Biochemical Characterization and Functional Analysis of Heat Stable High Potential Protease of Bacillus amyloliquefaciens Strain HM48 from Soils of Dachigam National Park in Kashmir Himalaya

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
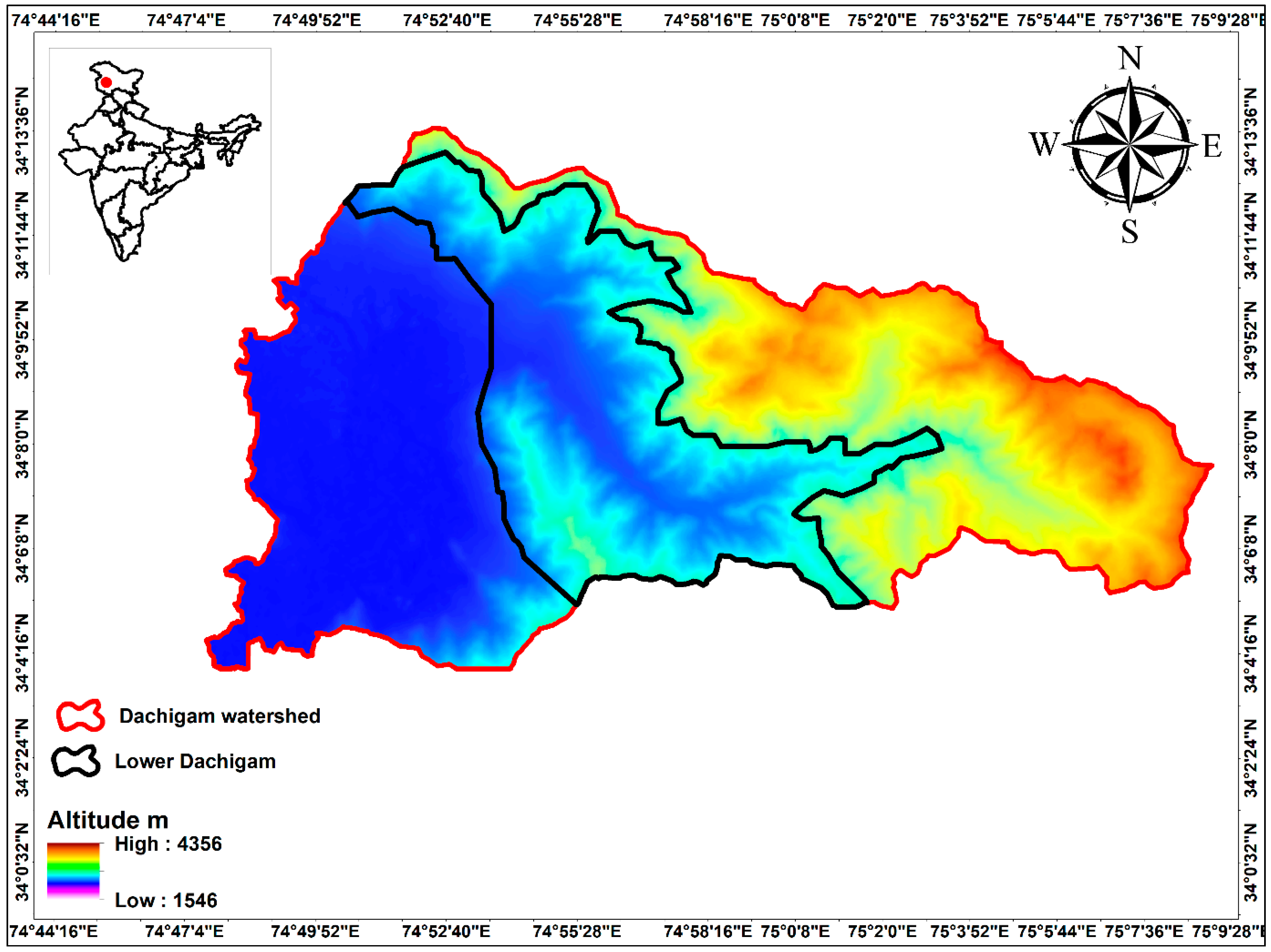
2.1. Sample Collection, Isolation and Preservation of Bacteria
2.2. Preliminary Screening of Isolated Bacterial Strains for the Protease Activity
2.3. Qualitative Screening for Protease Secretion on Different Media
2.4. Identification of Strain, HM48
2.5. Enzyme Production
2.5.1. Media for Protease Production and Culture Conditions
2.5.2. Extraction of the Crude Proteases
2.5.3. Protein Estimation and Protease Activity Assay
2.6. Enzyme Purification
2.6.1. Ammonium Sulfate Precipitation of Crude Proteases
2.6.2. Gel Filtration Column Chromatography and Gel Electrophoresis (SDS–PAGE)
2.7. Enzyme Characterization
2.7.1. Effect of Temperature and pH on Activity/Stability of Protease from Strain, HM48
2.7.2. Influence of Different Substrates, Metal Ions and EDTA on Enzyme Activity of Strain, HM48
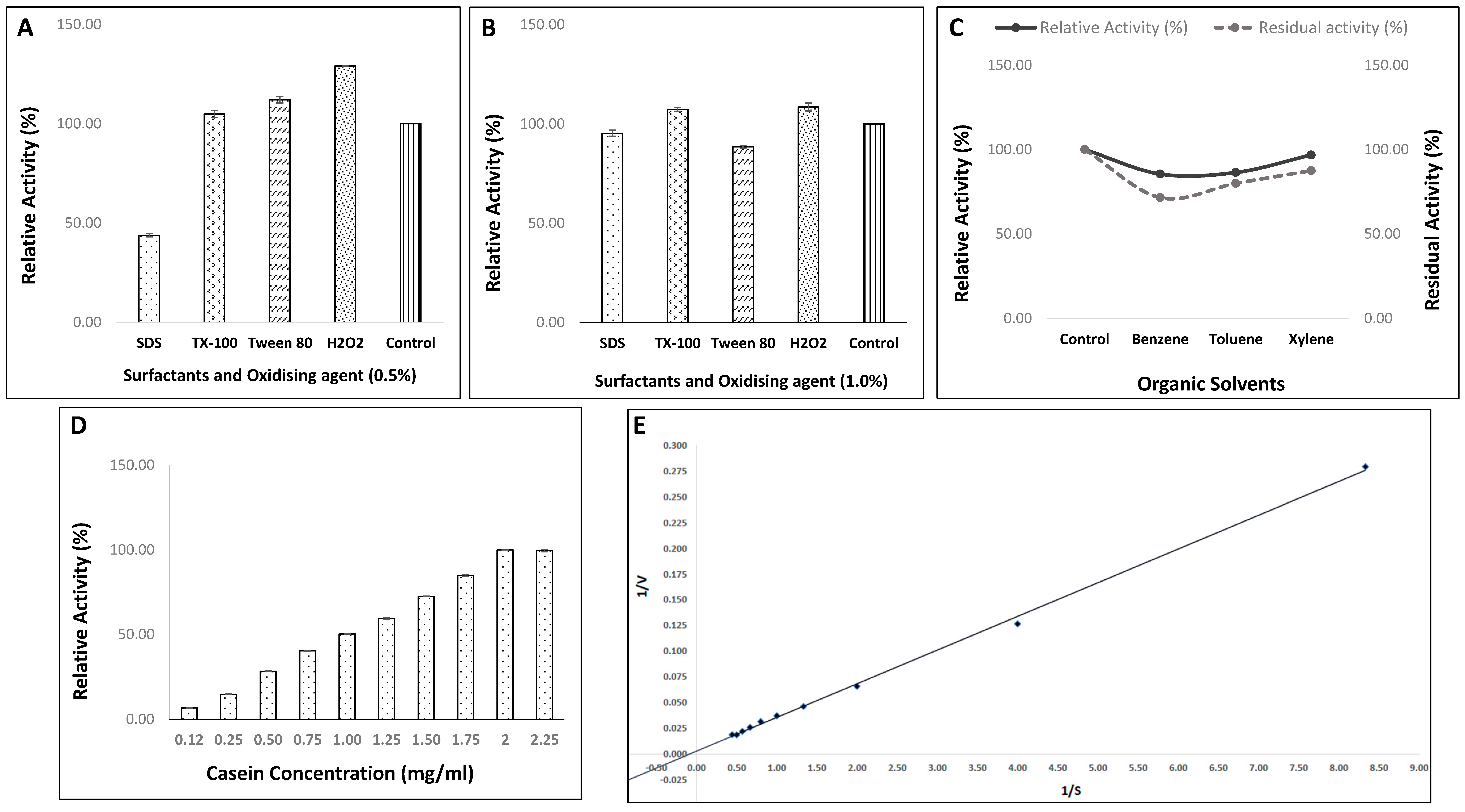
2.7.3. Impact of Various Surfactants and Oxidizing Agent on Enzyme Efficiency of Strain, HM48
2.7.4. Effect of Different Organic Solvents on Activity/Stability of Protease from Strain, HM48
2.7.5. Effect of Varied Casein Concentrations (Enzyme Kinetics) on Enzymatic Activity of Strain, HM48
2.8. Protease Gene Amplification of Strain, HM48
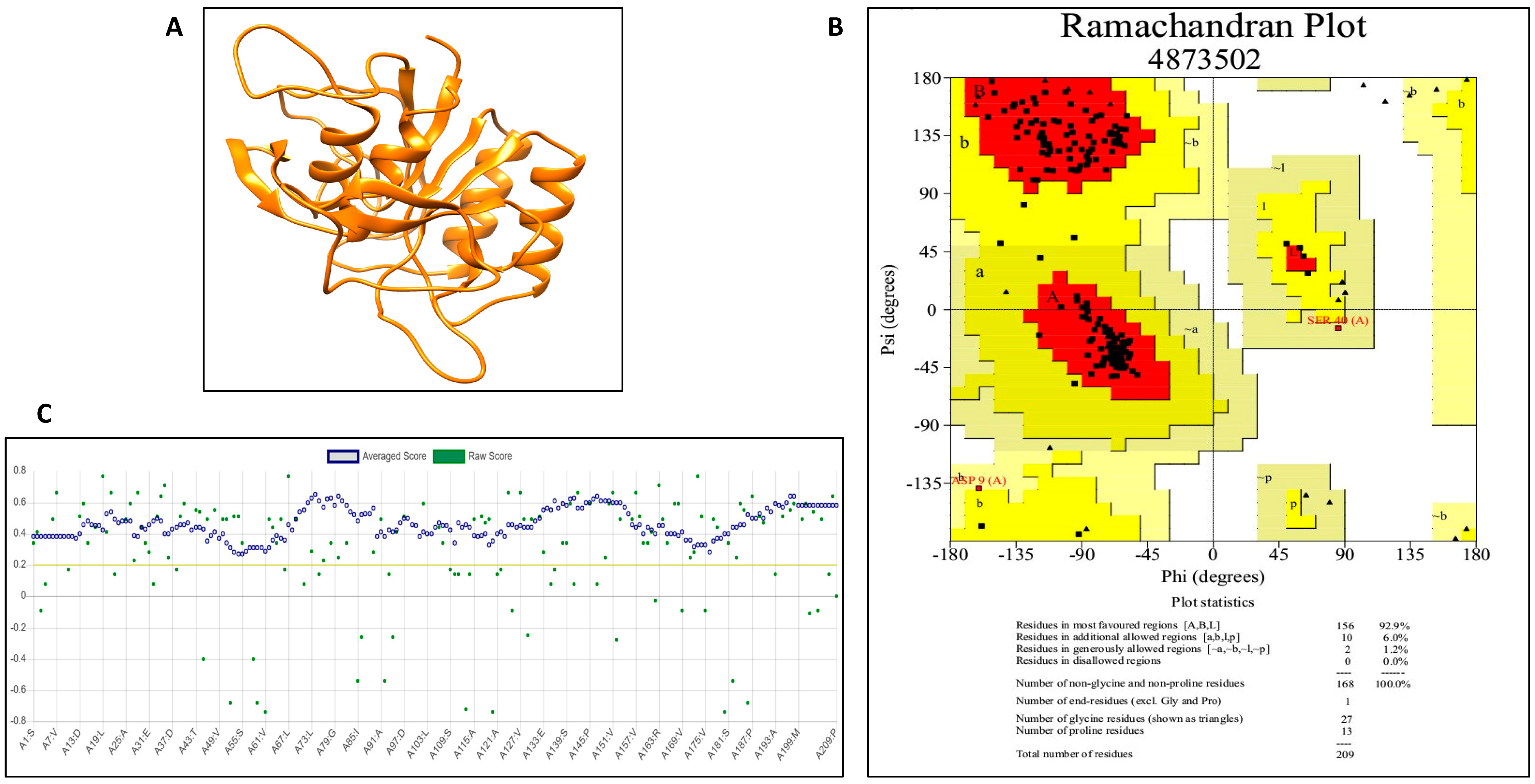
2.8.1. Model Generation
2.8.2. Improvement of Model Quality
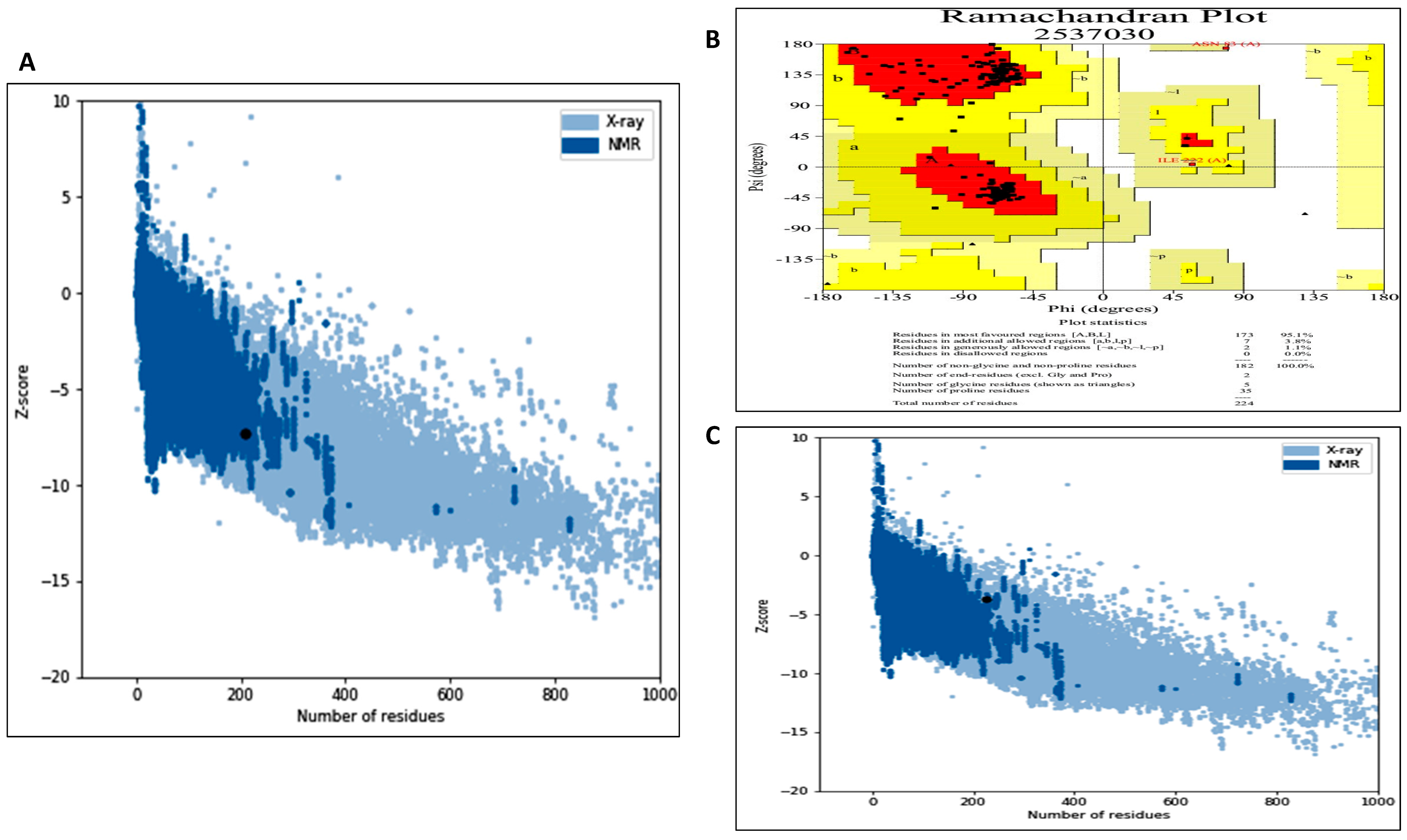
2.8.3. Model Validation through Different Approaches
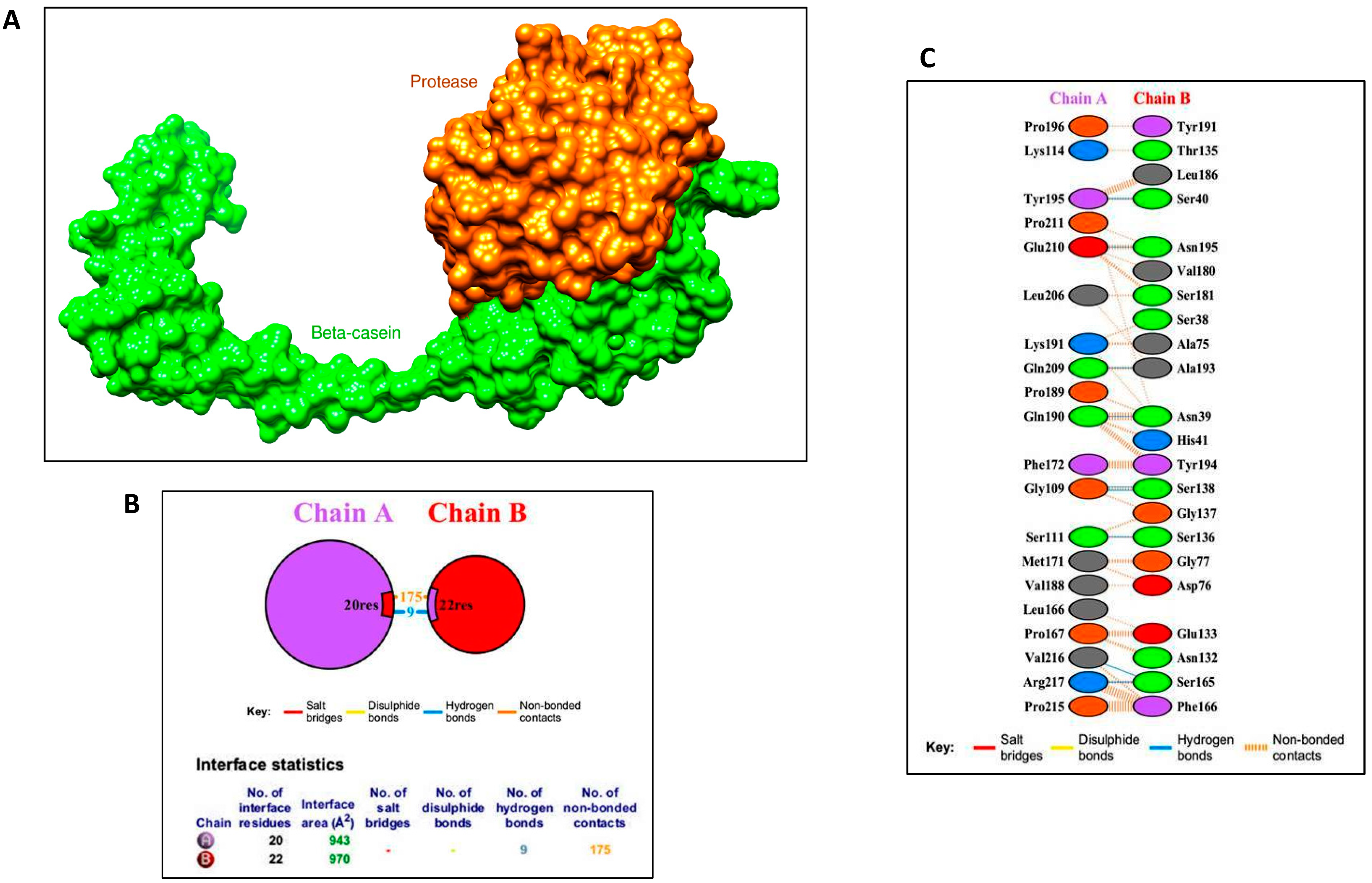
2.8.4. Protein–Protein Docking and Interaction Profile Generation
2.9. Enzyme Application
2.9.1. Evaluation of Enzyme as a Detergent Additive: Enzyme Compatibility with Commercial Detergents
2.9.2. Wash Performance Analysis: Blood and Egg Yolk Stain Removal
- (i)
- 20 mL Surf Excel solution + SC piece
- (ii)
- 20 mL Tide solution + SC piece
- (iii)
- 20 mL Surf Excel solution + 400 μL of enzyme + SC piece
- (iv)
- 20 mL Tide solution + 400 μL of enzyme + SC piece
- (v)
- 20 mL tap water (T.W.) + 400 μL of enzyme + SC piece
- (vi)
- 20 mL tap water (T.W.) + each of blood and egg yolk SC piece as control, respectively
2.9.3. Application of Purified Enzyme in Waste Management: Degradation of Chicken Feather
3. Results
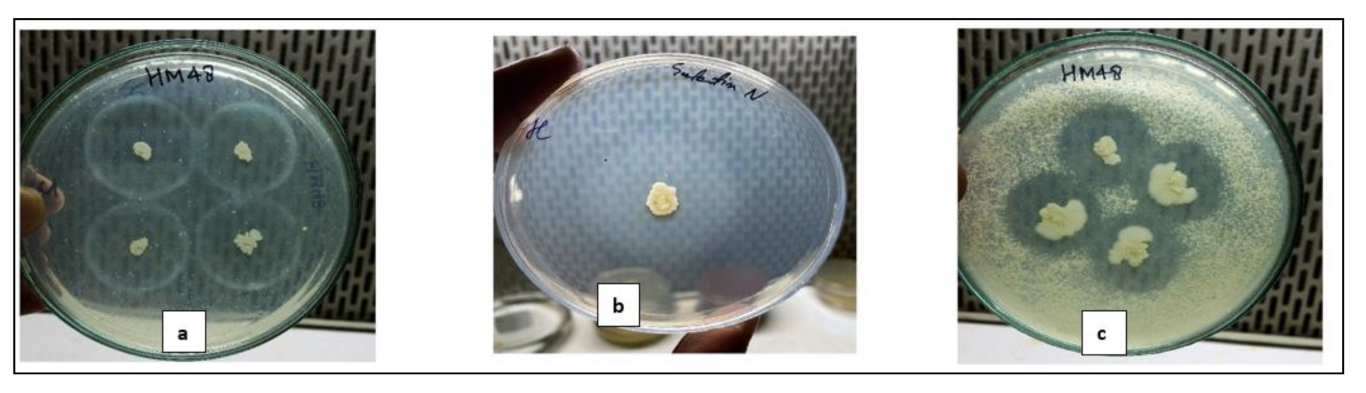
3.1. Isolation and Preliminary Screening of Bacterial Strains for the Protease Activity
3.2. Qualitative Screening and Identification of Strain, HM48
3.2.1. Qualitative Screening on Different Media
3.2.2. Morphological Identification
3.2.3. Biochemical Identification
3.2.4. Molecular Identification
3.3. Purification of Enzyme and Molecular Weight Determination of Protease from Strain, HM48
3.4. Enzyme Characterization
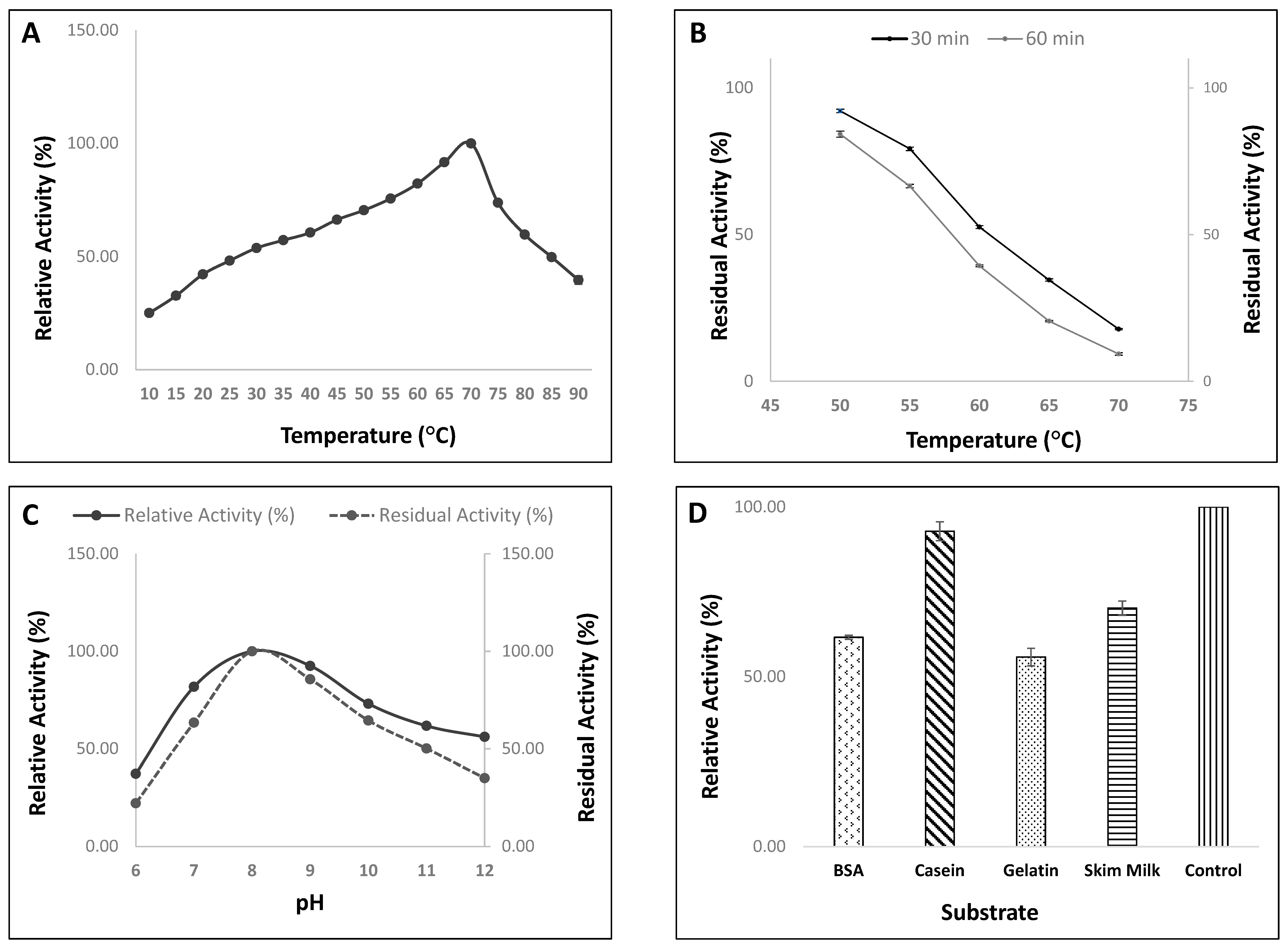
3.4.1. Effect of Temperature on Activity/Stability of Protease from Strain, HM48
3.4.2. Effect of pH on Activity/Stability of Protease Enzyme from Strain, HM48
3.4.3. Effect of Different Substrates on Enzyme Activity of Strain, HM48
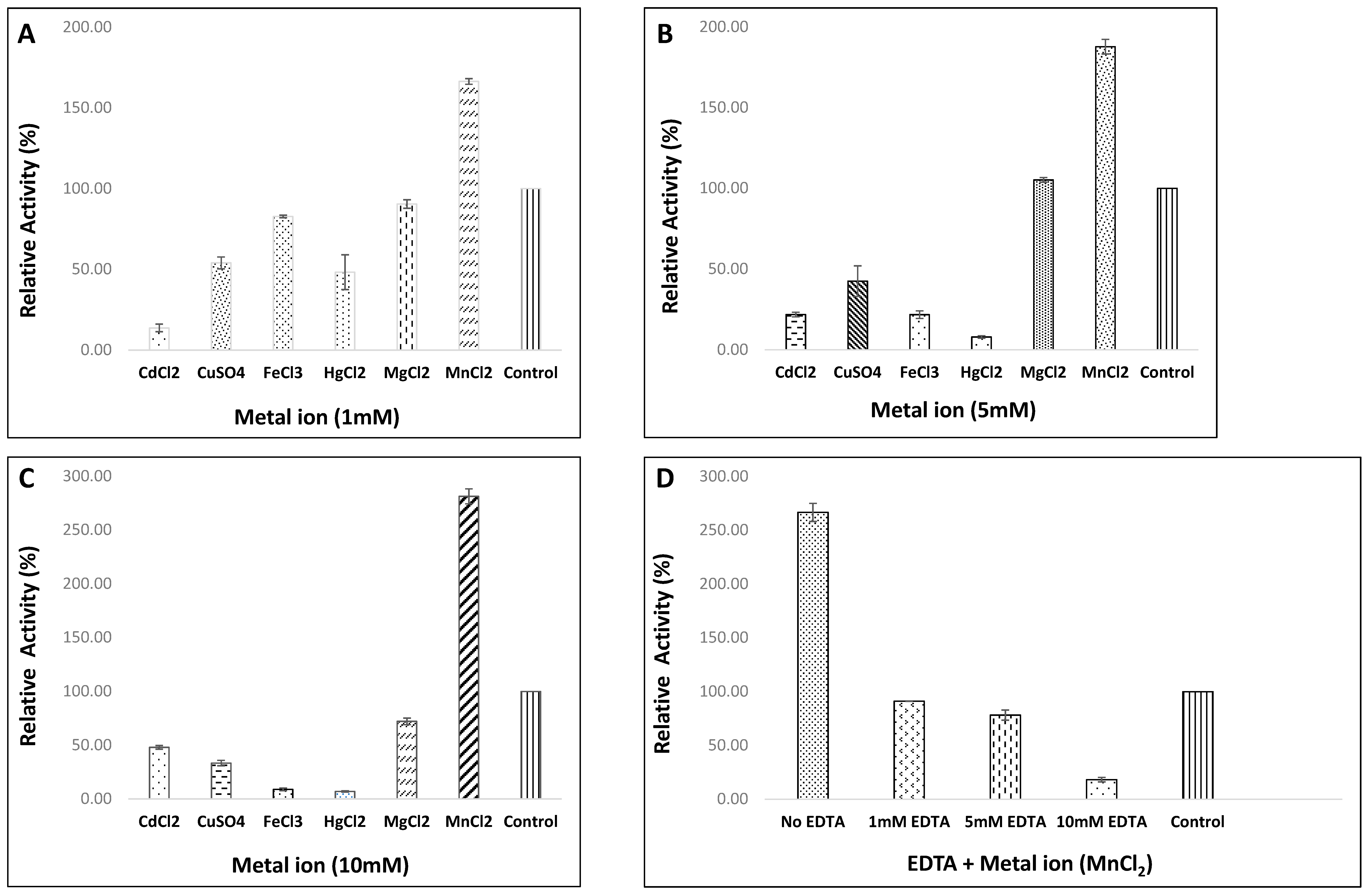
3.4.4. Influence of Various Metal Ions and EDTA on Enzyme Activity of Strain, HM48
3.4.5. Effect of Various Surfactants and Oxidizing Agent on Enzyme Efficiency of Strain, HM48
3.4.6. Effect of Various Organic Solvents on Activity/Stability of Protease from Strain, HM48
3.4.7. Effect of Varied Casein Concentrations (Enzyme Kinetics) on Activity of Protease from Strain, HM48
3.5. Identification of Protease Gene
3.5.1. Model Generation, Improvement and Validation
3.5.2. Protein–Protein Docking
3.6. Application of Enzyme from Strain, HM48
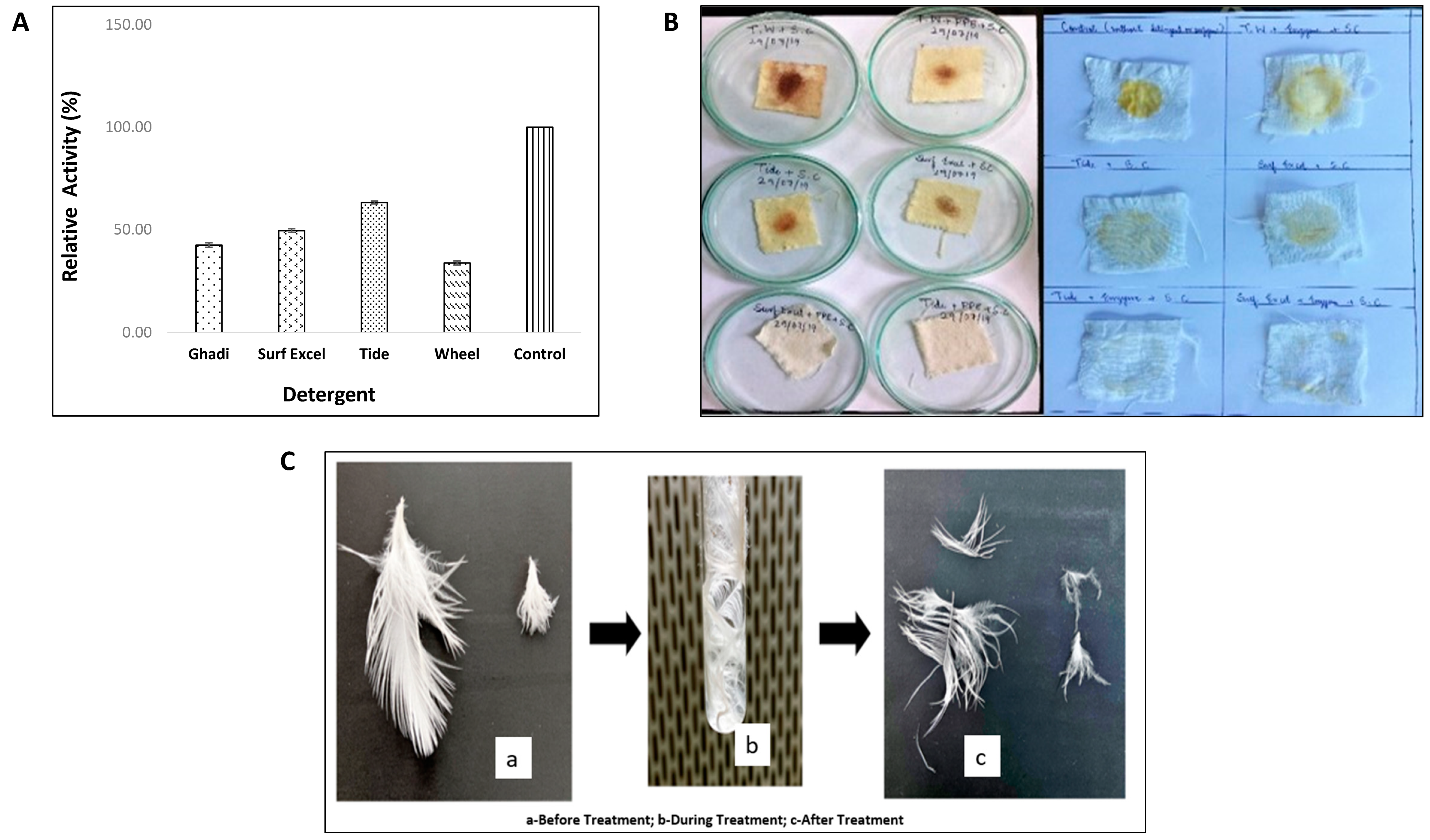
3.6.1. Evaluation of Enzyme as a Detergent Additive: Enzyme-Detergent Compatibility and Wash Performance Analysis
3.6.2. Evaluation of Protease Enzyme for Application in Waste Management: Chicken Feather Degradation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yaqoob, A.; Yunus, M.; Bhat, G.; Singh, D.P. Phytodiversity and Seasonal Variations in the Soil Characteristics of Shrublands of Dachigam National Park, Jammu and Kashmir, India. Clim. Chang. Environ. Sustain. 2015, 3, 137–143. [Google Scholar] [CrossRef]
- Thakur, M.; Sharma, L.K.; Charoo, S.A.; Sathyakumar, S. Conflict Bear Translocation: Investigating Population Genetics and Fate of Bear Translocation in Dachigam National Park, Jammu and Kashmir, India. PLoS ONE 2015, 10, e0132005. [Google Scholar] [CrossRef] [Green Version]
- Mir, Z.R.; Noor, A.; Habib, B.; Gopi, G.V. Seasonal population density and winter survival strategies of endangered Kashmir gray langur (Semnopithecus ajax) in Dachigam National Park, Kashmir, India. SpringerPlus 2015, 4, 562. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, K.; Qureshi, Q.; Agoramoorthy, G.; Nigam, P. Habitat use patterns and food habits of the Kashmir red deer or Hangul (Cervus elaphus hanglu) in Dachigam National Park, Kashmir, India. Ethol. Ecol. Evol. 2015, 28, 85–101. [Google Scholar] [CrossRef]
- Ahmad, N.; Johri, S.; Abdin, M.Z.; Qazi, G.N. Molecular characterization of bacterial population in the forest soil of Kashmir, India. World J. Microbiol. Biotechnol. 2008, 25, 107–113. [Google Scholar] [CrossRef]
- Furhan, J.; Awasthi, P.; Sharma, S. Biochemical characterization and homology modelling of cold-active alkophilic protease from Northwestern Himalayas and its application in detergent industry. Biocatal. Agric. Biotechnol. 2019, 17, 726–735. [Google Scholar] [CrossRef]
- Yadav, A.N.; Sachan, S.G.; Verma, P.; Kaushik, R.; Saxena, A.K. Cold active hydrolytic enzymes production by psychrotrophic Bacilli isolated from three sub-glacial lakes of NW Indian Himalayas. J. Basic Microbiol. 2016, 56, 294–307. [Google Scholar] [CrossRef]
- Ahmad, W.; Tayyab, M.; Aftab, M.N.; Hashmi, A.S.; Ahmad, M.D.; Firyal, S.; Wasim, M.; Awan, A.R. Optimization of Conditions for the Higher Level Production of Protease: Characterization of Protease from Geobacillus SBS-4S. Waste Biomass Valoriz. 2020, 11, 6613–6623. [Google Scholar] [CrossRef]
- Jisha, V.N.; Smitha, R.B.; Pradeep, S.; Sreedevi, S.; Unni, K.N.; Sajith, S.; Priji, P.; Josh, M.S.; Benjamin, S. Versatility of microbial proteases. Adv. Enzym. Res. 2013, 1, 39–51. [Google Scholar] [CrossRef]
- Wang, J.; Xu, A.; Wan, Y.; Li, Q. Purification and Characterization of a New Metallo-Neutral Protease for Beer Brewing from Bacillus amyloliquefaciens SYB-001. Appl. Biochem. Biotechnol. 2013, 170, 2021–2033. [Google Scholar] [CrossRef]
- Si, J.-B.; Jang, E.-J.; Charalampopoulos, D.; Wee, Y.-J. Purification and Characterization of Microbial Protease Produced Extracellularly from Bacillus subtilis FBL-1. Biotechnol. Bioprocess Eng. 2018, 23, 176–182. [Google Scholar] [CrossRef] [Green Version]
- Jadhav, H.P.; Sonawane, M.S.; Khairnar, M.H.; Sayyed, R.Z. Production of alkaline protease by rhizospheric Bacillus cereus HP_RZ17 and Paenibacillus xylanilyticus HP_RZ19. Environ. Sustain. 2020, 3, 5–13. [Google Scholar] [CrossRef]
- Griffin, H.L.; Greene, R.V.; Cotta, M. Isolation and characterization of an alkaline protease from the marine shipworm bacterium. Curr. Microbiol. 1992, 24, 111–117. [Google Scholar] [CrossRef]
- Iyer, P.V.; Ananthanarayan, L. Enzyme stability and stabilization—Aqueous and non-aqueous environment. Process. Biochem. 2008, 43, 1019–1032. [Google Scholar] [CrossRef]
- Qureshi, A.S.; Bhutto, M.A.; Khushk, I.; Dahot, M.U. Optimization of cultural conditions for protease production by Bacillus subtilis EFRL 01. Afr. J. Biotechnol. 2011, 10, 5173–5181. [Google Scholar] [CrossRef]
- Sumantha, A.; Larroche, C.; Pandey, A. Microbiology and industrial biotechnology of food-grade proteases: A perspective. Food Technol. Biotechnol. 2006, 44, 211. [Google Scholar]
- Gupta, R.; Beg, Q.; Lorenz, P. Bacterial alkaline proteases: Molecular approaches and industrial applications. Appl. Microbiol. Biotechnol. 2002, 59, 15–32. [Google Scholar] [CrossRef]
- Annamalai, N.; Rajeswari, M.V.; Balasubramanian, T. Extraction, purification and application of thermostable and halostable alkaline protease from Bacillus alveayuensis CAS 5 using marine wastes. Food Bioprod. Process. 2014, 92, 335–342. [Google Scholar] [CrossRef]
- Enshasy, H.E.; Enein, A.; Helmy, S.; Azaly, Y.E. Optimization of the industrial production of alkaline protease by Bacillus licheniformis in different production scales. Aust. J. Basic Appl. Sci. 2008, 2, 583–593. [Google Scholar]
- Wilson, P. Production and characterisation of protease enzyme produced by a novel moderate thermophilic bacterium (EP1001) isolated from an alkaline hot spring, Zimbabwe. Afr. J. Microbiol. Res. 2012, 6, 5542–5551. [Google Scholar] [CrossRef]
- Manachini, P.L.; Fortina, M.G. Production in sea-water of thermostable alkaline proteases by a halotolerant strain of Bacillus licheniformis. Biotechnol. Lett. 1998, 20, 565–568. [Google Scholar] [CrossRef]
- Denizci, A.A.; Kazan, D.; Abeln, E.; Erarslan, A. Newly isolated Bacillus clausii GMBAE 42: An alkaline protease producer capable to grow under higly alkaline conditions. J. Appl. Microbiol. 2004, 96, 320–327. [Google Scholar] [CrossRef] [Green Version]
- Haddar, A.; Agrebi, R.; Bougatef, A.; Hmidet, N.; Sellami-Kamoun, A.; Nasri, M. Two detergent stable alkaline serine-proteases from Bacillus mojavensis A21: Purification, characterization and potential application as a laundry detergent additive. Bioresour. Technol. 2009, 100, 3366–3373. [Google Scholar] [CrossRef]
- Chen, T.-L.; Chou, Y.-J.; Chen, W.-M.; Arun, B.; Young, C.-C. Tepidimonas taiwanensis sp. nov., a novel alkaline-protease-producing bacterium isolated from a hot spring. Extremophiles 2005, 10, 35–40. [Google Scholar] [CrossRef]
- El Hidri, D.; Guesmi, A.; Najjari, A.; Cherif, H.; Ettoumi, B.; Hamdi, C.; Boudabous, A.; Cherif, A. Cultivation-Dependant Assessment, Diversity, and Ecology of Haloalkaliphilic Bacteria in Arid Saline Systems of Southern Tunisia. BioMed Res. Int. 2013, 2013, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Nassar, F.R.; Abdelhafez, A.A.; El-Tayeb, T.S.; Abu-Hussein, S.H. Purification, Characterization and Applications of Proteases Produced by Bacillus amyloliquefaciens 35s Isolated from Soil of the Nile Delta of Egypt. Br. Microbiol. Res. J. 2015, 6, 286–302. [Google Scholar] [CrossRef]
- Cho, S.-J.; Oh, S.-H.; Pridmore, R.D.; Juillerat, M.A.; Lee, C.-H. Purification and Characterization of Proteases fromBacillus amyloliquefaciensIsolated from Traditional Soybean Fermentation Starter. J. Agric. Food Chem. 2003, 51, 7664–7670. [Google Scholar] [CrossRef]
- Longo, M.A.; Novella, I.S.; Garcia, L.A.; Diaz, M. Comparison of Bacillus subtilis and Serratia marcescens as protease producers under different operating conditions. J. Biosci. Bioeng. 1999, 88, 35–40. [Google Scholar] [CrossRef]
- Cui, H.; Yang, M.; Wang, L.; Xian, C.J. Identification of a New Marine Bacterial Strain SD8 and Optimization of Its Culture Conditions for Producing Alkaline Protease. PLoS ONE 2015, 10, e0146067. [Google Scholar] [CrossRef]
- Jammu and Kashmir Forest Department, JKFD. Handbook of Forest Statistics; Jammu and Kashmir Forest Department (JKFD): Srinagar, Jammu and Kashmir, India, 2011.
- Anderson, J.M.; Ingram, J.S.I. Tropical Soil Biology and Fertility: A Handbook of Methods; CAB International: Wallingford, WA, USA, 1993. [Google Scholar]
- Kumar, R.; Acharya, C.; Joshi, S.R. Isolation and analyses of uranium tolerant Serratia marcescens strains and their utilization for aerobic uranium U(VI) bioadsorption. J. Microbiol. 2011, 49, 568–574. [Google Scholar] [CrossRef]
- Baghel, V.S.; Tripathi, R.D.; Ramteke, P.W.; Gopal, K.; Dwivedi, S.; Jain, R.K.; Rai, U.N.; Singh, S.N. Psychrotrophic proteolytic bacteria from cold environment of Gangotri glacier, Western Himalaya, India. Enzym. Microb. Technol. 2005, 36, 654–659. [Google Scholar] [CrossRef]
- Baweja, M.; Tiwari, R.; Singh, P.K.; Nain, L.; Shukla, P. An Alkaline Protease from Bacillus pumilus MP 27: Functional Analysis of Its Binding Model toward Its Applications As Detergent Additive. Front. Microbiol. 2016, 7, 1195. [Google Scholar] [CrossRef] [Green Version]
- Bergey, D.H.; Holt, J.G. Bergey’s Manual of Determinative Bacteriology; Williams & Wilkins: Baltimore, MD, USA, 1994. [Google Scholar]
- Pammi, N.; Chaitanya, K.; Mahmood, S.K. Serratia marcescens OU50T sp. nov., a cellulose and pha producing bacterium isolated from polluted water. Int. J. Environ. Biol. 2015, 5, 32–36. [Google Scholar]
- Sivanandhini, T.; Subbaiya, R.; Gopinath, M.; Angrasan, J.K.V.; Kabilan, T.; Selvam, M.M. An investigation on morphological characterization of Actinomycetes isolated from marine sediments. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 1234–1243. [Google Scholar]
- Saggu, S.K.; Mishra, P.C. Characterization of thermostable alkaline proteases from Bacillus infantis SKS1 isolated from garden soil. PLoS ONE 2017, 12, e0188724. [Google Scholar] [CrossRef] [Green Version]
- Gürtler, V.; Stanisich, V.A. New approaches to typing and identification of bacteria using the 16S-23S rDNA spacer region. Microbiology 1996, 142, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.I.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Cupp-Enyard, C. Sigma’s Non-specific Protease Activity Assay—Casein as a Substrate. J. Vis. Exp. 2008, 17, e899. [Google Scholar] [CrossRef]
- Green, A.A.; Hughes, W.L. [10] Protein fractionation on the basis of solubility in aqueous solutions of salts and organic solvents. Methods Enzymol. 1955, 1, 67–90. [Google Scholar] [CrossRef]
- Sadia, A.A.; Shelly, D.; Mohan, P. Purification and characterization of a novel protease from Bacillus strain Sal1. Afr. J. Biotechnol. 2009, 8, 3603–3609. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Lineweaver, H.; Burk, D. The Determination of Enzyme Dissociation Constants. J. Am. Chem. Soc. 1934, 56, 658–666. [Google Scholar] [CrossRef]
- Ko, J.; Park, H.; Seok, C. GalaxyTBM: Template-based modeling by building a reliable core and refining unreliable local regions. BMC Bioinform. 2012, 13, 198. [Google Scholar] [CrossRef] [Green Version]
- Heo, L.; Park, H.; Seok, C. GalaxyRefine: Protein structure refinement driven by side-chain repacking. Nucleic Acids Res. 2013, 41, W384–W388. [Google Scholar] [CrossRef] [Green Version]
- Farooq, Z.; Abdullah, E.; Banday, S.; Ganai, S.A.; Rashid, R.; Mushtaq, A.; Rashid, S.; Altaf, M. Vigilin protein Vgl1 is required for heterochromatin-mediated gene silencing in Schizosaccharomyces pombe. J. Biol. Chem. 2019, 294, 18029–18040. [Google Scholar] [CrossRef]
- Lüthy, R.; Bowie, J.U.; Eisenberg, D. Assessment of protein models with three-dimensional profiles. Nat. Cell Biol. 1992, 356, 83–85. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35 (Suppl. 2), W407–W410. [Google Scholar] [CrossRef] [Green Version]
- Hou, T.; Qiao, X.; Zhang, W.; Xu, X. Empirical Aqueous Solvation Models Based on Accessible Surface Areas with Implicit Electrostatics. J. Phys. Chem. B 2002, 106, 11295–11304. [Google Scholar] [CrossRef]
- Zacharias, M. Protein-protein docking with a reduced protein model accounting for side-chain flexibility. Protein Sci. 2003, 12, 1271–1282. [Google Scholar] [CrossRef]
- Feng, T.; Chen, F.; Kang, Y.; Sun, H.; Liu, H.; Li, D.; Zhu, F.; Hou, T.-J. HawkRank: A new scoring function for protein–protein docking based on weighted energy terms. J. Cheminform. 2017, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Weng, G.; Wang, E.; Wang, Z.; Liu, H.; Zhu, F.; Li, D.; Hou, T.-J. HawkDock: A web server to predict and analyze the protein–protein complex based on computational docking and MM/GBSA. Nucleic Acids Res. 2019, 47, W322–W330. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Jabłońska, J.; Pravda, L.; Vařeková, R.S.; Thornton, J.M. PDBsum: Structural summaries of PDB entries. Protein Sci. 2018, 27, 129–134. [Google Scholar] [CrossRef]
- Bhunia, B.; Basak, B.; Mandal, T.; Bhattacharya, P.; Dey, A. Effect of pH and temperature on stability and kinetics of novel extracellular serine alkaline protease (70 kDa). Int. J. Biol. Macromol. 2013, 54, 1–8. [Google Scholar] [CrossRef]
- Jedynak, L.; Kowalska, J.; Harasimowicz, J.; Golimowskim, J. Speciation analysis of arsenic in terrestrial plants from arsenic contaminated area. Sci. Total Environ. 2009, 407, 945–952. [Google Scholar] [CrossRef]
- Singh, S.K.; Singh, S.K.; Tripathi, V.R.; Khare, S.K.; Garg, S.K. A novel psychrotrophic, solvent tolerant Pseudomonas putida SKG-1 and solvent stability of its psychro-thermoalkalistable protease. Process. Biochem. 2011, 46, 1430–1435. [Google Scholar] [CrossRef]
- Sánchez-Ramos, I.; Hernandez, C.A.; Castanera, P.; Ortego, F. Proteolytic activities in body and faecal extracts of the storage mite, Acarus farris. Med. Vet. Entomol. 2004, 18, 378–386. [Google Scholar] [CrossRef]
- Milton, D.K.; Chawla, R.K. Cotton dust contains proteolytic and elastolytic enzymes not inhibited by alpha-1-proteinase inhibitor. Am. J. Ind. Med. 1986, 9, 247–260. [Google Scholar] [CrossRef]
- Rudenskaya, G.E.; Pupov, D.V. Cysteine proteinases of microorganisms and viruses. Biochemistry (Moscow) 2011, 73, 1–13. [Google Scholar] [CrossRef]
- Shankar, S.; Rao, M.; Laxman, R.S. Purification and characterization of an alkaline protease by a new strain of Beauveria sp. Process. Biochem. 2011, 46, 579–585. [Google Scholar] [CrossRef]
- Willey, J.M.; Sherwood, L.; Woolverton, C.J. Prescott’s Microbiology; McGraw-Hill: New York, NY, USA, 2011; Volume 7. [Google Scholar]
- Thebti, W.; Riahi, Y.; Belhadj, O. Purification and Characterization of a New Thermostable, Haloalkaline, Solvent Stable, and Detergent Compatible Serine Protease fromGeobacillus toebiiStrain LBT 77. BioMed Res. Int. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kamran, A.; Rehman, H.U.; Qader, S.A.U.; Baloch, A.H.; Kamal, M. Purification and characterization of thiol dependent, oxidation-stable serine alkaline protease from thermophilic Bacillus sp. J. Genet. Eng. Biotechnol. 2015, 13, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Hakim, A.; Bhuiyan, F.R.; Iqbal, A.; Emon, T.H.; Ahmed, J.; Azad, A.K. Production and partial characterization of dehairing alkaline protease from Bacillus subtilis AKAL7 and Exiguobacterium indicum AKAL11 by using organic municipal solid wastes. Heliyon 2018, 4, e00646. [Google Scholar] [CrossRef]
- Ramadan, M.F.; Mahmoud, M.G.; El Shebwy, K.; Abdel-Aziz, M.S. Purification and characterization of two thermostable protease fractions from Bacillus megaterium. J. Genet. Eng. Biotechnol. 2013, 11, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Thakur, N.; Kumar, A.; Sharma, A.; Bhalla, T.C.; Kumar, D. Purification and characterization of alkaline, thermostable and organic solvent stable protease from a mutant of Bacillus sp. Biocatal. Agric. Biotechnol. 2018, 16, 217–224. [Google Scholar] [CrossRef]
- Sai-Ut, S.; Benjakul, S.; Sumpavapol, P.; Kishimura, H. Purification and Characterization of Extracellular Gelatinolytic Protease fromBacillus Amyloliquefaciens H11. J. Food Biochem. 2015, 39, 119–128. [Google Scholar] [CrossRef]
- Lakshmi, B.K.; Kumar, D.M.; Hemalatha, K.P. Purification and characterization of alkaline protease with novel properties from Bacillus cereus strain S8. J. Genet. Eng. Biotechnol. 2018, 16, 295–304. [Google Scholar] [CrossRef]
- Maruthiah, T.; Esakkiraj, P.; Prabakaran, G.; Palavesam, A.; Immanuel, G. Purification and characterization of moderately halophilic alkaline serine protease from marine Bacillus subtilis AP-MSU 6. Biocatal. Agric. Biotechnol. 2013, 2, 116–119. [Google Scholar] [CrossRef]
- Guleria, S.; Walia, A.; Chauhan, A.; Shirkot, C.K. Purification and characterization of detergent stable alkaline protease fromBacillus amyloliquefaciensSP1 isolated from apple rhizosphere. J. Basic Microbiol. 2015, 56, 138–152. [Google Scholar] [CrossRef]
- Klomklao, S.; Benjakul, S.; Kishimura, H.; Chaijan, M. Extraction, purification and properties of trypsin inhibitor from Thai mung bean (Vigna radiata (L.) R. Wilczek). Food Chem. 2011, 129, 1348–1354. [Google Scholar] [CrossRef]
- Sinha, R.; Khare, S.K. Thermostable Proteases. In Thermophilic Microbes in Environmental and Industrial Biotechnology; Satyanarayana, T., Littlechild, J., Kawarabayasi, Y., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 859–880. [Google Scholar]
- Kumar, S.; Nussinov, R. Salt bridge stability in monomeric proteins 1 1Edited by J. M. Thornton. J. Mol. Biol. 1999, 293, 1241–1255. [Google Scholar] [CrossRef]
- Ozawa, T.; Hakamada, Y.; Hatada, Y.; Kobayashi, T.; Shirai, T.; Ito, S. Thermostabilization by replacement of specific residues with lysine in a Bacillus alkaline cellulase: Building a structural model and implications of newly formed double intrahelical salt bridges. Protein Eng. 2001, 14, 501–504. [Google Scholar] [CrossRef] [Green Version]
- Szilágyi, A.; Závodszky, P. Structural differences between mesophilic, moderately thermophilic and extremely thermophilic protein subunits: Results of a comprehensive survey. Structure 2000, 8, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Dangi, A.K.; Shukla, P.; Baishya, D.; Khare, S.K. Thermozymes: Adaptive strategies and tools for their biotechnological applications. Bioresour. Technol. 2019, 278, 372–382. [Google Scholar] [CrossRef]
- Farhadian, S.; Asoodeh, A.; Lagzian, M. Purification, biochemical characterization and structural modeling of a potential htrA-like serine protease from Bacillus subtilis DR8806. J. Mol. Catal. B Enzym. 2015, 115, 51–58. [Google Scholar] [CrossRef]
- Mothe, T.; Reddy, S.V. Production, purification and characterization of a thermotolerant alkaline serine protease from a novel species Bacillus caseinilyticus. 3 Biotech 2016, 6, 53. [Google Scholar] [CrossRef] [Green Version]
- Theron, L.W.; Divol, B. Microbial aspartic proteases: Current and potential applications in industry. Appl. Microbiol. Biotechnol. 2014, 98, 8853–8868. [Google Scholar] [CrossRef]
- Olajuyigbe, F.M.; Falade, A.M. Purification and partial characterization of serine alkaline metalloprotease from Bacillus brevis MWB-01. Bioresour. Bioprocess. 2014, 1, 8. [Google Scholar] [CrossRef]
- Gulmus, E.O.; Görmez, A. Characterization and biotechnological application of protease from thermophilic Thermomonas haemolytica. Arch. Microbiol. 2019, 202, 153–159. [Google Scholar] [CrossRef]
- Durham, D.R.; Stewart, D.B.; Stellwag, E.J. Novel alkaline- and heat-stable serine proteases from alkalophilic Bacillus sp. strain GX6638. J. Bacteriol. 1987, 169, 2762–2768. [Google Scholar] [CrossRef] [Green Version]
- Rick, W. Methoden der Enzymatischen Analyse, 3rd ed.; Bergmeyer, H.U., Ed.; Verlag Chemic: Weinheim, Germany, 1974; pp. 1046–1056. [Google Scholar]
- Reimerdes, E.H.; Klostermeyer, H. substrates. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1976; Volume 45, pp. 26–28. [Google Scholar]
- Thomas, T.D.; Pritchard, G.G. Proteolytic enzymes of dairy starter cultures. FEMS Microbiol. Lett. 1987, 46, 245–268. [Google Scholar] [CrossRef]
- Kumar, C.; Takagi, H. Microbial alkaline proteases. Biotechnol. Adv. 1999, 17, 561–594. [Google Scholar] [CrossRef]
- Rao, C.S.; Sathish, T.; Ravichandra, P.; Prakasham, R. Characterization of thermo- and detergent stable serine protease from isolated Bacillus circulans and evaluation of eco-friendly applications. Process. Biochem. 2009, 44, 262–268. [Google Scholar] [CrossRef]
- Bernhoft, R.A. Mercury Toxicity and Treatment: A Review of the Literature. J. Environ. Public Heal. 2011, 2012, 1–10. [Google Scholar] [CrossRef]
- Ibrahim, A.S.S.; Elbadawi, Y.B.; El-Tayeb, M.A.; Al-Maary, K.S.; Maany, D.A.F.; Ibrahim, S.S.S.; Elagib, A.A. Alkaline serine protease from the new halotolerant alkaliphilic Salipaludibacillus agaradhaerens strain AK-R: Purification and properties. 3 Biotech 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Hammami, A.; Hamdi, M.; Abdelhedi, O.; Jridi, M.; Nasri, M.; Bayoudh, A. Surfactant- and oxidant-stable alkaline proteases from Bacillus invictae: Characterization and potential applications in chitin extraction and as a detergent additive. Int. J. Biol. Macromol. 2017, 96, 272–281. [Google Scholar] [CrossRef]
- El-Eskafy, H.; Abdel-Hamid, M.; Abbas, R.; Hamza, H.; Zanaty, A.E. Production of Thermostable Protease from “Bacillus amyloliquefaciens”: An Egyptian Marine Isolate. J. Agric. Chem. Biotechnol. 2016, 7, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Anwar, A.; Saleemuddin, M. Alkaline protease from Spilosoma obliqua: Potential applications in bio-formulations. Biotechnol. Appl. Biochem. 2000, 31, 85–89. [Google Scholar] [CrossRef]
- Takami, H.; Kobayashi, T.; Aono, R.; Horikoshi, K. Molecular cloning, nucleotide sequence and expression of the structural gene for a thermostable alkaline protease from Bacillus sp. no. AH-101. Appl. Microbiol. Biotechnol. 1992, 38, 101–108. [Google Scholar] [CrossRef]
- Ghorbel-Frikha, B.; Sellami-Kamoun, A.; Fakhfakh, N.; Haddar, A.; Manni, L.; Nasri, M. Production and purification of a calcium-dependent protease from Bacillus cereus BG1. J. Ind. Microbiol. Biotechnol. 2005, 32, 186–194. [Google Scholar] [CrossRef]
- Horn, S.J.; Sørlie, M.; Vaaje-Kolstad, G.; Norberg, A.L.; Synstad, B.; Vårum, K.M.; Eijsink, V.G.H. Comparative studies of chitinases A, B and C fromSerratia marcescens. Biocatal. Biotransform. 2006, 24, 39–53. [Google Scholar] [CrossRef]
- Emran, M.A.; Ismail, S.A.; Hashem, A.M. Production of detergent stable thermophilic alkaline protease by Bacillus licheniformis ALW1. Biocatal. Agric. Biotechnol. 2020, 26, 101631. [Google Scholar] [CrossRef]
- Gaonkar, S.K.; Furtado, I.J. Characterization of Extracellular Protease from the Haloarcheon Halococcus sp. Strain GUGFAWS-3 (MF425611). Curr. Microbiol. 2020, 77, 1024–1034. [Google Scholar] [CrossRef]
- Hooft, R.W.; Sander, C.; Vriend, G. Objectively judging the quality of a protein structure from a Ramachandran plot. Bioinformatics 1997, 13, 425–430. [Google Scholar] [CrossRef] [Green Version]
- Jain, C.K.; Gupta, M.; Prasad, Y.; Wadhwa, G.; Sharma, S.K. Homology modeling and protein engineering of alkane monooxygenase in Burkholderia thailandensis MSMB121: In silico insights. J. Mol. Model. 2014, 20, 1–11. [Google Scholar] [CrossRef]
- Prajapat, R.; Marwal, A.; Gaur, R.K. Recognition of Errors in the Refinement and Validation of Three-Dimensional Structures of AC1 Proteins of Begomovirus Strains by Using ProSA-Web. J. Viruses 2014, 2014, 1–6. [Google Scholar] [CrossRef]
- Fakhfakh, N.; Haddar, A.; Hmidet, N.; Frikha, F.; Nasri, M. Application of statistical experimental design for optimization of keratinases production by Bacillus pumilus A1 grown on chicken feather and some biochemical properties. Process. Biochem. 2010, 45, 617–626. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteinaceous Substrate | Mean Diameter of Colony, d (in mm) | Mean Diameter of Hydrolytic Zone, D (in mm) | D/d (in mm) |
|---|---|---|---|
| Casein | 3 | 28 | 9.33 |
| Gelatin | 10 | 33 | 3.3 |
| Skim milk | 7 | 26 | 3.71 |
| Size | Shape | Margin | Elevation | Texture | Appearance | Color | Transparency | Gram’s Reaction | Cell Shape | Cell Arrangement |
|---|---|---|---|---|---|---|---|---|---|---|
| Moderate | Irregular | Undulate | Umbonate | Rough | Dull | Cream | Opaque | Positive | Bacilli | Streptobacilli |
| Carbohydrate | Result | Antibiotic | Concentration * | Result |
|---|---|---|---|---|
| Dextrose utilization | Positive | Amoxiclav (amoxicillin/clavulanic acid) (AMC) | 30 mcg (20/10 mcg) | Sensitive |
| Esculin hydrolysis | Positive | Cefoxitin (Cephoxitin) (CX) | 30 mcg | Sensitive |
| Cellobiose utilization | Positive | Ceftazidime (CAZ) | 30 mcg | Sensitive |
| Citrate utilization | Positive | Cephalothin (CEP) | 30 mcg | Sensitive |
| Inositol utilization | Positive | Clindamycin (CD) | 2 mcg | Sensitive |
| Inulin utilization | Positive | Erythromycin (E) | 15 mcg | Sensitive |
| Mannitol utilization | Positive | Gentamicin (GEN) | 10 mcg | Sensitive |
| Sodium gluconate utilization | Positive | Ofloxacin (OF) | 5 mcg | Sensitive |
| Sorbitol utilization | Positive | Oxacillin (OX) | 1 mcg | Sensitive |
| Sucrose utilization | Positive | Penicillin-G (P) | 10 units | Sensitive |
| Trehalose utilization | Positive | Teicoplanin (TEI) | 30 mcg | Sensitive |
| Vancomycin (VA) | 30 mcg | Sensitive |
| Purification Process | Total Activity (U mL−1) | Total Protein (mg mL−1) | Specific Activity (U mg−1) | Yield (%) | Purification Fold |
|---|---|---|---|---|---|
| Culture supernatant | 313.18 | 29.70 | 10.54 | 100 | 1 |
| Ammonium sulfate precipitation | 215.63 | 9.45 | 22.81 | 68.9 | 2.2 |
| Dialysis | 160.06 | 4.15 | 38.54 | 51.1 | 3.7 |
| Gel filtration chromatography | 114.49 | 1.88 | 61.05 | 36.6 | 5.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mushtaq, H.; Jehangir, A.; Ganai, S.A.; Farooq, S.; Ganai, B.A.; Nazir, R. Biochemical Characterization and Functional Analysis of Heat Stable High Potential Protease of Bacillus amyloliquefaciens Strain HM48 from Soils of Dachigam National Park in Kashmir Himalaya. Biomolecules 2021, 11, 117. https://doi.org/10.3390/biom11010117
Mushtaq H, Jehangir A, Ganai SA, Farooq S, Ganai BA, Nazir R. Biochemical Characterization and Functional Analysis of Heat Stable High Potential Protease of Bacillus amyloliquefaciens Strain HM48 from Soils of Dachigam National Park in Kashmir Himalaya. Biomolecules. 2021; 11(1):117. https://doi.org/10.3390/biom11010117
Chicago/Turabian StyleMushtaq, Hina, Arshid Jehangir, Shabir Ahmad Ganai, Saleem Farooq, Bashir Ahmad Ganai, and Ruqeya Nazir. 2021. "Biochemical Characterization and Functional Analysis of Heat Stable High Potential Protease of Bacillus amyloliquefaciens Strain HM48 from Soils of Dachigam National Park in Kashmir Himalaya" Biomolecules 11, no. 1: 117. https://doi.org/10.3390/biom11010117
APA StyleMushtaq, H., Jehangir, A., Ganai, S. A., Farooq, S., Ganai, B. A., & Nazir, R. (2021). Biochemical Characterization and Functional Analysis of Heat Stable High Potential Protease of Bacillus amyloliquefaciens Strain HM48 from Soils of Dachigam National Park in Kashmir Himalaya. Biomolecules, 11(1), 117. https://doi.org/10.3390/biom11010117