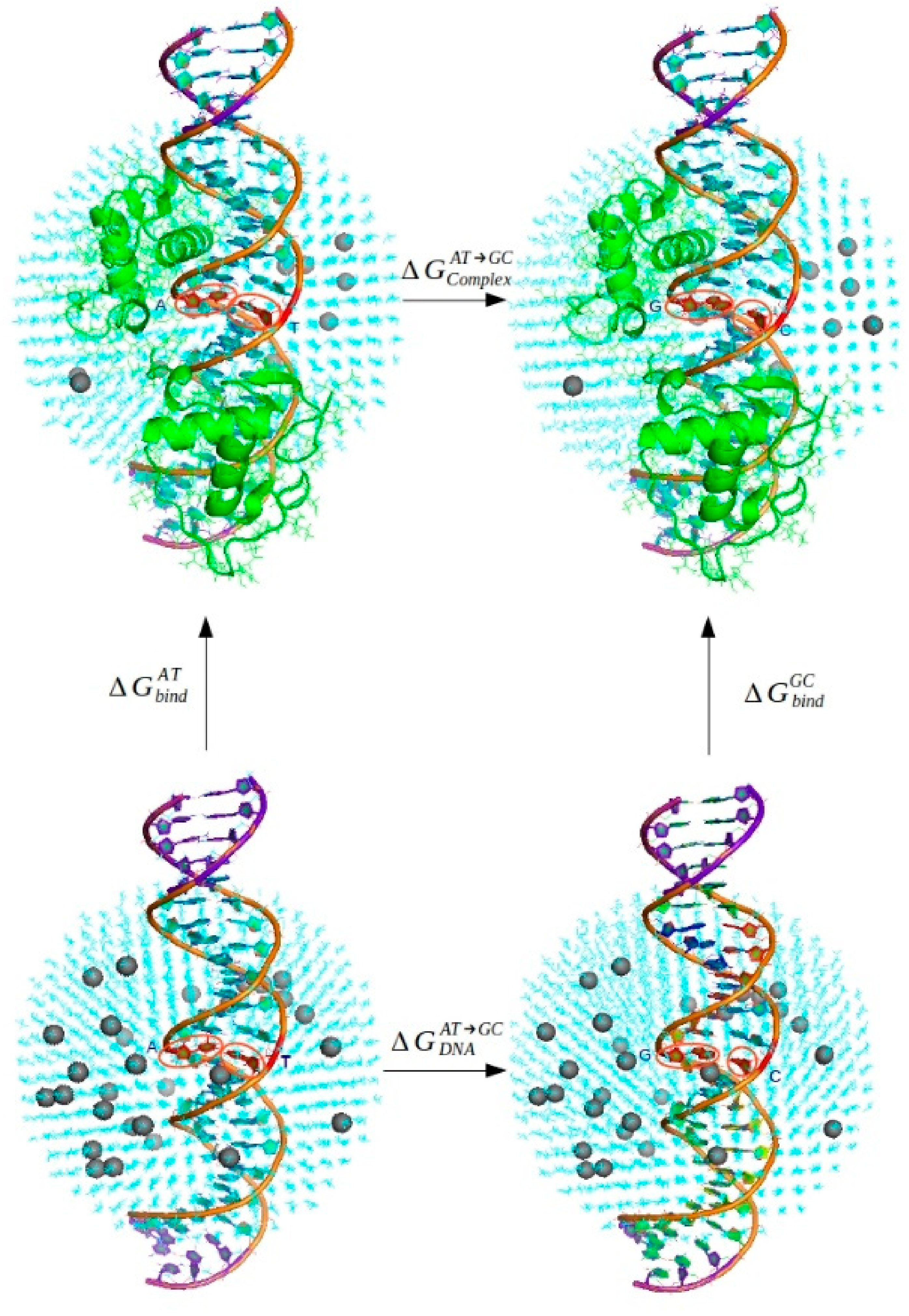
2.3.2. Thermodynamic Analysis
- the binding free energy (kcal/mol);
- average vdW interaction energies between the ligand and it’s surrounding (kcal/mol);
- average electrostatic interaction energies between the ligand and it’s surrounding (kcal/mol);
- l ligand;
- s surrounding environment;
- α, β empirical parameters of the LIE method.
- the binding free energy (kcal/mol);
- the difference between the binding free energy of the wild-type complex and the mutated rSNP rs35126805 containing complex (kcal/mol);
- average van der Waals interaction energies between the ligand and it’s surrounding (kcal/mol);
- average electrostatic interaction energies between the ligand and it’s surrounding (kcal/mol);
- l ligand: AT—wild-type base pair, GC—mutated rSNP base pair; Complex—complex of HNF-4α transcription factor bound to the DNA, DNA—solely the DNA double helix;
- s surrounding environment included in the water solvent sphere;
- α, β empirical parameters of the LIE method.
3. Results and Discussion
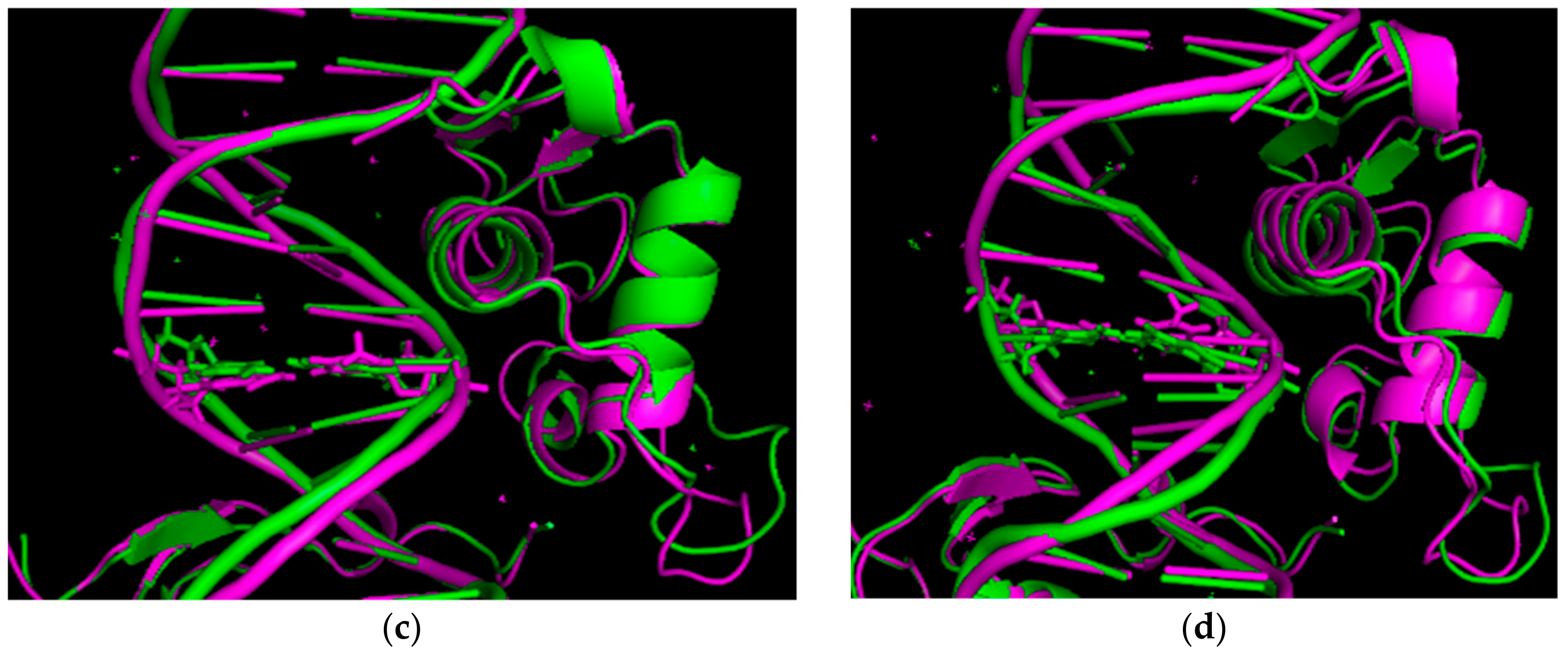
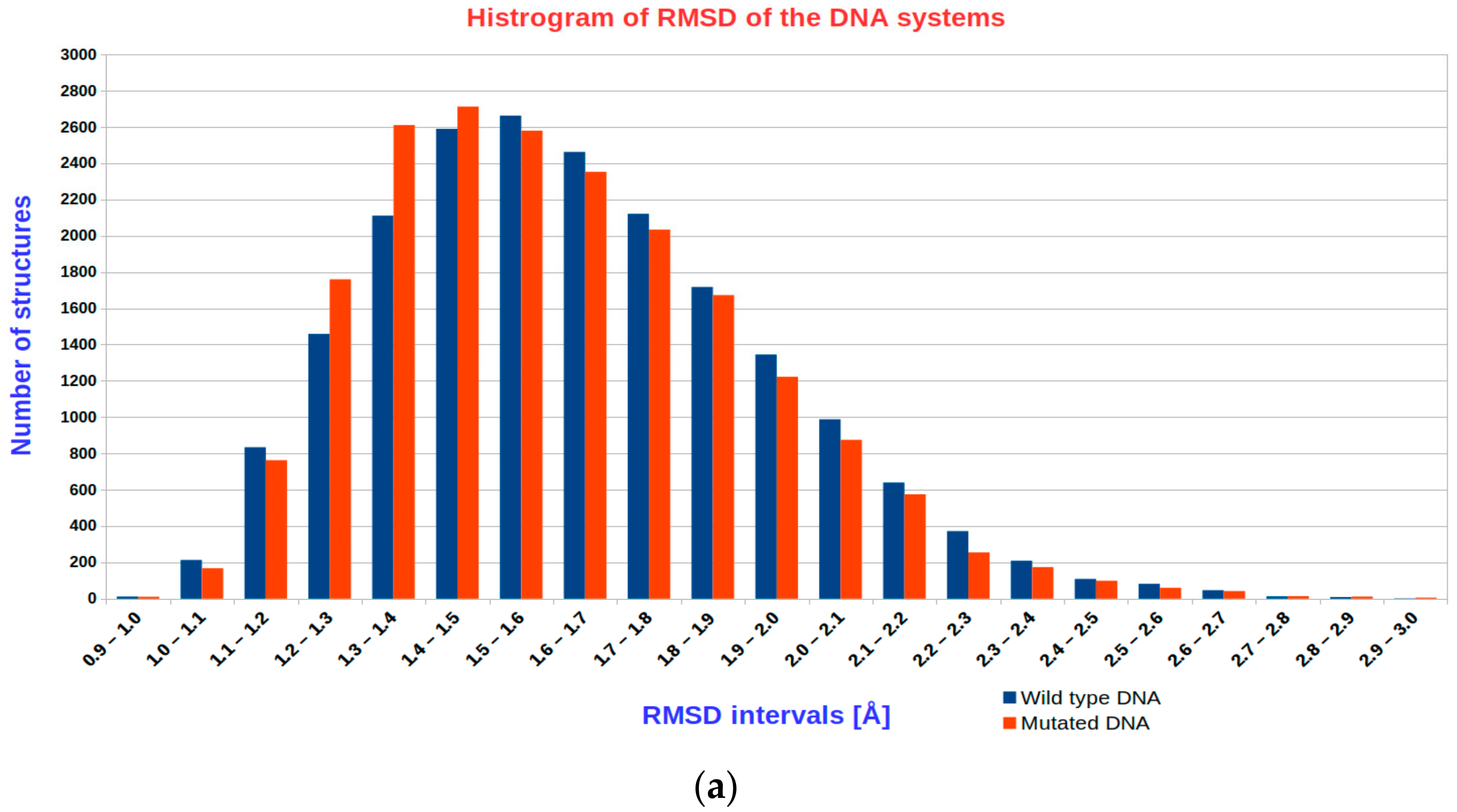
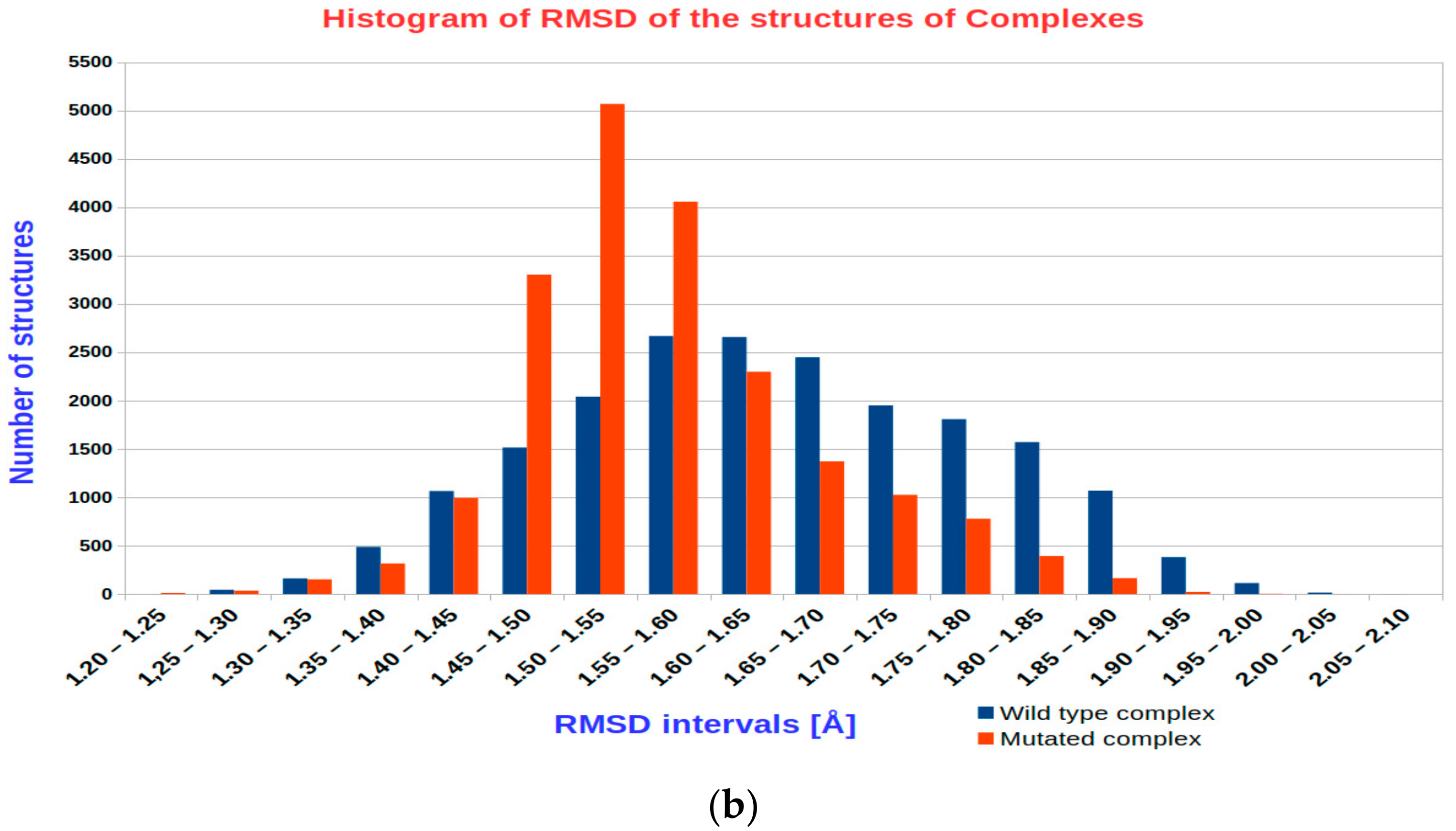
3.1. The Results of RMSD Analysis
3.2. Analysis of Intermolecular Interactions—Hydrogen Bonds
3.3. The Results of the Thermodynamical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Coot | Crystallographic Object-Oriented Toolkit |
| Cys | cysteine |
| GWAS | genome-wide association studies |
| HNF | hepatocyte nuclear factor |
| HNF-1α | hepatocyte nuclear factor 1 alpha |
| HNF-4α | hepatocyte nuclear factor 4 alpha |
| LIE | Linear Interaction Energy method |
| MODY3 | Maturity-onset diabetes of the young 3 |
| MD | molecular dynamics |
| PDB | Protein Data Bank |
| RMSD | root-mean-square deviation |
| rSNP | regulatory single nucleotide polymorphism |
| SNP | single nucleotide polymorphism |
References
- Fagny, M.; Platig, J.; Kuijjer, M.L.; Xihong Lin, X.; Quackenbush, J. Nongenic cancer-risk SNPs affect oncogenes, tumour-suppressor genes, and immune function. Br. J. Cancer 2020, 122, 569–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalia, N.; Sharma, A.; Kaur, M.; Kamboj, S.S.; Singh, J. A comprehensive in silico analysis of non-synonymous and regulatory SNPs of human MBL2 gene. Springerplus 2016, 5, 811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Liu, M.; Li, B.; Wang, Y.; Yue, J.; Liang, L.; Sun, J. Exploring the mechanism of regulatory SNP of KLK3 by molecular dynamics simulation. J. Biomol. Struct. Dyn. 2013, 31, 426–440. [Google Scholar] [CrossRef] [PubMed]
- Sud, A.; Kinnersley, B.; Houlston, R.S. Genome-wide association studies of cancer: Current insights and future perspectives. Nat. Rev. Cancer 2017, 17, 692–704. [Google Scholar] [CrossRef]
- Gragnoli, C.; Lindner, T.; Cockburn, B.N.; Kaisaki, P.J.; Gragnoli, F.; Marozzi, G.; Bell, G.I. Maturity-Onset Diabetes of the Young Due to a Mutation in the Hepatocyte Nuclear Factor-4α Binding Site in the Promoter of the Hepatocyte Nuclear Factor-1α Gene. Diabetes 1997, 46, 1648–1651. [Google Scholar] [CrossRef]
- Pihoker, C.; Gilliam, L.K.; Ellard, S.; Dabelea, D.; Davis, C.; Dolan, L.M.; Greenbaum, C.J.; Imperatore, G.; Lawrence, J.M.; Marcovina, S.M. Prevalence, characteristics and clinical diagnosis of maturity onset diabetes of the young due to mutations in HNF1A, HNF4A, and glucokinase: Results from the SEARCH for Diabetes in Youth. J. Clin. Endocrinol. Metab. 2013, 98, 4055–4062. Available online: https://www.niddk.nih.gov/health-information/diabetes/overview/what-is-diabetes/monogenic-neonatal-mellitus-mody (accessed on 13 March 2019).
- Hattersley, A.T.; Pearson, E.R. Minireview: Pharmacogenetics and beyond: The interaction of therapeutic response, beta-cell physiology, and genetics in diabetes. Endocrinology 2006, 147, 2657–2663. [Google Scholar] [CrossRef] [Green Version]
- Sel, S.; Ebert, T.; Ryffel, G.U.; Drewes, T. Human renal cell carcinogenesis is accompanied by a coordinate loss of the tissue specific transcription factors HNF4 alpha and HNF1 alpha. Cancer Lett. 1996, 101, 205–210. [Google Scholar] [CrossRef]
- Azmi, A.S.; Bao, G.F.; Gao, J.; Mohammad, R.M.; Sarkar, F.H. Network Insights into the Genes Regulated by Hepatocyte Nuclear Factor 4 in Response to Drug induced Perturbations. Curr. Drug. Discov. Technol. 2013, 10, 147–154. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information, U.S. National Library of Medicine. HNF1A. Available online: https://www.ncbi.nlm.nih.gov/gene/6927 (accessed on 14 March 2019).
- Maestro, M.A.; Cardalda, C.; Boj, S.F.; Luco, R.F.; Servitja, J.M.; Ferrer, J. Distinct roles of HNF1beta, HNF1alpha, and HNF4alpha in regulating pancreas development, beta-cell function and growth. Endocr. Dev. 2007, 12, 33–45. [Google Scholar] [CrossRef]
- Abel, E.V.; Goto, M.; Magnuson, B.; Abraham, S.; Ramanathan, N.; Hotaling, E.; Alaniz, A.A.; Kumar-Sinha, C.; Dziubinski, M.L.; Urs, S.; et al. HNF1A is a novel oncogene that regulates human pancreatic cancer stem cell properties. Elife 2018, 7, e33947. [Google Scholar] [CrossRef]
- Luo, Z.; Li, Y.; Wang, H.; Fleming, J.; Li, M.; Kang, Y.; Zhang, R.; Li, D. Hepatocyte Nuclear Factor 1A (HNF1A) as a Possible Tumor Suppressor in Pancreatic Cancer. PLoS ONE 2015, 10, e0121082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Center for Biotechnology Information, U.S. National Library of Medicine. HNF4A. Available online: https://www.ncbi.nlm.nih.gov/gene/3172 (accessed on 14 March 2019).
- Hanawa, M.; Takayama, K.; Sakurai, F.; Tachibana, M.; Mizuguchi, H. Hepatocyte Nuclear Factor 4 Alpha Promotes Definitive Endoderm Differentiation from Human Induced Pluripotent Stem Cells. Stem. Cell. Rev. Rep. 2017, 13, 542–551. [Google Scholar] [CrossRef]
- Yan, H.; Wang, Q.; Shen, Q.; Li, Z.; Tian, J.; Jiang, Q.; Gao, L. Identification of potential transcription factors, long noncoding RNAs, and microRNAs associated with hepatocellular carcinoma. J. Cancer Res. Ther. 2018, 14, S622–S627. [Google Scholar] [CrossRef]
- Mura, C.; McCammon, J.A. Molecular dynamics of a κB DNA element: Base flipping via cross-strand intercalative stacking in a microsecond-scale simulation. Nucleic Acids Res. 2008, 36, 4941–4955. [Google Scholar] [CrossRef] [PubMed]
- Marco, E.; García-Nieto, R.; Gago, F. Assessment by Molecular Dynamics Simulations of the Structural Determinants of DNA-binding Specificity for Transcription Factor Sp1. J. Mol. Biol. 2003, 328, 9–32. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Sun, T.; Pei, J.; Ouyang, Q. Mutation-induced protein interaction kinetics changes affect apoptotic network dynamic properties and facilitate oncogenesis. Proc. Natl. Acad. Sci. USA 2015, 112, E4046–E4054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.; Tan, S.; Fang, D.; Zhang, R.; Zhou, S.; Wu, W.; Zheng, K. Probing the binding mechanism of novel dual NF-κB/AP-1 inhibitors by 3D_QSAR, docking and molecular dynamics simulations. RSC Adv. 2015, 5, 81523–81532. [Google Scholar] [CrossRef]
- Klvaňa, M.; Bren, U.; Florián, J. Uniform Free-Energy Profiles of the P–O Bond Formation and Cleavage Reactions Catalyzed by DNA Polymerases β and λ. J. Phys. Chem. B 2016, 120, 13017–13030. [Google Scholar] [CrossRef]
- Martínek, V.; Bren, U.; Goodman, M.F.; Warshel, A.; Florián, J. DNA polymerase β catalytic efficiency mirrors the Asn279–dCTP H-bonding strength. Febs. Lett. 2007, 581, 775–780. [Google Scholar] [CrossRef] [Green Version]
- Fenollar-Ferrer, C.; Anselmi, C.; Carnevale, V.; Raugei, S.; Carloni, P. Insights on the acetylated NF-kB transcription factor complex with DNA from molecular dynamics simulations. Proteins 2012, 80, 1560–1568. [Google Scholar] [CrossRef]
- Dutta, S.; Agrawal, Y.; Mishra, A.; Kaur Dhanjal, J.; Sundar, D. A theoretical investigation of DNA dynamics and desolvation kinetics for zinc finger proteinZif268. BMC Genom. 2015, 16 (Suppl. 12), S5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouard, C.; Terreux, R.; Honorat, M.; Manship, B.; Ansieau, S.; Vigneron, A.M.; Puisieux, A.; Payen, L. Deciphering the molecular mechanisms underlying the binding of the TWIST1/E12 complex to regulatory E-box sequences. Nucleic. Acids. Res. 2016, 44, 5470–5489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacKerell, A.D.; Nilsson, L. Molecular dynamics simulations of nucleic acid–protein complexes. Curr. Opin. Struct. Biol. 2008, 18, 194–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George Priya Doss, C.; Nagasundaram, N.; Chakraborty, C.; Chen, L.; Zhu, H. Extrapolating the effect of deleterious nsSNPs in the binding adaptability of flavopiridol with CDK7 protein: A molecular dynamics approach. Hum. Genom. 2013, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aken, B.L.; Ayling, S.; Barrell, D.; Clarke, L.; Curwen, V.; Fairley, S.; Banet, J.F.; Billis, K.; Girón, C.G.; Hourlier, T.; et al. The Ensembl gene annotation system. Database 2016, 2016, baw093. Available online: https://www.ensembl.org/index.html (accessed on 15 July 2016). [CrossRef]
- National Center for Biotechnology Information, U.S. National Library of Medicine. dbSNP. Available online: https://www.ncbi.nlm.nih.gov/SNP/ (accessed on 13 March 2019).
- Marelius, J.; Kolmodin, K.; Feierberg, I.; Åqvist, J.Q. A molecular dynamics program for free energy calculations and empirical valence bond simulations in biomolecular systems. J. Mol. Graph. Model. 1998, 16, 213–225. Available online: http://xray.bmc.uu.se/aqwww/Q (accessed on 1 January 2004). [CrossRef]
- Hoops, S.C.; Anderson, K.W.; Merz, K.M., Jr. Force Field Design for Metalloproteins. J. Am. Chem. Soc. 1991, 113, 8262–8270. [Google Scholar] [CrossRef]
- Peters, M.B.; Yang, Y.; Wang, B.; Füsti-Molnár, L.; Weaver, M.N.; Merz, K.M., Jr. Structural Survey of Zinc-Containing Proteins and Development of the Zinc AMBER Force Field (ZAFF). J. Chem. Theory Comput. 2010, 6, 2935–2947. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and Development of Coot. Acta Crystallogr. D 2010, 66, 486–501. Available online: https://www2.mrc-lmb.cam.ac.uk/personal/pemsley/coot/ (accessed on 18 December 2020). [CrossRef] [Green Version]
- The PyMOL Molecular Graphics System, Version 2.0 Schrödinger, LLC. Available online: https://pymol.org/2/ (accessed on 18 December 2020).
- King, G.; Warshel, A. A surface constrained all-atom solvent model for effective simulations of polar solutions. J. Chem. Phys. 1989, 91, 3647. [Google Scholar] [CrossRef]
- Sham, Y.Y.; Warshel, A. The surface constraint all atom model provides size independent results in calculations of hydration free energies. J. Chem. Phys. 1998, 109, 7940. [Google Scholar] [CrossRef]
- Bren, U.; Lah, J.; Bren, M.; Martínek, V.; Jan Florián, J. DNA Duplex Stability: The Role of Preorganized Electrostatics. J. Phys. Chem. B 2010, 114, 2876–2885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.-J.; Olson, W.K. 3DNA: A software package for the analysis, rebuilding and visualization of three-dimensional nucleic acid structures. Nucleic Acids Res. 2003, 31, 5108–5121, The 3DNA. Available online: https://x3dna.org/ (accessed on 18 December 2020). [CrossRef] [PubMed] [Green Version]
- Florián, J.; Goodman, M.F.; Warshel, A. Free-Energy Perturbation Calculations of DNA Destabilization by Base Substitutions: The Effect of Neutral Guanine·Thymine, Adenine·Cytosine and Adenine·Difluorotoluene Mismatches. J. Phys. Chem. B 2000, 104, 10092–10099. [Google Scholar] [CrossRef]
- Cheatham, T.E., III; Kollman, P.A. Molecular Dynamics Simulation of Nucleic Acids. Annu. Rev. Physchem. 2000, 51, 435–471. [Google Scholar] [CrossRef]
- Ryckaert, J.P.; Ciccotti, G.; Berendsen, H.J.C. Numerical integration of the cartesian equations of motion of a system with constraints: Molecular dynamics of n-alkanes. J. Comp. Phys. 1977, 23, 327–341. [Google Scholar] [CrossRef] [Green Version]
- Lee, F.S.; Warshel, A. A local reaction field method for fast evaluation of long-range electrostatic interactions in molecular simulations. J. Chem. Phys. 1992, 97, 3100. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD—Visual Molecular Dynamics. J. Mol. Graph. 1996, 14, 33–38. Available online: http://www.ks.uiuc.edu/Research/vmd/ (accessed on 18 December 2020). [CrossRef]
- Hansson, T.; Marelius, J.; Åqvist, J. Ligand binding affinity prediction by linear interaction energy methods. J. Comput. Aided Mol. Des. 1998, 12, 27–35. [Google Scholar] [CrossRef]
- Klvaňa, M.; Bren, U. Aflatoxin B1-Formamidopyrimidine DNA adducts: Relationships between structures, free energies, and melting temperatures. Molecules 2019, 24, 150. [Google Scholar] [CrossRef] [Green Version]
- Mikulskis, P.; Genheden, S.; Rydberg, P.; Sandberg, L.; Olsen, L.; Ryde, U. Binding affinities in the SAMPL3 trypsin and host–guest blind tests estimated with the MM/PBSA and LIE methods. J. Comput. Aided Mol. Des. 2012, 26, 527–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- König, G.; Brooks, B.R. Predicting binding affinities of host-guest systems in the SAMPL3 blind challenge: The performance of relative free energy calculations. J. Comput. Aided Mol. Des. 2012, 26, 543–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan-Yao-Chong, M.; Marsin, S.; Quevillon-Cheruel, S.; Durand, D.; Ha-Duong, T. Structural ensemble and biological activity of DciA intrinsically disordered region. J. Struct. Biol. 2020, 212, 107573. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.E.; van Gunsteren, W.F. When Are Free Energy Components Meaningful? J. Phys. Chem. 1994, 98, 13735–13740. [Google Scholar] [CrossRef]
- Galindo-Murillo, R.; Robertson, J.C.; Zgarbová, M.; Šponer, J.; Otyepka, M.; Jurečka, P.; Cheatham, T.E., III. Assessing the Current State of Amber Force Field Modifications for DNA. J. Chem. Theory Comput. 2016, 12, 4114–4127. [Google Scholar] [CrossRef] [PubMed]
- Zgarbová, M.; Luque, F.J.; Šponer, J.; Cheatham III, T.E.; Otyepka, M.; Jurečka, P. Toward Improved Description of DNA Backbone: Revisiting Epsilon and Zeta Torsion Force Field Parameters. J. Chem. Theory Comput. 2013, 9, 2339–2354. [Google Scholar] [CrossRef] [PubMed]
- Ivani, I.; Dans, P.D.; Noy, A.; Pérez, A.; Faustino, I.; Hospital, A.; Walther, J.; Andrio, P.; Goñi, R.; Balaceanu, A.; et al. PARMBSC1: A Refined Force-Field for DNA Simulations. Nat. Methods 2016, 13, 55–58. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Production Run | [kcal/mol] | ||
|---|---|---|---|
| Production run 1 | −0.20 | −0.44 | −0.64 |
| Production run 2 | 0.14 | −0.92 | −0.78 |
| Production run 3 | −0.31 | −0.95 | −1.30 |
| Production run 4 | −0.09 | −0.35 | −0.44 |
| Average | −0.1 ± 0.1 | −0.7 ± 0.3 | −0.8 ± 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Španinger, E.; Potočnik, U.; Bren, U. Molecular Dynamics Simulations Predict that rSNP Located in the HNF-1α Gene Promotor Region Linked with MODY3 and Hepatocellular Carcinoma Promotes Stronger Binding of the HNF-4α Transcription Factor. Biomolecules 2020, 10, 1700. https://doi.org/10.3390/biom10121700
Španinger E, Potočnik U, Bren U. Molecular Dynamics Simulations Predict that rSNP Located in the HNF-1α Gene Promotor Region Linked with MODY3 and Hepatocellular Carcinoma Promotes Stronger Binding of the HNF-4α Transcription Factor. Biomolecules. 2020; 10(12):1700. https://doi.org/10.3390/biom10121700
Chicago/Turabian StyleŠpaninger, Eva, Uroš Potočnik, and Urban Bren. 2020. "Molecular Dynamics Simulations Predict that rSNP Located in the HNF-1α Gene Promotor Region Linked with MODY3 and Hepatocellular Carcinoma Promotes Stronger Binding of the HNF-4α Transcription Factor" Biomolecules 10, no. 12: 1700. https://doi.org/10.3390/biom10121700
APA StyleŠpaninger, E., Potočnik, U., & Bren, U. (2020). Molecular Dynamics Simulations Predict that rSNP Located in the HNF-1α Gene Promotor Region Linked with MODY3 and Hepatocellular Carcinoma Promotes Stronger Binding of the HNF-4α Transcription Factor. Biomolecules, 10(12), 1700. https://doi.org/10.3390/biom10121700