The Kinetics of Amyloid Fibril Formation by de Novo Protein Albebetin and Its Mutant Variants



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
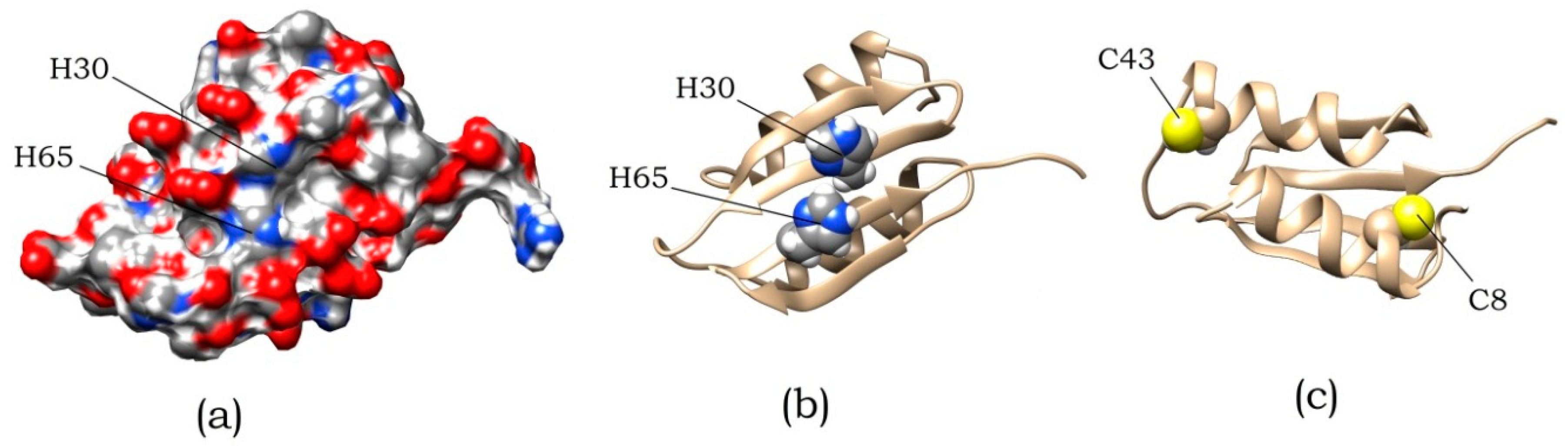{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Constructing Mutant ABB Genes
2.2. Gene Expression; Isolation and Purification of Recombinant Proteins
2.3. Protein Purity Analisis
2.4. Absorption Spectroscopy
2.5. Fluorescence Spectroscopy
2.6. Light Scattering
2.7. CD Spectroscopy
2.8. Electron Microscopy
2.9. Atomic Force Microscopy
2.10. Confocal Fluorescence Microscopy
2.11. Error Calculation
3. Results and Discussion
3.1. Studying the Artificial Protein ABB Amyloid Formation
3.1.1. Fluorescence Spectroscopy and Light Scattering
3.1.2. Far-UV Circular Dichroism
3.1.3. Electron Microscopy
3.1.4. Atomic Force Microscopy
3.1.5. Confocal Fluorescence Microscopy
3.2. The Effect of External Factors on Amyloid Formation
3.3. Design of Mutant Variants of ABB and Study of Their Amyloidogenicity
3.3.1. Studying the Kinetics of Amyloid Formation by the Mutant Variants ABB(H30F) and ABB(H65F)
3.3.2. Studying the Kinetics of Amyloid Formation by the Mutant Variant ABB(C8S/C43S)
3.3.3. Studying the Kinetics of Amyloid Formation by the ABB-TRX Hybrid Protein
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lutter, L.; Serpell, C.J.; Tuite, M.F.; Xue, W.F. The molecular lifecycle of amyloid—Mechanism of assembly, mesoscopic organisation, polymorphism, suprastructures, and biological consequences. BBA Proteins Proteom. 2019, 11, 140257. [Google Scholar] [CrossRef] [PubMed]
- Eanes, E.D.; Glenner, J. X-ray diffraction studies on amyloid filaments. Histochem. Cytochem. 1968, 16, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Nizhnikov, A.A.; Antonets, K.S.; Inge-Vechtomov, S.G. Amyloids: From Pathogenesis to Function. Biochemistry (Moscow) 2015, 80–89, 1127–1144. [Google Scholar] [CrossRef]
- Maji, S.K.; Perrin, M.H.; Sawaya, M.R.; Jessberger, S.; Vadodaria, K.; Rissman, R.A.; Singru, P.S.; Nilsson, K.P.; Simon, R.; Schubert, D.; et al. Functional amyloids as natural storage of peptide hormones in pituitary secretory granules. Science 2009, 325, 328–332. [Google Scholar] [CrossRef] [Green Version]
- Guyonnet, B.; Egge, N.; Cornwall, G.A. Functional amyloids in the mouse sperm acrosome. Mol. Cell. Biol. 2014, 34, 2624–2634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; McQuade, T.; Siemer, A.B.; Napetschnig, J.; Moriwaki, K.; Hsiao, Y.S.; Damko, E.; Moquin, D.; Walz, T.; McDermott, A.; et al. The RIP1/RIP3 necrosome forms a functional amyloid signaling complex required for programmed necrosis. Cell 2012, 150, 339–350. [Google Scholar] [CrossRef] [Green Version]
- Romero, D.; Aguilar, C.; Losick, R.; Kolter, R. Amyloid fibers provide structural integrity to Bacillus subtilis biofilms. Proc. Natl. Acad. Sci. USA 2010, 107, 2230–2234. [Google Scholar] [CrossRef] [Green Version]
- Dueholm, M.S.; Petersen, S.V.; Sonderkaer, M.; Larsen, P.; Christiansen, G.; Hein, K.L.; Enghild, J.J.; Nielsen, J.L.; Nielsen, K.L.; Nielsen, P.H.; et al. Functional amyloid in Pseudomonas. Mol. Microbiol. 2010, 77, 1009–1020. [Google Scholar] [CrossRef] [Green Version]
- Alteri, C.J.; Xicohtencatl Cortes, J.; Hess, S.; Caballero Olin, G.; Giron, J.A.; Friedman, R.L. Mycobacterium tuberculosis produces pili during human infection. Proc. Natl. Acad. Sci. USA 2007, 104, 5145–5150. [Google Scholar] [CrossRef] [Green Version]
- Fowler, D.M.; Koulov, A.V.; Alory Jost, C.; Marks, M.S.; Balch, W.E.; Kelly, J.W. Functional amyloid formation within mammalian tissue. PLoS Biol. 2006, 4, e6. [Google Scholar] [CrossRef]
- Dolgikh, D.A.; Kolomiets, A.P.; Bolotina, I.A.; Ptitsyn, O.B. Molten-globule state accumulates in carbonic-anhydrase folding. FEBS Lett. 1984, 165, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Krobath, H.; Estacio, S.G.; Faisca, P.F.N.; Shakhnovich, E.I. Identification of conserved aggregation-prone intermediate state in the folding pathways of Sps-SH3 amyloidogenic variants. J. Mol. Biol. 2012, 422, 705–722. [Google Scholar] [CrossRef] [PubMed]
- Fedorov, A.N.; Dolgikh, D.A.; Chemeris, V.V.; Chernov, B.K.; Finkelshtein, A.V.; Shulga, A.A.; Alakhov, Y.B.; Kirpichnikov, M.P.; Ptitsyn, O.B. De novo design, synthesis and study of albebetin, a polypeptide with a predetermined 3-dimensional structure—Probing the structure at the nanogram level. J. Mol. Biol. 1992, 225, 927–931. [Google Scholar] [CrossRef]
- Dolgikh, D.A.; Gabrielyan, A.E.; Navolotskaya, Y.V.; Chemeris, V.V.; Kirpichnikov, M.P. Artificial protein with a set spatial structure and biological activity. Biophysics 1993, 38, 59–65. [Google Scholar]
- Aphasizheva, I.Y.; Dolgikh, D.A.; Abdullaev, Z.K.; Uversky, V.N.; Kirpichnikov, M.P.; Ptitsyn, O.B. Can grafting of an octapeptide improve the structure of de novo protein? FEBS 1998, 425, 101–104. [Google Scholar] [CrossRef] [Green Version]
- Dolgikh, D.A.; Uversky, V.N.; Gabrielyan, A.E.; Chemeris, V.V.; Fedorov, A.N.; Navolotskaya, Y.V.; Zav’yalov, V.P.; Kirpichnikov, M.P. The de novo protein with grafted biological function: Transferring of interferon blast-transforming activity to albebetin. Protein Eng. 1996, 9, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Aphasizheva, I.Y.; Dolgikh, D.A.; Kirpichnikov, M.P. Study of immunogenic properties of albeferon, biologically active de novo protein. Mol. Biol. 1999, 33–34, 602–606. [Google Scholar]
- Chertkova, R.V.; Abdullaev, Z.K.; Dolgikh, D.A.; Zav’yalov, V.P.; Kirpichnikov, M.P. Artificial proteins with antiviral properties of alpha-interferons. Russ. J. Bioorg. Chem. 2003, 29, 231–241. [Google Scholar] [CrossRef]
- Chertkova, R.V.; Kostanyan, I.A.; Astapova, M.V.; Surina, E.A.; Dolgikh, D.A.; Kirpichnikov, M.P. An artificial protein with the biological activity of the differentiation factor for the HL-60 cell line of human promyelocyte leukemia. Russ. J. Bioorg. Chem. 2003, 29, 30–37. [Google Scholar]
- Bocharova, O.V.; Moshkovskii, S.A.; Chertkova, R.V.; Kolesanova, E.F.; Dolgikh, D.A.; Kirpichnikov, M.P. The immune response to artificial proteins containing biologically active fragments of human IFN-alpha 2 and insulin. Mol. Biol. 2002, 36, 65–70. [Google Scholar] [CrossRef]
- Bocharova, O.V.; Moshkovskii, S.A.; Chertkova, R.V.; Abdullaev, Z.K.; Kolesanova, E.F.; Dolgikh, D.A.; Kirpichnikov, M.P. Attachment of biologically active fragments of interferon-alpha 2 and insulin to the artificial protein, albebetin, affects immunogenicity of the resulting construct. Vopr. Meditsinskoi Khimii 2002, 48, 94–102. [Google Scholar]
- Morozova-Roche, L.A.; Zamotin, V.; Malisauskas, M.; Ohman, A.; Chertkova, R.; Lavrikova, M.A.; Kostanyan, I.A.; Dolgikh, D.A.; Kirpichnikov, M.P. Fibrillation of carrier protein albebetin and its biologically active constructs. multiple oligomeric intermediates and pathways. Biochemistry 2004, 43, 9609–9619. [Google Scholar] [CrossRef] [PubMed]
- Lavrikova, M.A.; Zamotin, V.V.; Malisauskas, M.; Chertkova, R.V.; Kostanyan, I.A.; Dolgikh, D.A.; Kirpichnikov, M.P.; Morozova-Roche, L.A. Amyloidogenic properties of the artificial protein albebetin and its biologically active derivatives. The role of electrostatic interactions in fibril formation. Biochemistry (Moscow) 2006, 3, 386–395. [Google Scholar] [CrossRef]
- Zamotin, V.; Gharibyana, A.; Gibanova, N.V.; Lavrikova, M.A.; Dolgikh, D.A.; Kirpichnikov, M.P.; Kostanyan, I.A.; Morozova-Roche, L.A. Cytotoxicity of albebetin oligomers depends on cross-beta-sheet formation. FEBS Lett. 2006, 580, 2451–2457. [Google Scholar] [CrossRef] [Green Version]
- Schagger, H.; Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef]
- Provencher, S.W.; Glocker, J. Estimation of globular protein secondary structure from circular dichroism. Biochemistry 1981, 20, 33–37. [Google Scholar] [CrossRef]
- Brender, J.R.; Krishnamoorthy, J.; Sciacca, M.F.; Vivekanandan, S.; D’Urso, L.; Chen, J.; La Rosa, C.; Ramamoorthy, A. Probing the sources of the apparent irreproducibility of amyloid formation: Drastic changes in kinetics and a switch in mechanism due to micellelike oligomer formation at critical concentrations of IAPP. J. Phys. Chem. B 2015, 119, 2886–2896. [Google Scholar] [CrossRef]
- Dovidchenko, N.V.; Finkelstein, A.V.; Galzitskaya, O.V. How to determine the size of folding nuclei of protofibrils from the concentration dependence of the rate and lag-time of aggregation. I. Modeling the amyloid photofibril formation. J. Phys. Chem. B 2014, 118, 1189–1197. [Google Scholar] [CrossRef]
- Galzitskaya, O.V.; Garbuzynskiy, S.O.; Lobanov, M.Y. Is it possible to predict amyloidogenic regions from sequence alone? J. Bioinform. Comput. Biol. 2006, 4, 373–388. [Google Scholar] [CrossRef]
- Raman, S.; Vernon, R.; Thompson, J.; Tyka, M.; Sadreyev, R.; Pei, J.; Kim, D.; Kellogg, E.; DiMaio, F.; Lange, O.; et al. Structure prediction for CASP8 with all-atom refinement using Rosetta. Proteins 2009, 77 (Suppl. 9), 89–99. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Gong, H.; Sun, Y.; Yan, J.; Cheng, B.; Zhang, X.; Huang, J.; Yu, M.; Guo, Y.; Zheng, L.; et al. Dissecting the role of disulfide bonds on the amyloid formation of insulin. Biochem. Biophys. Res. Commun. 2012, 423, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Vinther, T.N.; Norrman, M.; Ribe, U.; Huus, K.; Schlein, M.; Steensgaard, D.B.; Pedersen, T.A.; Pettersson, I.; Ludvigsen, S.; Kjeldsen, T.; et al. Insulin analog with additional disulfide bond has increased stability and preserved activity. Protein Sci. 2013, 22, 296–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piejko, M.; Dec, R.; Babenko, V.; Hoang, A.; Szewczyk, M.; Mak, P.; Dzwolak, W. Highly amyloidogenic two-chain peptide fragments are released upon partial digestion of insulin with pepsin. J. Biol. Chem. 2015, 290, 5947–5958. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, N.; Kumar, M.; Dubey, V.K. Effect of sodium tetrathionate on amyloid fibril: Insight into the role of disulfide bond in amyloid progression. Biochimie 2011, 93–95, 962–968. [Google Scholar] [CrossRef] [PubMed]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balobanov, V.; Chertkova, R.; Egorova, A.; Dolgikh, D.; Bychkova, V.; Kirpichnikov, M. The Kinetics of Amyloid Fibril Formation by de Novo Protein Albebetin and Its Mutant Variants. Biomolecules 2020, 10, 241. https://doi.org/10.3390/biom10020241
Balobanov V, Chertkova R, Egorova A, Dolgikh D, Bychkova V, Kirpichnikov M. The Kinetics of Amyloid Fibril Formation by de Novo Protein Albebetin and Its Mutant Variants. Biomolecules. 2020; 10(2):241. https://doi.org/10.3390/biom10020241
Chicago/Turabian StyleBalobanov, Vitalii, Rita Chertkova, Anna Egorova, Dmitry Dolgikh, Valentina Bychkova, and Mikhail Kirpichnikov. 2020. "The Kinetics of Amyloid Fibril Formation by de Novo Protein Albebetin and Its Mutant Variants" Biomolecules 10, no. 2: 241. https://doi.org/10.3390/biom10020241
APA StyleBalobanov, V., Chertkova, R., Egorova, A., Dolgikh, D., Bychkova, V., & Kirpichnikov, M. (2020). The Kinetics of Amyloid Fibril Formation by de Novo Protein Albebetin and Its Mutant Variants. Biomolecules, 10(2), 241. https://doi.org/10.3390/biom10020241