Mitochondrial HMG-Box Containing Proteins: From Biochemical Properties to the Roles in Human Diseases

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Mitochondrial DNA Forms Higher-Order Structures Called Mitochondrial Nucleoids
3. Setting the Stage: The Identification of HMG-Box Proteins in Yeast and Human Mitochondria
4. Genetic Studies on mtHMG Proteins Reveal Their Role in Regulating mtDNA Copy Number and Mt-Nucleoid Morphology and Dynamics
5. mtHMG Proteins Are Rapidly Evolving
6. mtHMG Proteins Protect mtDNA against Damage and the Accumulation of Mutations
7. On the Mechanism of mtDNA Compaction by mtHMG Proteins
8. Regulation of mtHMG Proteins
9. Involvement of TFAM in Cellular Pathologies
10. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sagan, L. On the origin of mitosing cells. J. Theor. Biol. 1967, 14, 255–274. [Google Scholar] [CrossRef]
- Margulis, L. Origin of Eukaryotic Cells; Evidence and Research Implications for a Theory of the Origin and Evolution of Microbial, Plant, and Animal Cells on the Precambrian Earth, 1st ed.; Yale University Press: New Haven, NH, USA, 1970. [Google Scholar]
- Archibald, J.M. One Plus one Equals One: Symbiosis and the Evolution of Complex Life, 1st ed.; Oxford University Press: New York, NY, USA, 2016. [Google Scholar]
- Archibald, J.M. Endosymbiosis and eukaryotic cell evolution. Curr. Biol. 2015, 25, R911–R921. [Google Scholar] [CrossRef] [Green Version]
- Martijn, J.; Vosseberg, J.; Guy, L.; Offre, P.; Ettema, T.J.G. Deep mitochondrial origin outside the sampled alphaproteobacteria. Nature 2018, 557, 101–105. [Google Scholar] [CrossRef]
- Martin, W.F.; Garg, S.; Zimorski, V. Endosymbiotic theories for eukaryote origin. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140330. [Google Scholar] [CrossRef] [Green Version]
- Zaremba-Niedzwiedzka, K.; Caceres, E.F.; Saw, J.H.; Bäckström, D.; Juzokaite, L.; Vancaester, E.; Seitz, K.W.; Anantharaman, K.; Starnawski, P.; Kjeldsen, K.U.; et al. Asgard archaea illuminate the origin of eukaryotic cellular complexity. Nature 2017, 541, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Eme, L.; Spang, A.; Lombard, J.; Stairs, C.W.; Ettema, T.J.G. Archaea and the origin of eukaryotes. Nat. Rev. Microbiol. 2017, 15, 711–723. [Google Scholar] [CrossRef]
- Johnston, I.G.; Williams, B.P. Evolutionary inference across eukaryotes identifies specific pressures favoring mitochondrial gene retention. Cell Syst. 2016, 2, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.F. Why chloroplasts and mitochondria retain their own genomes and genetic systems: Colocation for redox regulation of gene expression. Proc. Natl. Acad. Sci. USA 2015, 112, 10231–10238. [Google Scholar] [CrossRef] [Green Version]
- Labbé, K.; Murley, A.; Nunnari, J. Determinants and functions of mitochondrial behavior. Annu. Rev. Cell Dev. Biol. 2014, 30, 357–391. [Google Scholar] [CrossRef]
- Klecker, T.; Westermann, B. Asymmetric inheritance of mitochondria in yeast. Biol. Chem. 2020, 401, 779–791. [Google Scholar] [CrossRef]
- Friedman, J.R.; Nunnari, J. Mitochondrial form and function. Nature 2014, 505, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Bendich, A.J. Structural analysis of mitochondrial DNA molecules from fungi and plants using moving pictures and pulsed-field gel electrophoresis. J. Mol. Biol. 1996, 255, 564–588. [Google Scholar] [CrossRef]
- Grimes, G.W.; Mahler, H.R.; Perlman, P.S. Nuclear gene dosage effects on mitochondrial mass and DNA. J. Cell Biol. 1974, 61, 565–574. [Google Scholar] [CrossRef]
- Foury, F.; Roganti, T.; Lecrenier, N.; Purnelle, B. The complete sequence of the mitochondrial genome of Saccharomyces cerevisiae. FEBS Lett. 1998, 440, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Williamson, D. The curious history of yeast mitochondrial DNA. Nat. Rev. Genet. 2002, 3, 475–481. [Google Scholar] [CrossRef]
- Wai, T.; Ao, A.; Zhang, X.; Cyr, D.; Dufort, D.; Shoubridge, E.A. The role of mitochondrial DNA copy number in mammalian fertility. Biol. Reprod. 2010, 83, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Anderson, S.; Bankier, A.T.; Barrell, B.G.; De Bruijn, M.H.L.; Coulson, A.R.; Drouin, J.; Eperon, I.C.; Nierlich, D.P.; Roe, B.A.; Sanger, F.; et al. Sequence and organization of the human mitochondrial genome. Nature 1981, 290, 457–465. [Google Scholar] [CrossRef]
- Marquard, K.L.; Stephens, S.M.; Jungheim, E.S.; Ratts, V.S.; Odem, R.R.; Lanzendorf, S.; Moley, K.H. Polycystic ovary syndrome and maternal obesity affect oocyte size in in vitro fertilization/intracytoplasmic sperm injection cycles. Fertil. Steril. 2011, 95, 2146–2149. [Google Scholar] [CrossRef] [Green Version]
- O’Hara, R.; Tedone, E.; Ludlow, A.; Huang, E.; Arosio, B.; Mari, D.; Shay, J.W. Quantitative mitochondrial DNA copy number determination using droplet digital PCR with single-cell resolution. Genome Res. 2019, 29, 1878–1888. [Google Scholar] [CrossRef] [Green Version]
- Liou, C.-W.; Lin, T.-K.; Chen, J.-B.; Tiao, M.-M.; Weng, S.-W.; Chen, S.-D.; Chuang, Y.-C.; Chuang, J.-H.; Wang, P.-W. Association between a common mitochondrial DNA D-loop polycytosine variant and alteration of mitochondrial copy number in human peripheral blood cells. J. Med. Genet. 2010, 47, 723–728. [Google Scholar] [CrossRef]
- Miyakawa, I. Organization and dynamics of yeast mitochondrial nucleoids. Proc. Japan Acad. Ser. B 2017, 93, 339–359. [Google Scholar] [CrossRef] [Green Version]
- Miyakawa, I.; Sando, N.; Kawano, S.; Nakamura, S.; Kuroiwa, T. Isolation of morphologically intact mitochondrial nucleoids from the yeast, Saccharomyces cerevisiae. J. Cell Sci. 1987, 88, 431–439. [Google Scholar]
- Miyakawa, I.; Kanayama, M.; Fujita, Y.; Sato, H. Morphology and protein composition of the mitochondrial nucleoids in yeast cells lacking Abf2p, a high mobility group protein. J. Gen. Appl. Microbiol. 2010, 56, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Williamson, D.H.; Fennell, D.J. Visualization of yeast mitochondrial DNA with the fluorescent stain “DAPI”. Methods Enzymol. 1979, 56, 728–733. [Google Scholar] [CrossRef]
- Williamson, D.H.; Fennell, D.J. The use of fluorescent DNA-binding agent for detecting and separating yeast mitochondrial DNA. Methods Cell Biol. 1975, 12, 335–351. [Google Scholar] [CrossRef]
- Kaufman, B.A.; Newman, S.M.; Hallberg, R.L.; Slaughter, C.A.; Perlman, P.S.; Butow, R.A. In organello formaldehyde crosslinking of proteins to mtDNA: Identification of bifunctional proteins. Proc. Natl. Acad. Sci. USA 2000, 97, 7772–7777. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, B.A.; Kolesar, J.E.; Perlman, P.S.; Butow, R.A. A function for the mitochondrial chaperonin Hsp60 in the structure and transmission of mitochondrial DNA nucleoids in Saccharomyces cerevisiae. J. Cell Biol. 2003, 163, 457–461. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, S.; Inai, T.; Miyakawa, I. Morphology of mitochondrial nucleoids in respiratory-deficient yeast cells varies depending on the unit length of the mitochondrial DNA sequence. FEMS Yeast Res. 2016, 16, fow055. [Google Scholar] [CrossRef] [Green Version]
- Miyakawa, I.; Aoi, H.; Sando, N.; Kuroiwa, T. Fluorescence microscopic studies of mitochondrial nucleoids during meiosis and sporulation in the yeast, Saccharomyces cerevisiae. J. Cell Sci. 1984, 66, 21–38. [Google Scholar]
- Farge, G.É.R.; Falkenberg, M. Organization of DNA in mammalian mitochondria. Int. J. Mol. Sci. 2019, 20, 2770. [Google Scholar] [CrossRef] [Green Version]
- Bogenhagen, D.F. Mitochondrial DNA nucleoid structure. Biochim. Biophys. Acta—Gene Regul. Mech. 2012, 1819, 914–920. [Google Scholar] [CrossRef]
- Bonekamp, N.A.; Larsson, N.G. SnapShot: Mitochondrial nucleoid. Cell 2018, 172, 388. [Google Scholar] [CrossRef] [PubMed]
- Garrido, N.; Griparic, L.; Jokitalo, E.; Wartiovaara, J.; Van der Bliek, A.M.; Spelbrink, J.N. Composition and dynamics of human mitochondrial nucleoids. Mol. Biol. Cell 2003, 14, 1583–1596. [Google Scholar] [CrossRef] [Green Version]
- Gilkerson, R.; Bravo, L.; Garcia, I.; Gaytan, N.; Herrera, A.; Maldonado, A.; Quintanilla, B. The mitochondrial nucleoid: Integrating mitochondrial DNA into cellular homeostasis. Cold Spring Harb. Perspect. Biol. 2013, 5, a011080. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, B.A.; Durisic, N.; Mativetsky, J.M.; Costantino, S.; Hancock, M.A.; Grutter, P.; Shoubridge, E.A. The mitochondrial transcription factor TFAM coordinates the assembly of multiple DNA molecules into nucleoid-like structures. Mol. Biol. Cell 2007, 18, 3225–3236. [Google Scholar] [CrossRef]
- Prachař, J. Ultrastructure of mitochondrial nucleoid and its surroundings. Gen. Physiol. Biophys. 2016, 35, 273–286. [Google Scholar] [CrossRef] [Green Version]
- Kukat, C.; Wurm, C.A.; Spåhr, H.; Falkenberg, M.; Larsson, N.G.; Jakobs, S. Super-resolution microscopy reveals that mammalian mitochondrial nucleoids have a uniform size and frequently contain a single copy of mtDNA. Proc. Natl. Acad. Sci. USA 2011, 108, 13534–13539. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.A.; Tkachuk, A.N.; Shtengel, G.; Kopek, B.G.; Bogenhagen, D.F.; Hess, H.F.; Clayton, D.A. Superresolution fluorescence imaging of mitochondrial nucleoids reveals their spatial range, limits, and membrane interaction. Mol. Cell. Biol. 2011, 31, 4994–5010. [Google Scholar] [CrossRef] [Green Version]
- Bogenhagen, D.F.; Wang, Y.; Shen, E.L.; Kobayashi, R. Protein components of mitochondrial DNA nucleoids in higher eukaryotes. Mol. Cell. Proteomics. 2003, 2, 1205–1216. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Bogenhagen, D.F. Human mitochondrial DNA nucleoids are linked to protein folding machinery and metabolic enzymes at the mitochondrial inner membrane. J. Biol. Chem. 2006, 281, 25791–25802. [Google Scholar] [CrossRef] [Green Version]
- Bogenhagen, D.F.; Rousseau, D.; Burke, S. The layered structure of human mitochondrial DNA nucleoids. J. Biol. Chem. 2008, 283, 3665–3675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.J.; Wang, X.; Kaufman, B.A.; Butow, R.A. Aconitase couples metabolic regulation to mitochondrial DNA maintenance. Science 2005, 307, 714–717. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Butow, R.A. The organization and inheritance of the mitochondrial genome. Nat. Rev. Genet. 2005, 6, 815–825. [Google Scholar] [CrossRef]
- Miyakawa, I.; Okamuro, A.; Kinsky, S.; Visacka, K.; Tomaska, L.; Nosek, J. Mitochondrial nucleoids from the yeast Candida parapsilosis: Expansion of the repertoire of proteins associated with mitochondrial DNA. Microbiology 2009, 155, 1558–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nosek, J.; Tomaska, L.; Bolotin-Fukuhara, M.; Miyakawa, I. Mitochondrial chromosome structure: An insight from analysis of complete yeast genomes. FEMS Yeast Res. 2006, 6, 356–370. [Google Scholar] [CrossRef] [PubMed]
- Contamine, V.; Picard, M. Maintenance and integrity of the mitochondrial genome: A plethora of nuclear genes in the budding yeast. Microbiol. Mol. Biol. Rev. 2000, 64, 281–315. [Google Scholar] [CrossRef] [Green Version]
- Rajala, N.; Hensen, F.; Wessels, H.J.C.T.; Ives, D.; Gloerich, J.; Spelbrink, J.N. Whole cell formaldehyde cross-linking simplifies purification of mitochondrial nucleoids and associated proteins involved in mitochondrial gene expression. PLoS ONE 2015, 10, e0116726. [Google Scholar] [CrossRef] [Green Version]
- Kucej, M.; Kucejova, B.; Subramanian, R.; Chen, X.J.; Butow, R.A. Mitochondrial nucleoids undergo remodeling in response to metabolic cues. J. Cell Sci. 2008, 121, 1861–1868. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.R.; Han, J. Mitochondrial nucleoid: Shield and switch of the mitochondrial genome. Oxid. Med. Cell. Longev. 2017, 2017, 8060949. [Google Scholar] [CrossRef]
- Meeusen, S.; Nunnari, J. Evidence for a two membrane-spanning autonomous mitochondrial DNA replisome. J. Cell Biol. 2003, 163, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Gustafsson, C.M.; Falkenberg, M.; Larsson, N.-G. Maintenance and expression of mammalian mitochondrial DNA. Annu. Rev. Biochem. 2016, 85, 133–160. [Google Scholar] [CrossRef] [PubMed]
- Caron, F.; Jacq, C.; Rouviere Yaniv, J. Characterization of a histone-like protein extracted from yeast mitochondria. Proc. Natl. Acad. Sci. USA 1979, 76, 4265–4269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouvière-Yaniv, J.; Gros, F. Characterization of a novel, low molecular weight DNA binding protein from Escherichia coli. Proc. Natl. Acad. Sci. USA 1975, 72, 3428–3432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Certa, U.; Colavito-Shepanski, M.; Grunstein, M. Yeast may not contain histone H1: The only known “histone H1-like” protein in Saccharomyces cerevisiae is a mitochondrial protein. Nucleic Acids Res. 1984, 12, 7975–7985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommer, A. Yeast chromatin: Search for histone H1. Mol. Gen. Genet. 1978, 161, 323–331. [Google Scholar] [CrossRef]
- Diffley, J.F.X.; Stillman, B. A close relative of the nuclear, chromosomal high-mobility group protein HMG1 in yeast mitochondria. Proc. Natl. Acad. Sci. USA 1991, 88, 7864–7868. [Google Scholar] [CrossRef] [Green Version]
- Diffley, J.F.X.; Stillman, B. Purification of a yeast protein that binds to origins of DNA replication and a transcriptional silencer. Proc. Natl. Acad. Sci. USA 1988, 85, 2120–2124. [Google Scholar] [CrossRef] [Green Version]
- Diffley, J.F.X.; Stillman, B. Similarity between the transcriptional silencer binding proteins Abf1 and Rap1. Science 1989, 246, 1034–1038. [Google Scholar] [CrossRef]
- Vizoso-Vázquez, Á.; Lamas-Maceiras, M.; Becerra, M.; González-Siso, M.I.; Rodríguez-Belmonte, E.; Cerdán, M.E. Ixr1p and the control of the Saccharomyces cerevisiae hypoxic response. Appl. Microbiol. Biotechnol. 2012, 94, 173–184. [Google Scholar] [CrossRef]
- Brown, S.J.; Kellett, P.J.; Lippard, S.J. Ixr1, a yeast protein that binds to platinated DNA and confers sensitivity to cisplatin. Science 1993, 261, 603–605. [Google Scholar] [CrossRef]
- Byrne, K.P.; Wolfe, K.H. The Yeast Gene Order Browser: Combining curated homology and syntenic context reveals gene fate in polyploid species. Genome Res. 2005, 15, 1456–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierro, P.; Capaccio, L.; Gadaleta, G. The 25 kDa protein recognizing the rat curved region upstream of the origin of the L-strand replication is the rat homologue of the human mitochondrial transcription factor A. FEBS Lett. 1999, 457, 307–310. [Google Scholar] [CrossRef] [Green Version]
- Alami-Ouahabi, N.; Veilleux, S.; Meistrich, M.L.; Boissonneault, G. The testis-specific high-mobility-group protein, a phosphorylation-dependent DNA-packaging factor of elongating and condensing spermatids. Mol. Cell. Biol. 1996, 16, 3720–3729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, L.R.; Megraw, T.L.; Chae, C.B. Essential role of the HMG domain in the function of yeast mitochondrial histone HM: Functional complementation of HM by the nuclear nonhistone protein NHP6A. Proc. Natl. Acad. Sci. USA 1993, 90, 5598–5602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxevanis, A.D.; Landsman, D. The HMG-1 box protein family: Classification and functional relationships. Nucleic Acids Res. 1995, 23, 1604–1613. [Google Scholar] [CrossRef] [PubMed]
- Bustin, M. Revised nomenclature for high mobility group (HMG) chromosomal proteins. Trends Biochem. Sci. 2001, 26, 152–153. [Google Scholar] [CrossRef]
- Megraw, T.L.; Chae, C.-B. Functional complementarity between the HMG1-like yeast mitochondrial histone HM and the bacterial histone-like protein HU. J. Biol. Chem. 1993, 268, 12758–12763. [Google Scholar]
- Dillon, S.C.; Dorman, C.J. Bacterial nucleoid-associated proteins, nucleoid structure and gene expression. Nat. Rev. Microbiol. 2010, 8, 185–195. [Google Scholar] [CrossRef]
- Kobayashi, T.; Takahara, M.; Miyagishima, S.Y.; Kuroiwa, H.; Sasaki, N.; Ohta, N.; Matsuzaki, M.; Kuroiwa, T. Detection and localization of a chloroplast-encoded HU-like protein that organizes chloroplast nucleoids. Plant. Cell 2002, 14, 1579–1589. [Google Scholar] [CrossRef] [Green Version]
- Pastor, M.M.; Proft, M.; Pascual-Ahuir, A. Mitochondrial function is an inducible determinant of osmotic stress adaptation in yeast. J. Biol. Chem. 2009, 284, 30307–30317. [Google Scholar] [CrossRef] [Green Version]
- Diffley, J.F.; Stillman, B. DNA binding properties of an HMG1-related protein from yeast mitochondria. J. Biol. Chem. 1992, 267, 3368–3374. [Google Scholar]
- Fisher, R.P.; Clayton, D.A. Purification and characterization of human mitochondrial transcription factor 1. Mol. Cell. Biol. 1988, 8, 3496–3509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, R.P.; Parisi, M.A.; Clayton, D.A. Flexible recognition of rapidly evolving promoter sequences by mitochondrial transcription factor 1. Genes Dev. 1989, 3, 2202–2217. [Google Scholar] [CrossRef] [PubMed]
- Parisi, M.A.; Clayton, D.A. Similarity of human mitochondrial transcription factor 1 to high mobility group proteins. Science 1991, 252, 965–969. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.P.; Lisowsky, T.; Breen, G.A.M.; Clayton, D.A. A rapid, efficient method for purifying DNA-binding proteins. J. Biol. Chem. 1991, 266, 9153–9160. [Google Scholar]
- Barshad, G.; Marom, S.; Cohen, T.; Mishmar, D. Mitochondrial DNA transcription and its regulation: An evolutionary perspective. Trends Genet. 2018, 34, 682–692. [Google Scholar] [CrossRef]
- Bouda, E.; Stapon, A.; Garcia-Diaz, M. Mechanisms of mammalian mitochondrial transcription. Protein Sci. 2019, 28, 1594–1605. [Google Scholar] [CrossRef]
- Fisher, R.P.; Lisowsky, T.; Parisi, M.A.; Clayton, D.A. DNA wrapping and bending by a mitochondrial high mobility group-like transcriptional activator protein. J. Biol. Chem. 1992, 267, 3358–3367. [Google Scholar]
- Takamatsu, C.; Umeda, S.; Ohsato, T.; Ohno, T.; Abe, Y.; Fukuoh, A.; Shinagawa, H.; Hamasaki, N.; Kang, D. Regulation of mitochondrial D-loops by transcription factor A and single-stranded DNA-binding protein. EMBO Rep. 2002, 3, 451–456. [Google Scholar] [CrossRef] [Green Version]
- Kanki, T.; Ohgaki, K.; Gaspari, M.; Gustafsson, C.M.; Fukuoh, A.; Sasaki, N.; Hamasaki, N.; Kang, D. Architectural role of mitochondrial transcription factor A in maintenance of human mitochondrial DNA. Mol. Cell. Biol. 2004, 24, 9823–9834. [Google Scholar] [CrossRef] [Green Version]
- Campbell, C.T.; Kolesar, J.E.; Kaufman, B.A. Mitochondrial transcription factor A regulates mitochondrial transcription initiation, DNA packaging, and genome copy number. Biochim. Biophys. Acta—Gene Regul. Mech. 2012, 1819, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Clayton, D.A. Assignment of a yeast protein necessary for mitochondrial transcription initiation. Nucleic Acids Res. 1992, 20, 1053–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dyck, E.; Clayton, D.A. Transcription-dependent DNA transactions in the mitochondrial genome of a yeast hypersuppressive petite mutant. Mol. Cell. Biol. 1998, 18, 2976–2985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dairaghi, D.J.; Shadel, G.S.; Clayton, D.A. Addition of a 29 residue carboxyl-terminal tail converts a simple HMG box-containing protein into a transcriptional activator. J. Mol. Biol. 1995, 249, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Goto, A.; Matsushima, Y.; Kadowaki, T.; Kitagawa, Y. Drosophila mitochondrial transcription factor A (d-TFAM) is dispensable for the transcription of mitochondrial DNA in Kc167 cells. Biochem. J. 2001, 354, 243–248. [Google Scholar] [CrossRef]
- Parisi, M.A.; Xu, B.; Clayton, D.A. A human mitochondrial transcriptional activator can functionally replace a yeast mitochondrial HMG-box protein both in vivo and in vitro. Mol. Cell. Biol. 1993, 13, 1951–1961. [Google Scholar] [CrossRef] [Green Version]
- Yoon, Y.G.; Koob, M.D.; Yoo, Y.H. Mitochondrial genome-maintaining activity of mouse mitochondrial transcription factor A and its transcript isoform in Saccharomyces cerevisiae. Gene 2011, 484, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Yoon, Y.G. Transfer of xenomitochondria containing the entire mouse mitochondrial genome into a genetically modified yeast expressing mitochondrial transcription factor A. J. Microbiol. Biotechnol. 2020, in press. [Google Scholar] [CrossRef]
- Zelenaya-Troitskaya, O.; Newman, S.M.; Okamoto, K.; Perlman, P.S.; Butow, R.A. Functions of the high mobility group protein, Abf2p, in mitochondrial DNA segregation, recombination and copy number in Saccharomyces cerevisiae. Genetics 1998, 148, 1763–1776. [Google Scholar]
- Blank, H.M.; Li, C.; Mueller, J.E.; Bogomolnaya, L.M.; Bryk, M.; Polymenis, M. An increase in mitochondrial DNA promotes nuclear DNA replication in yeast. PLoS Genet. 2008, 4, e1000047. [Google Scholar] [CrossRef]
- Taylor, S.D.; Zhang, H.; Eaton, J.S.; Rodeheffer, M.S.; Lebedeva, M.A.; O’Rourke, T.W.; Siede, W.; Shadel, G.S. The conserved Mec1/Rad53 nuclear checkpoint pathway regulates mitochondrial DNA copy number in Saccharomyces cerevisiae. Mol. Biol. Cell 2005, 16, 3010–3018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebedeva, M.A.; Shadel, G.S. Cell cycle- and ribonucleotide reductase-driven changes in mtDNA copy number influence mtDNA inheritance without compromising mitochondrial gene expression. Cell Cycle 2007, 6, 2048–2057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsaponina, O.; Barsoum, E.; Åström, S.U.; Chabes, A. Ixr1 is required for the expression of the ribonucleotide reductase Rnr1 and maintenance of dNTP pools. PLoS Genet. 2011, 7, e1002061. [Google Scholar] [CrossRef] [Green Version]
- Bakkaiová, J.; Marini, V.; Willcox, S.; Nosek, J.; Griffith, J.D.; Krejci, L.; Tomáška, Ľ. Yeast mitochondrial HMG proteins: DNA-binding properties of the most evolutionarily divergent component of mitochondrial nucleoids. Biosci. Rep. 2016, 36, e00288. [Google Scholar] [CrossRef] [Green Version]
- Samoilova, E.O.; Krasheninnikov, I.A.; Levitskii, S.A. Interaction between Saccharomyces cerevisiae mitochondrial DNA-binding protein Abf2p and Cce1p resolvase. Biochem. 2016, 81, 1111–1117. [Google Scholar] [CrossRef] [PubMed]
- Ling, F.; Bradshaw, E.; Yoshida, M. Prevention of mitochondrial genomic instability in yeast by the mitochondrial recombinase Mhr1. Sci. Rep. 2019, 9, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Lockshon, D.; Zweifel, S.G.; Freeman-Cook, L.L.; Lorimer, H.E.; Brewer, B.J.; Fangman, W.L. A role for recombination junctions in the segregation of mitochondrial DNA in yeast. Cell 1995, 81, 947–955. [Google Scholar] [CrossRef] [Green Version]
- Gerhold, J.M.; Aun, A.; Sedman, T.; Jõers, P.; Sedman, J. Strand invasion structures in the inverted repeat of Candida albicans mitochondrial DNA reveal a role for homologous recombination in replication. Mol. Cell 2010, 39, 851–861. [Google Scholar] [CrossRef]
- Gerhold, J.M.; Sedman, T.; Visacka, K.; Slezakova, J.; Tomaska, L.; Nosek, J.; Sedman, J. Replication intermediates of the linear mitochondrial DNA of Candida parapsilosis suggest a common recombination based mechanism for yeast mitochondria. J. Biol. Chem. 2014, 289, 22659–22670. [Google Scholar] [CrossRef] [Green Version]
- Bendich, A.J. The end of the circle for yeast mitochondrial DNA. Mol. Cell 2010, 39, 831–832. [Google Scholar] [CrossRef]
- Bendich, A.J. Reaching for the ring: The study of mitochondrial genome structure. Curr. Genet. 1993, 24, 279–290. [Google Scholar] [CrossRef] [PubMed]
- MacAlpine, D.M.; Perlman, P.S.; Butow, R.A. The high mobility group protein Abf2p influences the level of yeast mitochondrial DNA recombination intermediates in vivo. Proc. Natl. Acad. Sci. USA 1998, 95, 6739–6743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, K.; Perlman, P.S.; Butow, R.A. The sorting of mitochondrial DNA and mitochondrial proteins in zygotes: Preferential transmission of mitochondrial DNA to the medial bud. J. Cell Biol. 1998, 142, 613–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, S.M.; Zelenaya-Troitskaya, O.; Perlman, P.S.; Butow, R.A. Analysis of mitochondrial DNA nucleoids in wild-type and a mutant strain of Saccharomyces cerevisiae that lacks the mitochondrial HMG box protein Abf2p. Nucleic Acids Res. 1996, 24, 386–393. [Google Scholar] [CrossRef]
- Zelenaya-Troitskaya, O.; Perlman, P.S.; Butow, R.A. An enzyme in yeast mitochondria that catalyzes a step in branched-chain amino acid biosynthesis also functions in mitochondrial DNA stability. EMBO J. 1995, 14, 3268–3276. [Google Scholar] [CrossRef]
- MacAlpine, D.M.; Perlman, P.S.; Butow, R.A. The numbers of individual mitochondrial DNA molecules and mitochondrial DNA nucleoids in yeast are co-regulated by the general amino acid control pathway. EMBO J. 2000, 19, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Bateman, J.M.; Perlman, P.S.; Butow, R.A. Mutational bisection of the mitochondrial DNA stability and amino acid biosynthetic functions of Ilv5p of budding yeast. Genetics 2002, 161, 1043–1052. [Google Scholar]
- Macierzanka, M.; Plotka, M.; Pryputniewicz-Drobinska, D.; Lewandowska, A.; Lightowlers, R.; Marszalek, J. Maintenance and stabilization of mtDNA can be facilitated by the DNA-binding activity of Ilv5p. Biochim. Biophys. Acta—Mol. Cell Res. 2008, 1783, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Huangyang, P.; Simon, M.C. Hidden features: Exploring the non-canonical functions of metabolic enzymes. DMM Dis. Model. Mech. 2018, 11, dmm033365. [Google Scholar] [CrossRef] [Green Version]
- Gancedo, C.; Flores, C.-L. Moonlighting proteins in yeasts. Microbiol. Mol. Biol. Rev. 2008, 72, 197–210. [Google Scholar] [CrossRef] [Green Version]
- Megraw, T.L.; Kao, L.R.; Chae, C.B. The mitochondrial histone HM: An evolutionary link between bacterial HU and nuclear HMG1 proteins. Biochimie 1994, 76, 909–916. [Google Scholar] [CrossRef]
- Kao, L.R.; Megraw, T.L.; Chae, C.B. SHM1: A multicopy suppressor of a temperature-sensitive null mutation in the HMG1-like abf2 gene. Yeast 1996, 12, 1239–1250. [Google Scholar] [CrossRef]
- Vozza, A.; Blanco, E.; Palmieri, L.; Palmieri, F. Identification of the mitochondrial GTP/GDP transporter in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 20850–20857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castegna, A.; Scarcia, P.; Agrimi, G.; Palmieri, L.; Rottensteiner, H.; Spera, I.; Germinario, L.; Palmieri, F. Identification and functional characterization of a novel mitochondrial carrier for citrate and oxoglutarate in Saccharomyces cerevisiae. J. Biol. Chem. 2010, 285, 17359–17370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.H.; Ha, S.J.; Kao, L.R.; Megraw, T.L.; Chae, C.-B. A novel DNA-binding protein bound to the mitochondrial inner membrane restores the null mutation of mitochondrial histone Abf2p in Saccharomyces cerevisiae. Mol. Cell. Biol. 1998, 18, 5712–5723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacovino, M.; Granycome, C.; Sembongi, H.; Bokori-Brown, M.; Butow, R.A.; Holt, I.J.; Bateman, J.M. The conserved translocase Tim17 prevents mitochondrial DNA loss. Hum. Mol. Genet. 2009, 18, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Matta, S.K.; Pareek, G.; Bankapalli, K.; Oblesha, A.; D’Silva, P. Role of Tim17 transmembrane regions in regulating the architecture of presequence translocase and mitochondrial DNA stability. Mol. Cell. Biol. 2017, 37, e00491-16. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Ivessa, A.S. Association of the yeast DNA helicase Pif1p with mitochondrial membranes and mitochondrial DNA. Eur. J. Cell Biol. 2010, 89, 742–747. [Google Scholar] [CrossRef]
- Wai, T.; Teoli, D.; Shoubridge, E.A. The mitochondrial DNA genetic bottleneck results from replication of a subpopulation of genomes. Nat. Genet. 2008, 40, 1484–1488. [Google Scholar] [CrossRef]
- DeRisi, J.L.; Iyer, V.R.; Brown, P.O. Exploring the metabolic and genetic control of gene expression on a genomic scale. Science 1997, 278, 680–686. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.J.; Wang, X.; Butow, R.A. Yeast aconitase binds and provides metabolically coupled protection to mitochondrial DNA. Proc. Natl. Acad. Sci. USA 2007, 104, 13738–13743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klausner, R.D.; Rouault, T.A. A double life: Cytosolic aconitase as a regulatory RNA binding protein. Mol. Biol. Cell 1993, 4, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, M.C.; Mende-Mueller, L.; Blondin, G.A.; Beinert, H. Purification and characterization of cytosolic aconitase from beef liver and its relationship to the iron-responsive element binding protein. Proc. Natl. Acad. Sci. USA 1992, 89, 11730–11734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangloff, S.P.; Marguet, D.; Lauquin, G.J. Molecular cloning of the yeast mitochondrial aconitase gene (ACO1) and evidence of a synergistic regulation of expression by glucose plus glutamate. Mol. Cell. Biol. 1990, 10, 3551–3561. [Google Scholar] [CrossRef]
- Liu, Z.; Butow, R.A. A transcriptional switch in the expression of yeast tricarboxylic acid cycle genes in response to a reduction or loss of respiratory function. Mol. Cell. Biol. 1999, 19, 6720–6728. [Google Scholar] [CrossRef] [Green Version]
- Shadel, G.S. Mitochondrial DNA, aconitase “wraps” it up. Trends Biochem. Sci. 2005, 30, 294–296. [Google Scholar] [CrossRef]
- Poveda-Huertes, D.; Matic, S.; Marada, A.; Habernig, L.; Licheva, M.; Myketin, L.; Gilsbach, R.; Tosal-Castano, S.; Papinski, D.; Mulica, P.; et al. An early mtUPR: Redistribution of the nuclear transcription factor Rox1 to mitochondria protects against intramitochondrial proteotoxic aggregates. Mol. Cell 2020, 77, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Larsson, N.G.; Wang, J.; Wilhelmsson, H.; Oldfors, A.; Rustin, P.; Lewandoski, M.; Barsh, G.S.; Clayton, D.A. Mitochondrial transcription factor A is necessary for mtDNA maintenance and embryogenesis in mice. Nat. Genet. 1998, 18, 231–236. [Google Scholar] [CrossRef]
- West, A.P.; Khoury-Hanold, W.; Staron, M.; Tal, M.C.; Pineda, C.M.; Lang, S.M.; Bestwick, M.; Duguay, B.A.; Raimundo, N.; MacDuff, D.A.; et al. Mitochondrial DNA stress primes the antiviral innate immune response. Nature 2015, 520, 553–557. [Google Scholar] [CrossRef] [Green Version]
- Ekstrand, M.I.; Falkenberg, M.; Rantanen, A.; Park, C.B.; Gaspari, M.; Hultenby, K.; Rustin, P.; Gustafsson, C.M.; Ran Larsson, N.-G. Mitochondrial transcription factor A regulates mtDNA copy number in mammals. Hum. Mol. Genet. 2004, 13, 935–944. [Google Scholar] [CrossRef]
- Pohjoismäki, J.L.O.; Wanrooij, S.; Hyvärinen, A.K.; Goffart, S.; Holt, I.J.; Spelbrink, J.N.; Jacobs, H.T. Alterations to the expression level of mitochondrial transcription factor A, TFAM, modify the mode of mitochondrial DNA replication in cultured human cells. Nucleic Acids Res. 2006, 34, 5815–5828. [Google Scholar] [CrossRef] [PubMed]
- Kasashima, K.; Sumitani, M.; Endo, H. Human mitochondrial transcription factor A is required for the segregation of mitochondrial DNA in cultured cells. Exp. Cell Res. 2011, 317, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, J.; Wilhelmsson, H.; Hansson, A.; Thorén, P.; Duffy, J.; Rustin, P.; Larsson, N.G. Genetic modification of survival in tissue-specific knockout mice with mitochondrial cardiomyopathy. Proc. Natl. Acad. Sci. USA 2000, 97, 3467–3472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, J.P.; Köhler, M.; Graff, C.; Oldfors, A.; Magnuson, M.A.; Berggren, P.O.; Larsson, N.G. Impaired insulin secretion and β-cell loss in tissue-specific knockout mice with mitochondrial diabetes. Nat. Genet. 2000, 26, 336–340. [Google Scholar] [CrossRef]
- Sörensen, L.; Ekstrand, M.; Silva, J.P.; Lindqvist, E.; Xu, B.; Rustin, P.; Olson, L.; Larsson, N.G. Late-onset corticohippocampal neurodepletion attributable to catastrophic failure of oxidative phosphorylation in MILON mice. J. Neurosci. 2001, 21, 8082–8090. [Google Scholar] [CrossRef]
- Wang, J.; Wilhelmsson, H.; Graff, C.; Li, H.; Oldfors, A.; Rustin, P.; Brüning, J.C.; Kahn, C.R.; Clayton, D.A.; Barsh, G.S.; et al. Dilated cardiomyopathy and atrioventricular conduction blocks induced by heart-specific inactivation of mitochondrial DNA gene expression. Nat. Genet. 1999, 21, 133–137. [Google Scholar] [CrossRef]
- Wredenberg, A.; Wibom, R.; Wilhelmsson, H.; Graff, C.; Wiener, H.H.; Burden, S.J.; Oldfors, A.; Westerblad, H.; Larsson, N.G. Increased mitochondrial mass in mitochondrial myopathy mice. Proc. Natl. Acad. Sci. USA 2002, 99, 15066–15071. [Google Scholar] [CrossRef] [Green Version]
- Buschbeck, M.; Hake, S.B. Variants of core histones and their roles in cell fate decisions, development and cancer. Nat. Rev. Mol. Cell Biol. 2017, 18, 299–314. [Google Scholar] [CrossRef]
- Dequard-Chablat, M.; Alland, C. Two copies of mtHMG1, encoding a novel mitochondrial HMG-like protein, delay accumulation of mitochondrial DNA deletions in Podospora anserina. Eukaryot. Cell 2002, 1, 503–513. [Google Scholar] [CrossRef] [Green Version]
- Miyakawa, I.; Sato, H.; Maruyama, Y.; Nakaoka, T. Isolation of the mitochondrial nucleoids from yeast Kluyveromyces lactis and analyses of the nucleoid proteins. J. Gen. Appl. Microbiol. 2003, 49, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Miyakawa, I.; Yawata, K. Purification of an Abf2p-like protein from mitochondrial nucleoids of yeast Pichia jadinii and its role in the packaging of mitochondrial DNA. Antonie van Leeuwenhoek, Int. J. Gen. Mol. Microbiol. 2007, 91, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Višacká, K.; Gerhold, J.M.; Petrovičová, J.; Kinský, S.; Jõers, P.; Nosek, J.; Sedman, J.; Tomáška, Ľ. Novel subfamily of mitochondrial HMG box-containing proteins: Functional analysis of Gcf1p from Candida albicans. Microbiology 2009, 155, 1226–1240. [Google Scholar] [CrossRef] [Green Version]
- Bakkaiova, J.; Arata, K.; Matsunobu, M.; Ono, B.; Aoki, T.; Lajdova, D.; Nebohacova, M.; Nosek, J.; Miyakawa, I.; Tomaska, L. The strictly aerobic yeast Yarrowia lipolytica tolerates loss of a mitochondrial DNA-packaging protein. Eukaryot. Cell 2014, 13, 1143–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyakawa, I.; Kitamura, Y.; Jyozaki, T.; Sato, H.; Umezaki, T. Simple detection of a yeast mitochondrial DNA-binding protein, Abf2p, on SDS-DNA gels. J. Gen. Appl. Microbiol. 2000, 46, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umezaki, T.; Miyakawa, I. Use of SDS-DNA PAGE for detection of mitochondrial Abf2p-like proteins and mitochondrial nuclease in Saccharomyces yeasts and Arxiozyma telluris. Cytologia (Tokyo) 2002, 67, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2017, 46, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Nosek, J.; Dinouël, N.; Kovac, L.; Fukuhara, H. Linear mitochondrial DNAs from yeasts: Telomeres with large tandem repetitions. Mol. Gen. Genet. 1995, 247, 61–72. [Google Scholar] [CrossRef]
- Tomáška, Ľ.; Nosek, J.; Makhov, A.M.; Pastoráková, A.; Griffith, J.D. Extragenomic double-stranded DNA circles in yeast with linear mitochondrial genomes: Potential involvement in telomere maintenance. Nucleic Acids Res. 2000, 28, 4479–4487. [Google Scholar] [CrossRef] [Green Version]
- Nosek, J.; Ryčovská, A.; Makhov, A.M.; Griffith, J.D.; Tomáška, Ľ. Amplification of telomeric arrays via rolling-circle mechanism. J. Biol. Chem. 2005, 280, 10840–10845. [Google Scholar] [CrossRef] [Green Version]
- Christen, S.; Sauer, U. Intracellular characterization of aerobic glucose metabolism in seven yeast species by 13C flux analysis and metabolomics. FEMS Yeast Res. 2011, 11, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Goldring, E.S.; Grossman, L.I.; Krupnick, D.; Cryer, D.R.; Marmur, J. The petite mutation in yeast. Loss of mitochondrial deoxyribonucleic acid during induction of petites with ethidium bromide. J. Mol. Biol. 1970, 52, 323–335. [Google Scholar] [CrossRef]
- Perlman, P.S.; Mahler, H.R. Molecular consequences of ethidium bromide mutagenesis. Nat. New Biol. 1971, 231, 12–16. [Google Scholar] [CrossRef]
- Richter-Dennerlein, R.; Oeljeklaus, S.; Lorenzi, I.; Ronsör, C.; Bareth, B.; Schendzielorz, A.B.; Wang, C.; Warscheid, B.; Rehling, P.; Dennerlein, S. Mitochondrial protein synthesis adapts to influx of nuclear-encoded protein. Cell 2016, 167, 471–483. [Google Scholar] [CrossRef] [Green Version]
- Couvillion, M.T.; Soto, I.C.; Shipkovenska, G.; Churchman, L.S. Synchronized mitochondrial and cytosolic translation programs. Nature 2016, 533, 499–503. [Google Scholar] [CrossRef] [Green Version]
- Suhm, T.; Kaimal, J.M.; Dawitz, H.; Peselj, C.; Masser, A.E.; Hanzén, S.; Ambrožič, M.; Smialowska, A.; Björck, M.L.; Brzezinski, P.; et al. Mitochondrial translation efficiency controls cytoplasmic protein homeostasis. Cell Metab. 2018, 27, 1309–1322. [Google Scholar] [CrossRef] [Green Version]
- Fleck, O.; Kunz, C.; Rudolph, C.; Kohli, J. The high mobility group domain protein Cmb1 of Schizosaccharomyces pombe binds to cytosines in base mismatches and opposite chemically altered guanines. J. Biol. Chem. 1998, 273, 30398–30405. [Google Scholar] [CrossRef] [Green Version]
- Matsuyama, A.; Arai, R.; Yashiroda, Y.; Shirai, A.; Kamata, A.; Sekido, S.; Kobayashi, Y.; Hashimoto, A.; Hamamoto, M.; Hiraoka, Y.; et al. ORFeome cloning and global analysis of protein localization in the fission yeast Schizosaccharomyces pombe. Nat. Biotechnol. 2006, 24, 841–847. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, 585–587. [Google Scholar] [CrossRef] [Green Version]
- Claros, M.G.; Vincens, P. Computational method to predict mitochondrially imported proteins and their targeting sequences. Eur. J. Biochem. 1996, 241, 779–786. [Google Scholar] [CrossRef]
- Reeves, R.; Nissen, M.S. The AT-DNA-binding domain of mammalian high mobility group I chromosomal proteins. J. Biol. Chem. 1990, 265, 8573–8658. [Google Scholar] [PubMed]
- Karácsony, Z.; Gácser, A.; Vágvölgyi, C.; Scazzocchio, C.; Hamari, Z. A dually located multi-HMG-box protein of Aspergillus nidulans has a crucial role in conidial and ascospore germination. Mol. Microbiol. 2014, 94, 383–402. [Google Scholar] [CrossRef]
- Karácsony, Z.; Gácser, A.; Vágvölgyi, C.; Hamari, Z. Further characterization of the role of the mitochondrial high-mobility group box protein in the intracellular redox environment of Aspergillus nidulans. Microbiology (United Kingdom) 2015, 161, 1897–1908. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, N.; Kuroiwa, H.; Nishitani, C.; Takano, H.; Higashiyama, T.; Kobayashi, T.; Shirai, Y.; Sakai, A.; Kawano, S.; Murakami-Murofushi, K.; et al. Glom is a novel mitochondrial DNA packaging protein in Physarum polycephalum and causes intense chromatin condensation without suppressing DNA functions. Mol. Biol. Cell 2003, 14, 4758–4769. [Google Scholar] [CrossRef] [Green Version]
- Itoh, K.; Izumi, A.; Mori, T.; Dohmae, N.; Yui, R.; Maeda-Sano, K.; Shirai, Y.; Kanaoka, M.M.; Kuroiwa, T.; Higashiyama, T.; et al. DNA packaging proteins Glom and Glom2 coordinately organize the mitochondrial nucleoid of Physarum polycephalum. Mitochondrion 2011, 11, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Jensen, R.E.; Englund, P.T. Network news: The replication of kinetoplast DNA. Annu. Rev. Microbiol. 2012, 66, 473–491. [Google Scholar] [CrossRef]
- Xu, C.; Ray, D.S. Isolation of proteins associated with kinetoplast DNA networks in vivo. Proc. Natl. Acad. Sci. USA 1993, 90, 1786–1789. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.W.; Hines, J.C.; Engel, M.L.; Russell, D.G.; Ray, D.S. Nucleus-encoded histone H1-like proteins are associated with kinetoplast DNA in the trypanosomatid Crithidia fasciculata. Mol. Cell. Biol. 1996, 16, 564–576. [Google Scholar] [CrossRef] [Green Version]
- Avliyakulov, N.K.; Lukeš, J.; Ray, D.S. Mitochondrial histone-like DNA-binding proteins are essential for normal cell growth and mitochondrial function in Crithidia fasciculata. Eukaryot. Cell 2004, 3, 518–526. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Pappas-Brown, V.; Englund, P.T.; Jensen, R.E. TbKAP6, a mitochondrial HMG box-containing protein in Trypanosoma brucei, Is the first trypanosomatid kinetoplast-associated protein essential for kinetoplast DNA replication and maintenance. Eukaryot. Cell 2014, 13, 919–932. [Google Scholar] [CrossRef] [Green Version]
- Rajakumara, E.; Satish, M.; Abhishek, S. In vitro studies on non-canonical DNA binding specificities of KAP6 and HMO1 and mechanistic insights into DNA bound and unbinding dynamics of KAP6. Int. J. Biol. Macromol. 2020. [Google Scholar] [CrossRef]
- Fey, J.; Vermel, M.; Grienenberger, J.M.; Maréchal-Drouard, L.; Gualberto, J.M. Characterization of a plant mitochondrial active chromosome. FEBS Lett. 1999, 458, 124–128. [Google Scholar] [CrossRef] [Green Version]
- Dai, H.; Lo, Y.-S.; Litvinchuk, A.; Wang, Y.-T.; Jane, W.-N.; Hsiao, L.-J.; Chiang, K.-S. Structural and functional characterizations of mung bean mitochondrial nucleoids. Nucleic Acids Res. 2005, 33, 4725–4739. [Google Scholar] [CrossRef] [Green Version]
- Sakai, A.; Takano, H.; Kuroiwa, T. Organelle nuclei in higher plants: Structure, composition, function, and evolution. Int. Rev. Cytol. 2004, 238, 59–118. [Google Scholar] [CrossRef]
- Gualberto, J.M.; Kühn, K. DNA-binding proteins in plant mitochondria: Implications for transcription. Mitochondrion 2014, 19, 323–328. [Google Scholar] [CrossRef]
- Tomáška, Ľ.; Nosek, J. Co-evolution in the jungle: From leafcutter ant colonies to chromosomal ends. J. Mol. Evol. 2020, 88, 293–318. [Google Scholar] [CrossRef]
- Tomáška, Ľ.; Nosek, J.; Kramara, J.; Griffith, J.D. Telomeric circles: Universal players in telomere maintenance? Nat. Struct. Mol. Biol. 2009, 16, 1010–1015. [Google Scholar] [CrossRef]
- Ohno, T.; Umeda, S.; Hamasaki, N.; Kang, D. Binding of human mitochondrial transcription factor A, an HMG box protein, to a four-way DNA junction. Biochem. Biophys. Res. Commun. 2000, 271, 492–498. [Google Scholar] [CrossRef]
- Chen, X.J. Mechanism of homologous recombination and implications for aging-related deletions in mitochondrial DNA. Microbiol. Mol. Biol. Rev. 2013, 77, 476–496. [Google Scholar] [CrossRef] [Green Version]
- Pohjoismäki, J.L.O.; Goffart, S.; Tyynismaa, H.; Willcox, S.; Ide, T.; Kang, D.; Suomalainen, A.; Karhunen, P.J.; Griffith, J.D.; Holt, I.J.; et al. Human heart mitochondrial DNA is organized in complex catenated networks containing abundant four-way junctions and replication forks. J. Biol. Chem. 2009, 284, 21446–21457. [Google Scholar] [CrossRef] [Green Version]
- O’Rourke, T.W.; Doudican, N.A.; Mackereth, M.D.; Doetsch, P.W.; Shadel, G.S. Mitochondrial dysfunction due to oxidative mitochondrial DNA damage is reduced through cooperative actions of diverse proteins. Mol. Cell. Biol. 2002, 22, 4086–4093. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, E.A.; Shadel, G.S. Crosstalk between mitochondrial stress signals regulates yeast chronological lifespan. Mech. Ageing Dev. 2014, 135, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Sia, R.A.; Carrol, S.; Kalifa, L.; Hochmuth, C.; Sia, E.A. Loss of the mitochondrial nucleoid protein, Abf2p, destabilizes repetitive DNA in the yeast mitochondrial genome. Genetics 2009, 181, 331–334. [Google Scholar] [CrossRef] [Green Version]
- Chimienti, G.; Picca, A.; Sirago, G.; Fracasso, F.; Calvani, R.; Bernabei, R.; Russo, F.; Carter, C.S.; Leeuwenburgh, C.; Pesce, V.; et al. Increased TFAM binding to mtDNA damage hot spots is associated with mtDNA loss in aged rat heart. Free Radic. Biol. Med. 2018, 124, 447–453. [Google Scholar] [CrossRef]
- Huang, J.C.; Zamble, D.B.; Reardon, J.T.; Lippard, S.J.; Sancar, A. HMG-domain proteins specifically inhibit the repair of the major DNA adduct of the anticancer drug cisplatin by human excision nuclease. Proc. Natl. Acad. Sci. USA 1994, 91, 10394–10398. [Google Scholar] [CrossRef] [Green Version]
- Kazak, L.; Reyes, A.; Holt, I.J. Minimizing the damage: Repair pathways keep mitochondrial DNA intact. Nat. Rev. Mol. Cell Biol. 2012, 13, 659–671. [Google Scholar] [CrossRef]
- Yoshida, Y.; Izumi, H.; Torigoe, T.; Ishiguchi, H.; Itoh, H.; Kang, D.; Kohno, K. p53 physically interacts with mitochondrial transcription factor A and differentially regulates binding to damaged DNA. Cancer Res. 2003, 63, 3729–3734. [Google Scholar]
- Cho, J.H.; Lee, Y.K.; Chae, C.B. The modulation of the biological activities of mitochondrial histone Abf2p by yeast PKA and its possible role in the regulation of mitochondrial DNA content during glucose repression. Biochim. Biophys. Acta—Gene Struct. Expr. 2001, 1522, 175–186. [Google Scholar] [CrossRef]
- Brewer, L.R.; Friddle, R.; Noy, A.; Baldwin, E.; Martin, S.S.; Corzett, M.; Balhorn, R.; Baskin, R.J. Packaging of single DNA molecules by the yeast mitochondrial protein Abf2p. Biophys. J. 2003, 85, 2519–2524. [Google Scholar] [CrossRef] [Green Version]
- Friddle, R.W.; Klare, J.E.; Martin, S.S.; Corzett, M.; Balhorn, R.; Baldwin, E.P.; Baskin, R.J.; Noy, A. Mechanism of DNA compaction by yeast mitochondrial protein Abf2p. Biophys. J. 2004, 86, 1632–1639. [Google Scholar] [CrossRef] [Green Version]
- Stigter, D. Packaging of single DNA molecules by the yeast mitochondrial protein Abf2p: Reinterpretation of recent single molecule experiments. Biophys. Chem. 2004, 110, 171–178. [Google Scholar] [CrossRef]
- Ho, B.; Baryshnikova, A.; Brown, G.W. Unification of protein abundance datasets yields a quantitative Saccharomyces cerevisiae proteome. Cell Syst. 2018, 6, 192–205. [Google Scholar] [CrossRef] [Green Version]
- Davidson, G.S.; Joe, R.M.; Roy, S.; Meirelles, O.; Allen, C.P.; Wilson, M.R.; Tapia, P.H.; Manzanilla, E.E.; Dodson, A.E.; Chakraborty, S.; et al. The proteomics of quiescent and nonquiescent cell differentiation in yeast stationary-phase cultures. Mol. Biol. Cell 2011, 22, 988–998. [Google Scholar] [CrossRef]
- Lahtvee, P.J.; Sánchez, B.J.; Smialowska, A.; Kasvandik, S.; Elsemman, I.E.; Gatto, F.; Nielsen, J. Absolute quantification of protein and mRNA abundances demonstrate variability in gene-specific translation efficiency in yeast. Cell Syst. 2017, 4, 495–504. [Google Scholar] [CrossRef] [Green Version]
- Rafelski, S.M.; Viana, M.P.; Zhang, Y.; Chan, Y.H.M.; Thorn, K.S.; Yam, P.; Fung, J.C.; Li, H.; Costa, L.D.F.; Marshall, W.F. Mitochondrial network size scaling in budding yeast. Science 2012, 338, 822–824. [Google Scholar] [CrossRef] [Green Version]
- Swinger, K.K.; Lemberg, K.M.; Zhang, Y.; Rice, P.A. Flexible DNA bending in HU–DNA cocrystal structures. EMBO J. 2003, 22, 3749–3760. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Im, H.; Jee, J.G.; Jang, S.B.; Yoon, H.J.; Kwon, A.R.; Kang, S.M.; Lee, B.J. β-Arm flexibility of HU from Staphylococcus aureus dictates the DNA-binding and recognition mechanism. Acta Crystallogr. Sect. D Biol. Crystallogr. 2014, 70, 3273–3289. [Google Scholar] [CrossRef]
- Hammel, M.; Amlanjyoti, D.; Reyes, F.E.; Chen, J.H.; Parpana, R.; Tang, H.Y.H.; Larabell, C.A.; Tainer, J.A.; Adhya, S. HU multimerization shift controls nucleoid compaction. Sci. Adv. 2016, 2, e1600650. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, A.; Lyonnais, S.; Battistini, F.; Hospital, A.; Medici, G.; Prohens, R.; Orozco, M.; Vilardell, J.; Solà, M. DNA structure directs positioning of the mitochondrial genome packaging protein Abf2p. Nucleic Acids Res. 2017, 45, 951–967. [Google Scholar] [CrossRef] [Green Version]
- Lyonnais, S.; Tarrés-Soler, A.; Rubio-Cosials, A.; Cuppari, A.; Brito, R.; Jaumot, J.; Gargallo, R.; Vilaseca, M.; Silva, C.; Granzhan, A.; et al. The human mitochondrial transcription factor A is a versatile G-quadruplex binding protein. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Reeves, R. High mobility group (HMG) proteins: Modulators of chromatin structure and DNA repair in mammalian cells. DNA Repair (Amst) 2015, 36, 122–136. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Cosials, A.; Battistini, F.; Gansen, A.; Cuppari, A.; Bernadó, P.; Orozco, M.; Langowski, J.; Tóth, K.; Solà, M. Protein flexibility and synergy of HMG domains underlie U-turn bending of DNA by TFAM in solution. Biophys. J. 2018, 114, 2386–2396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaspari, M.; Falkenberg, M.; Larsson, N.-G.; Gustafsson, C.M. The mitochondrial RNA polymerase contributes critically to promoter specificity in mammalian cells. EMBO J. 2004, 23, 4606–4614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio-Cosials, A.; Sidow, J.F.; Jiménez-Menéndez, N.; Fernández-Millán, P.; Montoya, J.; Jacobs, H.T.; Coll, M.; Bernadó, P.; Solà, M. Human mitochondrial transcription factor A induces a U-turn structure in the light strand promoter. Nat. Struct. Mol. Biol. 2011, 18, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- Ngo, H.B.; Kaiser, J.T.; Chan, D.C. The mitochondrial transcription and packaging factor Tfam imposes a U-turn on mitochondrial DNA. Nat. Struct. Mol. Biol. 2011, 18, 1290–1296. [Google Scholar] [CrossRef] [Green Version]
- Ngo, H.B.; Lovely, G.A.; Phillips, R.; Chan, D.C. Distinct structural features of TFAM drive mitochondrial DNA packaging versus transcriptional activation. Nat. Commun. 2014, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kukat, C.; Davies, K.M.; Wurm, C.A.; Spåhr, H.; Bonekamp, N.A.; Kühl, I.; Joos, F.; Polosa, P.L.; Park, C.B.; Posse, V.; et al. Cross-strand binding of TFAM to a single mtDNA molecule forms the mitochondrial nucleoid. Proc. Natl. Acad. Sci. USA 2015, 112, 11288–11293. [Google Scholar] [CrossRef] [Green Version]
- Gangelhoff, T.A.; Mungalachetty, P.S.; Nix, J.C.; Churchill, M.E.A. Structural analysis and DNA binding of the HMG domains of the human mitochondrial transcription factor A. Nucleic Acids Res. 2009, 37, 3153–3164. [Google Scholar] [CrossRef] [Green Version]
- Dorman, C.J. Function of nucleoid-associated proteins in chromosome structuring and transcriptional regulation. J. Mol. Microbiol. Biotechnol. 2014, 24, 316–331. [Google Scholar] [CrossRef] [Green Version]
- Krogh, T.J.; Møller-Jensen, J.; Kaleta, C. Impact of chromosomal architecture on the function and evolution of bacterial genomes. Front. Microbiol. 2018, 9, 2019. [Google Scholar] [CrossRef]
- Atlung, T.; Ingmer, H. H-NS: A modulator of environmentally regulated gene expression. Mol. Microbiol. 1997, 24, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Talukder, A.A.; Ishihama, A. Growth phase dependent changes in the structure and protein composition of nucleoid in Escherichia coli. Sci. China Life Sci. 2015, 58, 902–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Colman, M.J.; Reverter-Branchat, G.; Sorolla, M.A.; Tamarit, J.; Ros, J.; Cabiscol, E. The forkhead transcription factor Hcm1 promotes mitochondrial biogenesis and stress resistance in yeast. J. Biol. Chem. 2010, 285, 37092–37101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venters, B.J.; Wachi, S.; Mavrich, T.N.; Andersen, B.E.; Jena, P.; Sinnamon, A.J.; Jain, P.; Rolleri, N.S.; Jiang, C.; Hemeryck-Walsh, C.; et al. A comprehensive genomic binding map of gene and chromatin regulatory proteins in Saccharomyces. Mol. Cell 2011, 41, 480–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corral-Debrinski, M.; Blugeon, C.; Jacq, C. In yeast, the 3′ untranslated region or the presequence of ATM1 is required for the exclusive localization of its mRNA to the vicinity of mitochondria. Mol. Cell. Biol. 2000, 20, 7881–7892. [Google Scholar] [CrossRef] [PubMed]
- Bandhakavi, S.; Xie, H.; O’Callaghan, B.; Sakurai, H.; Kim, D.H.; Griffin, T.J. Hsf1 activation inhibits rapamycin resistance and TOR signaling in yeast revealed by combined proteomic and genetic analysis. PLoS ONE 2008, 3, e1598. [Google Scholar] [CrossRef] [Green Version]
- Bonawitz, N.D.; Chatenay-Lapointe, M.; Pan, Y.; Shadel, G.S. Reduced TOR signaling extends chronological life span via increased respiration and upregulation of mitochondrial gene expression. Cell Metab. 2007, 5, 265–277. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, M.; Naka, A.; Sakamoto, Y.; Shibasaki, A.; Toh, M.; Tsukamoto, S.; Kondo, K.; Iida, K. Dietary isoflavone daidzein promotes Tfam expression that increases mitochondrial biogenesis in C2C12 muscle cells. J. Nutr. Biochem. 2015, 26, 1193–1199. [Google Scholar] [CrossRef]
- Virbasius, J.V.; Scarpulla, R.C. Activation of the human mitochondrial transcription factor A gene by nuclear respiratory factors: A potential regulatory link between nuclear and mitochondrial gene expression in organelle biogenesis. Proc. Natl. Acad. Sci. USA 1994, 91, 1309–1313. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Li, S.; Ke, Y.; Wu, S.; Huang, T.; Hu, W.; Fu, H.; Guo, X. KLF16 suppresses human glioma cell proliferation and tumourigenicity by targeting TFAM. Artif. Cells Nanomedicine Biotechnol. 2018, 46, 608–615. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Guo, T.; Chen, Z.; Zhang, H.; Cai, S.; Yang, M.; Chen, P.; Guan, C.; Fang, X. Hypermethylation of mitochondrial transcription factor A induced by cigarette smoke is associated with chronic obstructive pulmonary disease. Exp. Lung Res. 2019, 45, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Larsson, N.G.; Garman, J.D.; Oldfors, A.; Barsh, G.S.; Clayton, D.A. A single mouse gene encodes the mitochondrial transcription factor A and a testis-specific nuclear HMG-box protein. Nat. Genet. 1996, 13, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Larsson, N.-G.; Oldfors, A.; David Garman, J.; Barsh, G.S.; Clayton, D.A. Down-regulation of mitochondrial transcription factor A during spermatogenesis in humans. Hum. Mol. Genet. 1997, 6, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wang, J. HuR stabilizes TFAM mRNA in an ATM/p38-dependent manner in ionizing irradiated cancer cells. Cancer Sci. 2018, 109, 2446–2457. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Yang, J.; Wang, Z.; Wu, G.; Liu, F. TFAM is directly regulated by miR-23b in glioma. Oncol. Rep. 2013, 30, 2105–2110. [Google Scholar] [CrossRef]
- Wen, Z.; Lei, Z.; Jin-An, M.; Xue-Zhen, M.; Xing-Nan, Z.; Xiu-Wen, D. The inhibitory role of miR-214 in cervical cancer cells through directly targeting mitochondrial transcription factor A (TFAM). Eur. J. Gynaecol Oncol 2014, 35, 676–682. [Google Scholar] [CrossRef]
- Wu, K.; Zhao, Z.; Xiao, Y.; Peng, J.; Chen, J.; He, Y. Roles of mitochondrial transcription factor A and microRNA-590-3p in the development of colon cancer. Mol. Med. Rep. 2016, 14, 5475–5480. [Google Scholar] [CrossRef] [Green Version]
- Mo, M.; Peng, F.; Wang, L.; Peng, L.; Lan, G.; Yu, S. Roles of mitochondrial transcription factor A and microRNA-590-3p in the development of bladder cancer. Oncol. Lett. 2013, 6, 617–623. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, H.; Morino, K.; Nishio, Y.; Ugi, S.; Yoshizaki, T.; Kashiwagi, A.; Maegawa, H. MicroRNA-494 regulates mitochondrial biogenesis in skeletal muscle through mitochondrial transcription factor A and Forkhead box j3. Am. J. Physiol—Endocrinol. Metab. 2012, 303. [Google Scholar] [CrossRef] [Green Version]
- Lemecha, M.; Morino, K.; Imamura, T.; Iwasaki, H.; Ohashi, N.; Ida, S.; Sato, D.; Sekine, O.; Ugi, S.; Maegawa, H. MiR-494-3p regulates mitochondrial biogenesis and thermogenesis through PGC1-α signalling in beige adipocytes. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Cui, D.; Zhang, Z.; Zhang, Q.; Ji, L.; Ding, S. Voluntary wheel exercise alters the levels of miR-494 and miR-696 in the skeletal muscle of C57BL/6 mice. Comp. Biochem. Physiol. Part—B Biochem. Mol. Biol. 2016, 202, 16–22. [Google Scholar] [CrossRef]
- Bruno, S.; De Virgilio, C.; Gadaleta, G. The smaller isoform of the mitochondrial transcription factor A has a role in the mitochondrial transcription. Ital. J. Biochem. 2007, 56, 315–318. [Google Scholar] [PubMed]
- Reyes, A.; Mezzina, M.; Gadaleta, G. Human mitochondrial transcription factor A (mtTFA): Gene structure and characterization of related pseudogenes. Gene 2002, 291, 223–232. [Google Scholar] [CrossRef]
- Tominaga, K.; Hayashi, J.I.; Kagawa, Y.; Ohta, S. Smaller isoform of human mitochondrial transcription factor 1: Its wide distribution and production by alternative splicing. Biochem. Biophys. Res. Commun. 1993, 194, 544–551. [Google Scholar] [CrossRef] [PubMed]
- De Virgilio, C.; Pousis, C.; Bruno, S.; Gadaleta, G. New isoforms of human mitochondrial transcription factor A detected in normal and tumoral cells. Mitochondrion 2011, 11, 287–295. [Google Scholar] [CrossRef]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [Green Version]
- Tessarz, P.; Kouzarides, T. Histone core modifications regulating nucleosome structure and dynamics. Nat. Rev. Mol. Cell Biol. 2014, 15, 703–708. [Google Scholar] [CrossRef]
- Macek, B.; Mijakovic, I.; Olsen, J.V.; Gnad, F.; Kumar, C.; Jensen, P.R.; Mann, M. The serine/threonine/tyrosine phosphoproteome of the model bacterium Bacillus subtilis. Mol. Cell. Proteomics 2007, 6, 697–707. [Google Scholar] [CrossRef] [Green Version]
- Soufi, B.; Jers, C.; Hansen, M.E.; Petranovic, D.; Mijakovic, I. Insights from site-specific phosphoproteomics in bacteria. Biochim. Biophys. Acta—Proteins Proteomics 2008, 1784, 186–192. [Google Scholar] [CrossRef]
- Yu, J.B.; Kim, J.A.; Moon, J.H.; Pan, S.E.R.J. The diversity of lysine-acetylated proteins in Escherichia coli. J. Microbiol. Biotechnol. 2008, 18, 1529–1536. [Google Scholar]
- Cao, X.J.; Dai, J.; Xu, H.; Nie, S.; Chang, X.; Hu, B.Y.; Sheng, Q.H.; Wang, L.S.; Ning, Z.B.; Li, Y.X.; et al. High-coverage proteome analysis reveals the first insight of protein modification systems in the pathogenic spirochete Leptospira interrogans. Cell Res. 2010, 20, 197–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dilweg, I.W.; Dame, R.T. Post-translational modification of nucleoid-associated proteins: An extra layer of functional modulation in bacteria? Biochem. Soc. Trans. 2018, 46, 1381–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, B.; Lee, J.; Nie, X.; Li, M.; Morozov, Y.I.; Venkatesh, S.; Bogenhagen, D.F.; Temiakov, D.; Suzuki, C.K. Phosphorylation of human TFAM in mitochondria impairs DNA binding and promotes degradation by the AAA+ Lon protease. Mol. Cell 2013, 49, 121–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.Z.Q.; Zhu, J.; Dagda, R.K.; Uechi, G.; Cherra, S.J.; Gusdon, A.M.; Balasubramani, M.; Chu, C.T. ERK-mediated phosphorylation of TFAM downregulates mitochondrial transcription: Implications for Parkinson’s disease. Mitochondrion 2014, 17, 132–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunová, N.; Ondrovičová, G.; Bauer, J.A.; Bellová, J.; Ambro, L.; Martináková, L.; Kotrasová, V.; Kutejová, E.; Pevala, V. The role of Lon-mediated proteolysis in the dynamics of mitochondrial nucleic acid-protein complexes. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Chen, R.Q.; Yang, Q.K.; Lu, B.W.; Yi, W.; Cantin, G.; Chen, Y.L.; Fearns, C.; Yates, J.R.; Lee, J.D. CDC25B mediates rapamycin-induced oncogenic responses in cancer cells. Cancer Res. 2009, 69, 2663–2668. [Google Scholar] [CrossRef] [Green Version]
- Dinardo, M.M.; Musicco, C.; Fracasso, F.; Milella, F.; Gadaleta, M.N.; Gadaleta, G.; Cantatore, P. Acetylation and level of mitochondrial transcription factor A in several organs of young and old rats. Biochem. Biophys. Res. Commun. 2003, 301, 187–191. [Google Scholar] [CrossRef]
- King, G.A.; Hashemi Shabestari, M.; Taris, K.K.H.; Pandey, A.K.; Venkatesh, S.; Thilagavathi, J.; Singh, K.; Krishna Koppisetti, R.; Temiakov, D.; Roos, W.H.; et al. Acetylation and phosphorylation of human TFAM regulate TFAM-DNA interactions via contrasting mechanisms. Nucleic Acids Res. 2018, 46, 3633–3642. [Google Scholar] [CrossRef]
- Fang, Y.; Akimoto, M.; Mayanagi, K.; Hatano, A.; Matsumoto, M.; Matsuda, S.; Yasukawa, T.; Kang, D. Chemical acetylation of mitochondrial transcription factor A occurs on specific lysine residues and affects its ability to change global DNA topology. Mitochondrion 2020, 53, 99–108. [Google Scholar] [CrossRef]
- Liu, H.; Li, S.; Liu, X.; Chen, Y.; Deng, H. SIRT3 overexpression inhibits growth of kidney tumor cells and enhances mitochondrial biogenesis. J. Proteome Res. 2018, 17, 3143–3152. [Google Scholar] [CrossRef]
- Henriksen, P.; Wagner, S.A.; Weinert, B.T.; Sharma, S.; Bačinskaja, G.; Rehman, M.; Juffer, A.H.; Walther, T.C.; Lisby, M.; Choudhary, C. Proteome-wide analysis of lysine acetylation suggests its broad regulatory scope in Saccharomyces cerevisiae. Mol. Cell. Proteomics 2012, 11, 1510–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinert, B.T.; Schölz, C.; Wagner, S.A.; Iesmantavicius, V.; Su, D.; Daniel, J.A.; Choudhary, C. Lysine succinylation is a frequently occurring modification in prokaryotes and eukaryotes and extensively overlaps with acetylation. Cell Rep. 2013, 4, 842–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, J.M.; Mishra, M.; Kowluru, R.A. Posttranslational modification of mitochondrial transcription factor A in impaired mitochondria biogenesis: Implications in diabetic retinopathy and metabolic memory phenomenon. Exp. Eye Res. 2014, 121, 168–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Chan, D.W.; Jung, S.Y.; Malovannaya, A.; Wang, Y.; Qin, J. A data set of human endogenous protein ubiquitination sites. Mol. Cell. Proteomics 2011, 10. [Google Scholar] [CrossRef] [Green Version]
- Wagner, S.A.; Beli, P.; Weinert, B.T.; Nielsen, M.L.; Cox, J.; Mann, M.; Choudhary, C. A Proteome-wide, quantitative survey of in vivo ubiquitylation sites reveals widespread regulatory roles. Mol. Cell. Proteomics 2011, 10, M111.013284. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.R.C.; Dvoriantchikova, G.; Li, Y.; Mohammad, G.; Abu El-Asrar, A.M.; Wen, R.; Ivanov, D. Cellular mechanisms of High Mobility Group 1 (HMGB-1) protein action in the diabetic retinopathy. PLoS ONE 2014, 9, e87574. [Google Scholar] [CrossRef]
- Suarez, J.; Hu, Y.; Makino, A.; Fricovsky, E.; Wang, H.; Dillmann, W.H. Alterations in mitochondrial function and cytosolic calcium induced by hyperglycemia are restored by mitochondrial transcription factor A in cardiomyocytes. Am. J. Physiol—Cell Physiol. 2008, 295. [Google Scholar] [CrossRef] [Green Version]
- El-Hattab, A.W.; Craigen, W.J.; Scaglia, F. Mitochondrial DNA maintenance defects. Biochim. Biophys. Acta—Mol. Basis Dis. 2017, 1863, 1539–1555. [Google Scholar] [CrossRef]
- Suomalainen, A.; Isohanni, P. Mitochondrial DNA depletion syndromes: Many genes, common mechanisms. Neuromuscul. Disord. 2010, 20, 429–437. [Google Scholar] [CrossRef]
- Woo, D.K.; Green, P.D.; Santos, J.H.; D’Souza, A.D.; Walther, Z.; Martin, W.D.; Christian, B.E.; Chandel, N.S.; Shadel, G.S. Mitochondrial genome instability and ROS enhance intestinal tumorigenesis in APCMin/+ mice. Am. J. Pathol. 2012, 180, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Desdín-Micó, G.; Soto-Heredero, G.; Aranda, J.F.; Oller, J.; Carrasco, E.; Gabandé-Rodríguez, E.; Blanco, E.M.; Alfranca, A.; Cussó, L.; Desco, M.; et al. T cells with dysfunctional mitochondria induce multimorbidity and premature senescence. Science 2020, 368, 1371–1376. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Silva, J.P.; Gustafsson, C.M.; Rustin, P.; Larsson, N.G. Increased in vivo apoptosis in cells lacking mitochondrial DNA gene expression. Proc. Natl. Acad. Sci. USA 2001, 98, 4038–4043. [Google Scholar] [CrossRef] [Green Version]
- Aydin, J.; Andersson, D.C.; Hä Nninen, S.L.; Wredenberg, A.; Tavi, P.; Park, C.B.; Ran Larsson, N.-G.; Bruton, J.D.; Kan Westerblad, H. Increased mitochondrial Ca2+ and decreased sarcoplasmic reticulum Ca2+ in mitochondrial myopathy. Hum. Mol. Genet. 2009, 18, 278–288. [Google Scholar] [CrossRef] [Green Version]
- Farge, G.; Mehmedovic, M.; Baclayon, M.; VandenWildenberg, S.M.J.L.; Roos, W.H.; Gustafsson, C.M.; Wuite, G.J.L.; Falkenberg, M. In vitro-reconstituted nucleoids can block mitochondrial DNA replication and transcription. Cell Rep. 2014, 8, 66–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, I.; Chu, C.T.; Kaufman, B.A. The mitochondrial transcription factor TFAM in neurodegeneration: Emerging evidence and mechanisms. FEBS Lett. 2018, 592, 793–811. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, M.; Ide, T.; Fujino, T.; Arai, S.; Saku, K.; Kakino, T.; Tyynismaa, H.; Yamasaki, T.; Yamada, K.; Kang, D.; et al. Overexpression of TFAM or Twinkle increases mtDNA copy number and facilitates cardioprotection associated with limited mitochondrial oxidative stress. PLoS ONE 2015, 10, e0119687. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, G.H.; Kunkel, C.J.; Ozuna, H.; Miralda, I.; Tyagi, S.C. TFAM overexpression reduces pathological cardiac remodeling. Mol. Cell. Biochem. 2019, 454, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Picca, A.; Sirago, G.; Pesce, V.; Lezza, A.M.S.; Calvani, R.; Bossola, M.; Villani, E.R.; Landi, F.; Leeuwenburgh, C.; Bernabei, R.; et al. Administration of enalapril started late in life attenuates hypertrophy and oxidative stress burden, increases mitochondrial mass, and modulates mitochondrial quality control signaling in the rat heart. Biomolecules 2018, 8, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiles, A.R.; Simon, M.T.; Stover, A.; Eftekharian, S.; Khanlou, N.; Wang, H.L.; Magaki, S.; Lee, H.; Partynski, K.; Dorrani, N.; et al. Mutations in TFAM, encoding mitochondrial transcription factor A, cause neonatal liver failure associated with mtDNA depletion. Mol. Genet. Metab. 2016, 119, 91–99. [Google Scholar] [CrossRef]
- Arduini, A.; Serviddio, G.; Escobar, J.; Tormos, A.M.; Bellanti, F.; Viña, J.; Monsalve, M.; Sastre, J. Mitochondrial biogenesis fails in secondary biliary cirrhosis in rats leadingto mitochondrial DNA depletion and deletions. Am. J. Physiol—Gastrointest. Liver Physiol. 2011, 301. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.C.; Chen, Y.B.; Lin, H.; Pi, H.F.; Zhang, N.X.; Zhao, C.C.; Shuai, L.; Zhong, M.; Yu, Z.P.; Zhou, Z.; et al. Damage to mtDNA in liver injury of patients with extrahepatic cholestasis: The protective effects of mitochondrial transcription factor A. Free Radic. Biol. Med. 2012, 52, 1543–1551. [Google Scholar] [CrossRef] [PubMed]
- Tiao, M.M.; Lin, T.K.; Liou, C.W.; Wang, P.W.; Chen, J.B.; Kuo, F.Y.; Huang, C.C.; Chou, Y.M.; Chuang, J.H. Early transcriptional deregulation of hepatic mitochondrial biogenesis and its consequent effects on murine cholestatic liver injury. Apoptosis 2009, 14, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Poulton, J.; Morten, K.; Freman-Emmerson, C.; Potter, C.; Sewry, C.; Dubowitz, V.; Kidd, H.; Stephenson, J.; Whitehouse, W.; Hansen, F.J.; et al. Deficiency of the human mitochondrial transcription factor h-mtTFA in infantile mitochondrial myopathy is associated with mtDNA depletion. Hum. Mol. Genet. 1994, 3, 1763–1769. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Azuma, H.; Aihara, K.I.; Fujimura, M.; Akaike, M.; Mitsui, T.; Matsumoto, T. Vascular smooth muscle cell proliferation is dependent upon upregulation of mitochondrial transcription factor A (mtTFA) expression in injured rat carotid artery. Atherosclerosis 2005, 178, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Remels, A.H.; Schrauwen, P.; Broekhuizen, R.; Willems, J.; Kersten, S.; Gosker, H.R.; Schols, A.M. Peroxisome proliferator-activated receptor expression is reduced in skeletal muscle in COPD. Eur. Respir. J. 2007, 30, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Yang, M.; Chen, Z.Y.; Chen, P.; Guan, C.X.; Xiang, X.D.; Cai, S.; Chen, Y.; Fang, X. Expression and methylation of mitochondrial transcription factor a in chronic obstructive pulmonary disease patients with lung cancer. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Lee, W.R.; Na, H.; Lee, S.W.; Lim, W.J.; Kim, N.; Lee, J.E.; Kang, C. Transcriptomic analysis of mitochondrial TFAM depletion changing cell morphology and proliferation. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Zheng, L.; Liu, W.; Wang, X.; Wang, Z.; Wang, Z.; French, A.J.; Kang, D.; Chen, L.; Thibodeau, S.N.; et al. Frequent truncating mutation of TFAM induces mitochondrial DNA depletion and apoptotic resistance in microsatellite-unstable colorectal cancer. Cancer Res. 2011, 71, 2978–2987. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.; Ma, J.; Zhan, Y.; Liu, K.; Ye, Z.; Chen, J.; Xu, K.; Huang, H.; He, Y. Down-regulation of microRNA-214 contributed to the enhanced mitochondrial transcription factor A and inhibited proliferation of colorectal cancer cells. Cell. Physiol. Biochem. 2018, 49, 545–554. [Google Scholar] [CrossRef]
- Yoshida, Y.; Hasegawa, J.; Nezu, R.; Kim, Y.K.; Hirota, M.; Kawano, K.; Izumi, H.; Kohno, K. Clinical usefulness of mitochondrial transcription factor A expression as a predictive marker in colorectal cancer patients treated with FOLFOX. Cancer Sci. 2011, 102, 578–582. [Google Scholar] [CrossRef]
- Lin, C.S.; Lee, H.T.; Lee, M.H.; Pan, S.C.; Ke, C.Y.; Chiu, A.W.H.; Wei, Y.H. Role of mitochondrial DNA copy number alteration in human renal cell carcinoma. Int. J. Mol. Sci. 2016, 17, 814. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.; Wu, X.; Lan, L.; Shangguan, F.; Lin, X.; Chen, F.; Xu, S.; Zhang, Y.; Chen, Z.; Huang, K.; et al. Downregulation of TFAM inhibits the tumorigenesis of non-small cell lung cancer by activating ROS-mediated JNK/p38MAPK signaling and reducing cellular bioenergetics. Oncotarget 2016, 7, 11609–11624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattingly, K.A.; Ivanova, M.M.; Riggs, K.A.; Wickramasinghe, N.S.; Barch, M.J.; Klinge, C.M. Estradiol stimulates transcription of nuclear respiratory factor-1 and increases mitochondrial biogenesis. Mol. Endocrinol. 2008, 22, 609–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Garcia, S.; Prado-Garcia, H.; Valencia-Camargo, A.D.; Alvarez-Pulido, A. Lactic acidosis promotes mitochondrial biogenesis in lung adenocarcinoma cells, supporting proliferation under normoxia or survival under hypoxia. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Wang, Q.; Chen, N.; Sun, Y.; Wang, X.; Wu, L.; Chen, S.; Yuan, H.; Xu, A.; Wang, J. Mitochondrial transcription factor A regulated ionizing radiation-induced mitochondrial biogenesis in human lung adenocarcinoma A549 cells. J. Radiat. Res. 2013, 54, 998–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellance, N.; Benard, G.; Furt, F.; Begueret, H.; Smolková, K.; Passerieux, E.; Delage, J.P.; Baste, J.M.; Moreau, P.; Rossignol, R. Bioenergetics of lung tumors: Alteration of mitochondrial biogenesis and respiratory capacity. Int. J. Biochem. Cell Biol. 2009, 41, 2566–2577. [Google Scholar] [CrossRef]
- Lee, H.; Park, J.; Tran, Q.; Kim, D.; Hong, Y.; Cho, H.; Kwon, S.H.; Brazil, D.; Kim, S.H. Mitochondrial transcription factor a (TFAM) is upregulated in glioma. Mol. Med. Rep. 2017, 15, 3781–3786. [Google Scholar] [CrossRef]
- Franco, D.G.; Moretti, I.F.; Marie, S.K.N. Mitochondria transcription factor A: A putative target for the effect of melatonin on U87MG malignant glioma cell line. Molecules 2018, 23, 1129. [Google Scholar] [CrossRef] [Green Version]
- Correia, R.L.; Oba-Shinjo, S.M.; Uno, M.; Huang, N.; Marie, S.K.N. Mitochondrial DNA depletion and its correlation with TFAM, TFB1M, TFB2M and POLG in human diffusely infiltrating astrocytomas. Mitochondrion 2011, 11, 48–53. [Google Scholar] [CrossRef]
- Lee, C.H.; Wu, S.B.; Hong, C.H.; Liao, W.T.; Wu, C.Y.; Chen, G.S.; Wei, Y.H.; Yu, H.S. Aberrant cell proliferation by enhanced mitochondrial biogenesis via mtTFA in arsenical skin cancers. Am. J. Pathol. 2011, 178, 2066–2076. [Google Scholar] [CrossRef] [Green Version]
- Araujo, L.F.; Siena, A.D.D.; Plaça, J.R.; Brotto, D.B.; Barros, I.I.; Muys, B.R.; Biagi, C.A.O.; Peronni, K.C.; Sousa, J.F.; Molfetta, G.A.; et al. Mitochondrial transcription factor A (TFAM) shapes metabolic and invasion gene signatures in melanoma. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.P.; Kumari, R.; Treas, J.; Dumond, J.W. Chronic exposure to arsenic causes increased cell survival, DNA damage, and increased expression of mitochondrial transcription factor A (mtTFA) in human prostate epithelial cells. Chem. Res. Toxicol. 2011, 24, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Kurita, T.; Izumi, H.; Kagami, S.; Kawagoe, T.; Toki, N.; Matsuura, Y.; Hachisuga, T.; Kohno, K. Mitochondrial transcription factor A regulates BCL2L1 gene expression and is a prognostic factor in serous ovarian cancer. Cancer Sci. 2012, 103, 239–444. [Google Scholar] [CrossRef] [PubMed]
- Toki, N.; Kagami, S.; Kurita, T.; Kawagoe, T.; Matsuura, Y.; Hachisuga, T.; Matsuyama, A.; Hashimoto, H.; Izumi, H.; Kohno, K. Expression of mitochondrial transcription factor A in endometrial carcinomas: Clinicopathologic correlations and prognostic significance. Virchows Arch. 2010, 456, 387–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cormio, A.; Guerra, F.; Cormio, G.; Pesce, V.; Fracasso, F.; Loizzi, V.; Cantatore, P.; Selvaggi, L.; Gadaleta, M.N. The PGC-1α-dependent pathway of mitochondrial biogenesis is upregulated in type I endometrial cancer. Biochem. Biophys. Res. Commun. 2009, 390, 1182–1185. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Wu, M.; Wang, N.; Zhang, Y.; Hua, J.; Tang, G.; Wang, Y. Increased expression of mitochondrial transcription factor a and nuclear respiratory factor-1 predicts a poor clinical outcome of breast cancer. Oncol. Lett. 2018, 15, 1449–1458. [Google Scholar] [CrossRef]
- Gao, W.; Wu, M.H.; Wang, N.; Ying, M.Z.; Zhang, Y.Y.; Hua, J.; Chuan, L.; Wang, Y.J. Mitochondrial transcription factor A contributes to cisplatin resistance in patients with estrogen receptor-positive breast cancer. Mol. Med. Rep. 2016, 14, 5304–5310. [Google Scholar] [CrossRef] [Green Version]
- Qiao, L.; Ru, G.; Mao, Z.; Wang, C.; Nie, Z.; Li, Q.; Huang-yang, Y.; Zhu, L.; Liang, X.; Yu, J.; et al. Mitochondrial DNA depletion, mitochondrial mutations and high TFAM expression in hepatocellular carcinoma. Oncotarget 2017, 8, 84373–84383. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.S.; Lee, H.T.; Lee, S.Y.; Shen, Y.A.; Wang, L.S.; Chen, Y.J.; Wei, Y.H. High mitochondrial DNA copy number and bioenergetic function are associated with tumor invasion of esophageal squamous cell carcinoma cell lines. Int. J. Mol. Sci. 2012, 13, 11228–11246. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.S.; Chang, S.C.; Wang, L.S.; Chou, T.Y.; Hsu, W.H.; Wu, Y.C.; Wei, Y.H. The role of mitochondrial DNA alterations in esophageal squamous cell carcinomas. J. Thorac. Cardiovasc. Surg. 2010, 139. [Google Scholar] [CrossRef] [Green Version]
- Masuike, Y.; Tanaka, K.; Makino, T.; Yamasaki, M.; Miyazaki, Y.; Takahashi, T.; Kurokawa, Y.; Nakajima, K.; Mori, M.; Doki, Y. Esophageal squamous cell carcinoma with low mitochondrial copy number has mesenchymal and stem-like characteristics, and contributes to poor prognosis. PLoS ONE 2018, 13, e0193159. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Ju, Y.S.; Kim, Y.; Li, J.; Wang, Y.; Yoon, C.J.; Yang, Y.; Martincorena, I.; Creighton, C.J.; Weinstein, J.N.; et al. Comprehensive molecular characterization of mitochondrial genomes in human cancers. Nat. Genet. 2020, 52, 342–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Zhan, L.; Chen, Y.; Wang, G.; He, L.; Wang, Q.; Zhou, F.; Yang, F.; Wu, J.; Wu, Y.; et al. Increased mtDNA copy number promotes cancer progression by enhancing mitochondrial oxidative phosphorylation in microsatellite-stable colorectal cancer. Signal. Transduct. Target. Ther. 2018, 3, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Montes, C.; Castro, M.G.; Reguero, J.R.; Perrot, A.; Ozcelik, C.; Geier, C.; Posch, M.G.; Morís, C.; Alvarez, V.; Ruiz-Ortega, M.; et al. Mitochondrial transcription factors TFA, TFB1 and TFB2: A search for DNA variants/haplotypes and the risk of cardiac hypertrophy. Dis. Markers 2008, 25, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Li, G.D.; Fan, Y.; Zhang, D.F.; Bi, R.; Yu, X.F.; Long, H.; Li, Y.Y.; Yao, Y.G. The mtDNA replication-related genes TFAM and POLG are associated with leprosy in Han Chinese from Southwest China. J. Dermatol. Sci. 2017, 88, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Han, B.; Izumi, H.; Yasuniwa, Y.; Akiyama, M.; Yamaguchi, T.; Fujimoto, N.; Matsumoto, T.; Wu, B.; Tanimoto, A.; Sasaguri, Y.; et al. Human mitochondrial transcription factor A functions in both nuclei and mitochondria and regulates cancer cell growth. Biochem. Biophys. Res. Commun. 2011, 408, 45–51. [Google Scholar] [CrossRef]
- Kunkel, G.H.; Chaturvedi, P.; Thelian, N.; Nair, R.; Tyagi, S.C. Mechanisms of TFAM-mediated cardiomyocyte protection. Can. J. Physiol. Pharmacol. 2018, 96, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Ikeuchi, M.; Matsusaka, H.; Kang, D.; Matsushima, S.; Ide, T.; Kubota, T.; Fujiwara, T.; Hamasaki, N.; Takeshita, A.; Sunagawa, K.; et al. Overexpression of mitochondrial transcription factor A ameliorates mitochondrial deficiencies and cardiac failure after myocardial infarction. Circulation 2005, 112, 683–690. [Google Scholar] [CrossRef] [Green Version]
- Fujino, T.; Ide, T.; Yoshida, M.; Onitsuka, K.; Tanaka, A.; Hata, Y.; Nishida, M.; Takehara, T.; Kanemaru, T.; Kitajima, N.; et al. Recombinant mitochondrial transcription factor A protein inhibits nuclear factor of activated T cells signaling and attenuates pathological hypertrophy of cardiac myocytes. Mitochondrion 2012, 12, 449–458. [Google Scholar] [CrossRef]
- Theilen, N.T.; Kunkel, G.H.; Tyagi, S.C. The role of exercise and TFAM in preventing skeletal muscle atrophy. J. Cell. Physiol. 2017, 232, 2348–2358. [Google Scholar] [CrossRef]
- Chandrasekaran, K.; Muragundla, A.; Demarest, T.G.; Choi, J.; Sagi, A.R.; Najimi, N.; Kumar, P.; Singh, A.; Ho, C.Y.; Fiskum, G.; et al. mGluR2/3 activation of the SIRT1 axis preserves mitochondrial function in diabetic neuropathy. Ann. Clin. Transl. Neurol. 2017, 4, 844–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrasekaran, K.; Anjaneyulu, M.; Choi, J.; Kumar, P.; Salimian, M.; Ho, C.Y.; Russell, J.W. Role of mitochondria in diabetic peripheral neuropathy: Influencing the NAD + -dependent SIRT1–PGC-1α–TFAM pathway. In International Review of Neurobiology; Academic Press Inc.: Cambridge, MA, USA, 2019; Volume 145, pp. 177–209. [Google Scholar]
- Keeney, P.M.; Quigley, C.K.; Dunham, L.D.; Papageorge, C.M.; Iyer, S.; Thomas, R.R.; Schwarz, K.M.; Trimmer, P.A.; Khan, S.M.; Portell, F.R.; et al. Mitochondrial gene therapy augments mitochondrial physiology in a Parkinson’s disease cell model. Hum. Gene Ther. 2009, 20, 897–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oka, S.; Leon, J.; Sakumi, K.; Ide, T.; Kang, D.; LaFerla, F.M.; Nakabeppu, Y. Human mitochondrial transcriptional factor A breaks the mitochondria-mediated vicious cycle in Alzheimer’s disease. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
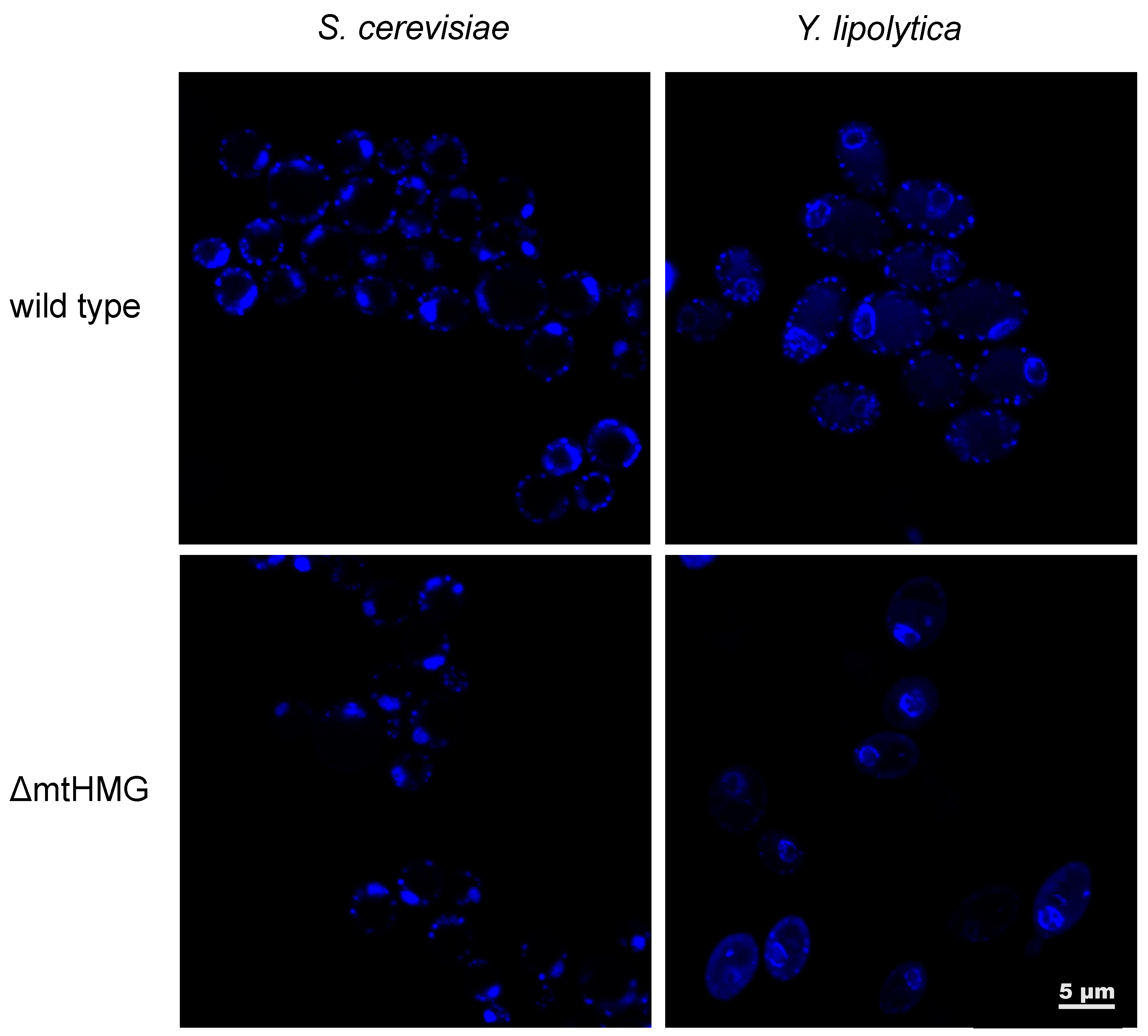{kind=link}
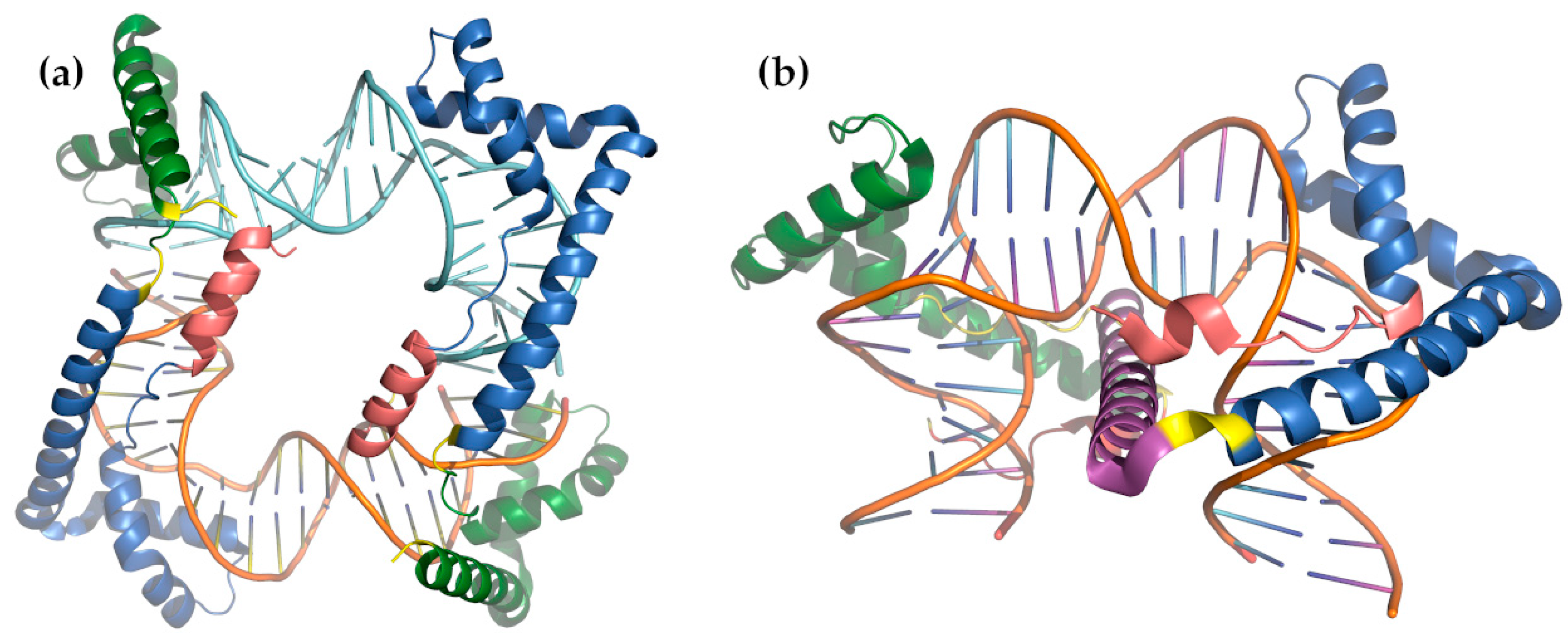{kind=link}
{kind=link}
| Disease | Model | Variant | mRNA | Protein | mtDNA | Ref. |
|---|---|---|---|---|---|---|
| Mitochondrial DNA depletion syndrome 15 | patient | +/+ Pro178Leu | ↑ in primary fibroblasts | ↓ in primary fibroblasts | ↓ copies in liver and skeletal muscle | [266] |
| Extrahepatic cholestasis | cell line rat, patient | gene disruption | ↓ in liver | (↑)↓ in liver | ↓ copies mtDNA deletions in liver | [267,268,269] |
| Cardiomyopathy | mouse | KO in germline | ↓ | ↓ | ↓copy in +/−depletion in +/+ | [130] |
| Cardiomyopathy | mouse | KO in heart | – | ↓ in heart | ↓ transcripts in heart | [135,138,264] |
| Myopathy | mouse | KO in skeletal muscle | – | ↓ in muscle | ↓ copies ↓ transcripts depletion in muscle | [138,139] |
| Infantile mitochondrial myopathy | cell line patient | – | ↑ 1 ↓ 3 | ↓ | depletion 1,3 skeletal muscle | [270] |
| Carotid artery injury | rat | KD | ↑after injury ↓ KD | ↑ after injury ↓ KD | ↑ mass after injury depletion KD | [271] |
| Chronic obstructive pulmonary disease (with squamous cell lung cancer) | cell line patient | hyper-methylation of promoter | ↓ in lungs 3 | ↓ in lungs, skeletal muscle 3 | – | [223,272,273] |
| Gastric cancer | cell line | KD | ↓ | ↓ | ↓ copies | [274] |
| Colorectal cancer with microsatellite instability | cell line patient | Leu149Stop frameshift | – | ↓ | ↓ copies | [275] |
| Colon cancer | mouse | KO +/− | – | ↓ in intestinal tissue 2 | depletion ↓ copy ↓mass | [262] |
| Colon cancer | cell line patient | ↑ miRNA-590-3p | ↑ | ↑ | – | [229] |
| Colon cancer | cell line patient | ↓ miRNA-214 | ↑ | ↑ | – | [276] |
| Epidermoid cancer/ Colon adenocarcinoma | cell line patient | chemo-therapeutics treatment | ↓↑ | ↓↑ | – | [189,277] |
| Bladder cancer | cell line | ↓ miRNA-590-3p | – | ↑ | – | [230] |
| Clear cell renal cell carcinoma | cell line patient | KD 1 | ↓ 1 | ↓ 1 | ↓ copies | [278] |
| Clear cell renal cell carcinoma | cell line | OE SIRT3 | ↑ | ↑ ↓K154 acetylation | ↑biogenesis | [252] |
| Non-small cell lung cancer | cell line patient | KD 1 | ↓ 1 ↑ 3 | ↓ 1 ↑ 3 | ↓ copies 1 ↑ copies 3 | [279] |
| Lung adenocarcinoma/ Breast cancer | cell line | – | ↑in lactic acidosis in long term estradiol treatment | – | ↑ copies ↑ mass in lactic acidosis in long term estradiol treatment | [280,281] |
| Lung adenocarcinoma | cell line | KD, α-irradiation (α-IR) | – | ↑ in α-IR | ↑ copies in α-IR ≈ copies KD in α-IR | [282] |
| Lung adenocarcinoma | cell line patient | – | – | ↓ 1 | ↓ biogenesis ↓ volume | [283] |
| Glioma | cell line patient | ↓ miRNA-23b | ↑ correlation with malignancy | ↑ correlation with malignancy | – | [227,284] |
| Glioma | cell line patient | interaction with KLF16 | ↓ in KLF16 OE | ↓ in KLF16 OE | – | [222] |
| Glioma | cell line | melatonin treatment | ↓ | ↓ | ↓ transcripts ≈ copies | [285] |
| Diffusely infiltrating astrocytoma (a type of glioma) | patient | – | ↑ correlation with malignancy | – | ↓ copies correlation with malignancy | [286] |
| Arsenic-induced Bowen’s disease | cell line patient | arsenic exposure 1,3 RNA interference 1 | ↑ | ↑ | ↑ copies | [287] |
| Melanoma | cell line patient | – | ↓↑ | – | mutations | [288] |
| Prostate cancer | cell line | – | ↑ in arsenic exposure | ↑ in arsenic exposure | mtDNA point mutation in arsenic exposure | [289] |
| Cervical cancer | cell line | miRNA-214 OE | ↓ | ↓ | – | [228] |
| Ovarian cancer | patient | – | – | ↑ in nuclei orrelation with grade | – | [290] |
| Endometrial cancer (the estrogen-related type I) | patient | – | – | ↑ correlation with stage | ↑ copies | [291,292] |
| Breast cancer (estrogen-positive type) | cell line patient | KD 1 | ↑ 3 | ↑ 3 | – | [293,294] |
| Hepatocellular carcinoma | patient | – | – | ↑ | ↓ copies mtDNA deletions | [295] |
| Esophageal squamous cell carcinoma | cell line patient | KD 1 | – | ↓ 1 | ↑ copies 3 ↓ copies 1 | [296,297] |
| Esophageal squamous cell carcinoma | cell line patient | KD 1 | ↓ 1 | ↓ 1 | ↓ copy correlation with malignancy ↓ copies 1 | [298] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vozáriková, V.; Kunová, N.; Bauer, J.A.; Frankovský, J.; Kotrasová, V.; Procházková, K.; Džugasová, V.; Kutejová, E.; Pevala, V.; Nosek, J.; et al. Mitochondrial HMG-Box Containing Proteins: From Biochemical Properties to the Roles in Human Diseases. Biomolecules 2020, 10, 1193. https://doi.org/10.3390/biom10081193
Vozáriková V, Kunová N, Bauer JA, Frankovský J, Kotrasová V, Procházková K, Džugasová V, Kutejová E, Pevala V, Nosek J, et al. Mitochondrial HMG-Box Containing Proteins: From Biochemical Properties to the Roles in Human Diseases. Biomolecules. 2020; 10(8):1193. https://doi.org/10.3390/biom10081193
Chicago/Turabian StyleVozáriková, Veronika, Nina Kunová, Jacob A. Bauer, Ján Frankovský, Veronika Kotrasová, Katarína Procházková, Vladimíra Džugasová, Eva Kutejová, Vladimír Pevala, Jozef Nosek, and et al. 2020. "Mitochondrial HMG-Box Containing Proteins: From Biochemical Properties to the Roles in Human Diseases" Biomolecules 10, no. 8: 1193. https://doi.org/10.3390/biom10081193
APA StyleVozáriková, V., Kunová, N., Bauer, J. A., Frankovský, J., Kotrasová, V., Procházková, K., Džugasová, V., Kutejová, E., Pevala, V., Nosek, J., & Tomáška, Ľ. (2020). Mitochondrial HMG-Box Containing Proteins: From Biochemical Properties to the Roles in Human Diseases. Biomolecules, 10(8), 1193. https://doi.org/10.3390/biom10081193