
Histamine Ingestion by Anopheles stephensi Alters Important Vector Transmission Behaviors and Infection Success with Diverse Plasmodium Species

 ,
, 
{kind=link}
{kind=link}
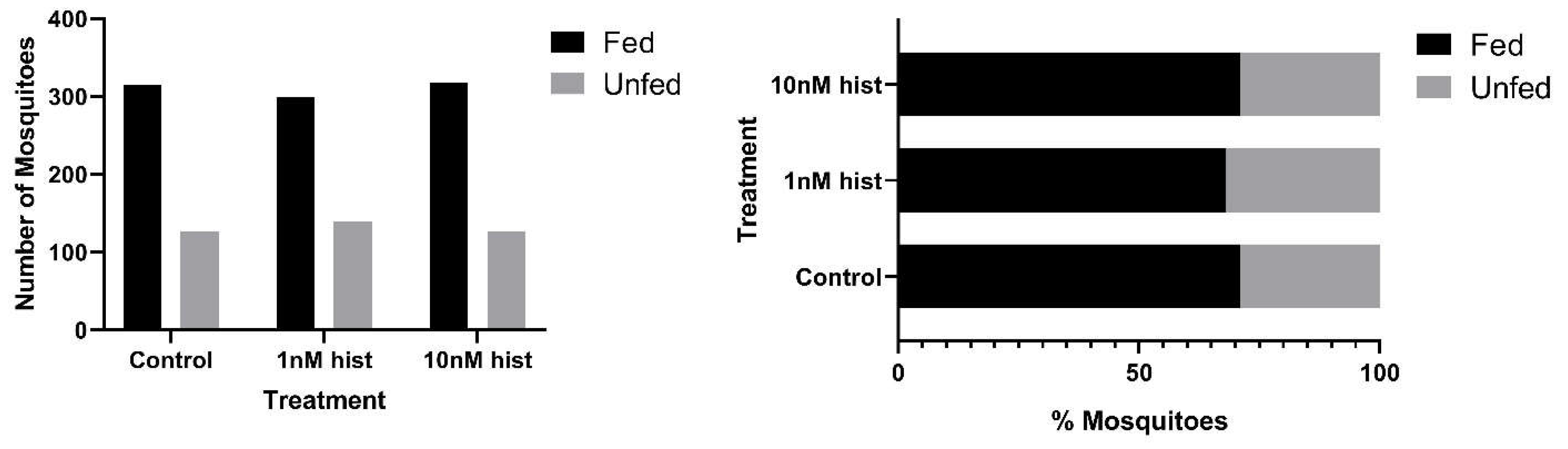{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Mosquito Rearing
2.3. Techniques
2.3.1. Histamine Priming of Female An. stephensi
2.3.2. Artificial Bloodmeal Delivery to Female An. stephensi
2.3.3. Histamine Provisioning in a Bloodmeal to Female An. stephensi
2.3.4. Mouse Infection with Plasmodium yoelii yoelii 17XNL
2.3.5. P. falciparum NF54 Culture
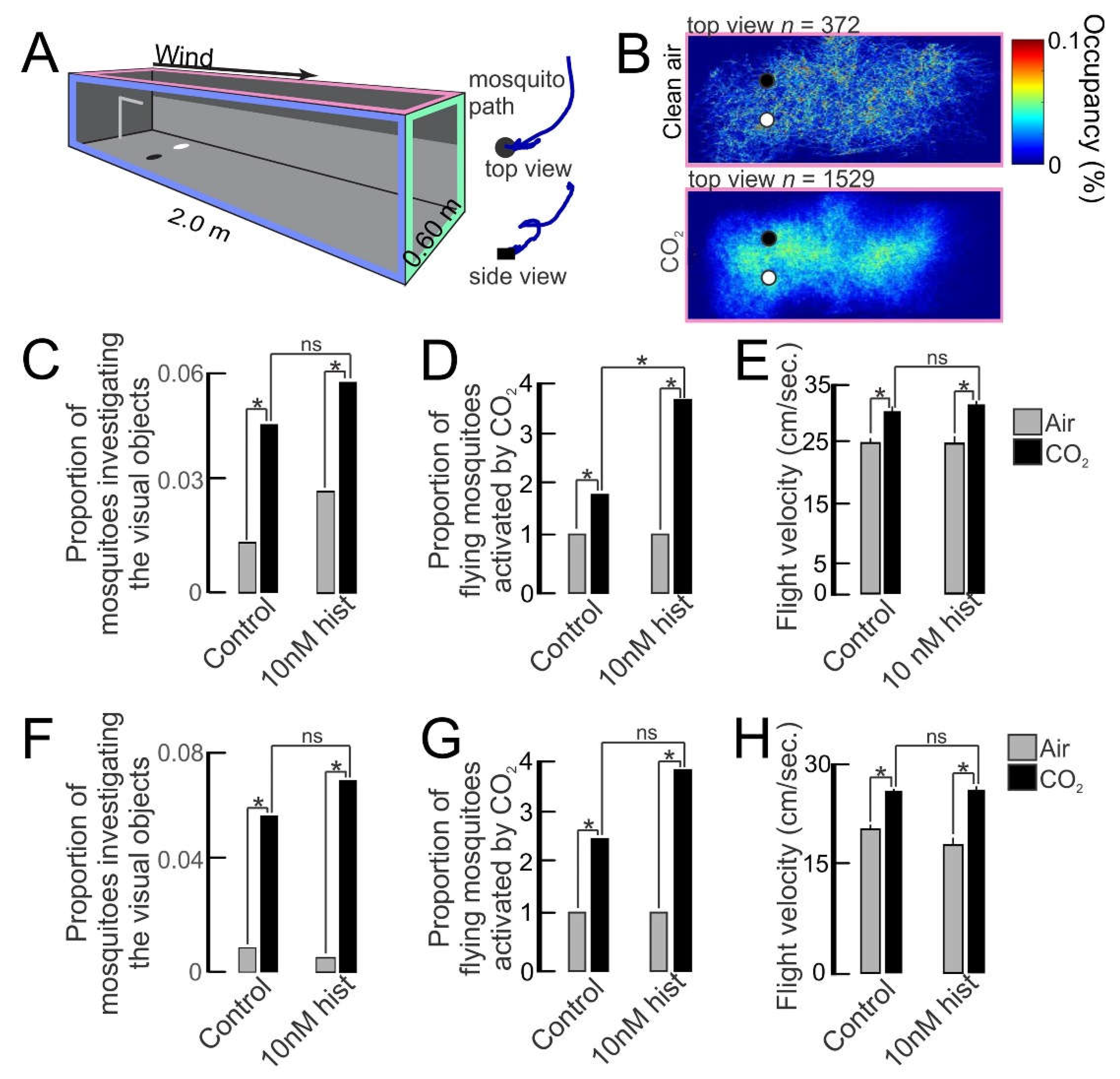
2.4. Impact of Histamine Provisioning on Uninfected Female An. stephensi Behavior
2.4.1. Tendency to Take a Second Bloodmeal
2.4.2. Flight Activity and Visual Object Investigation in Response to CO2
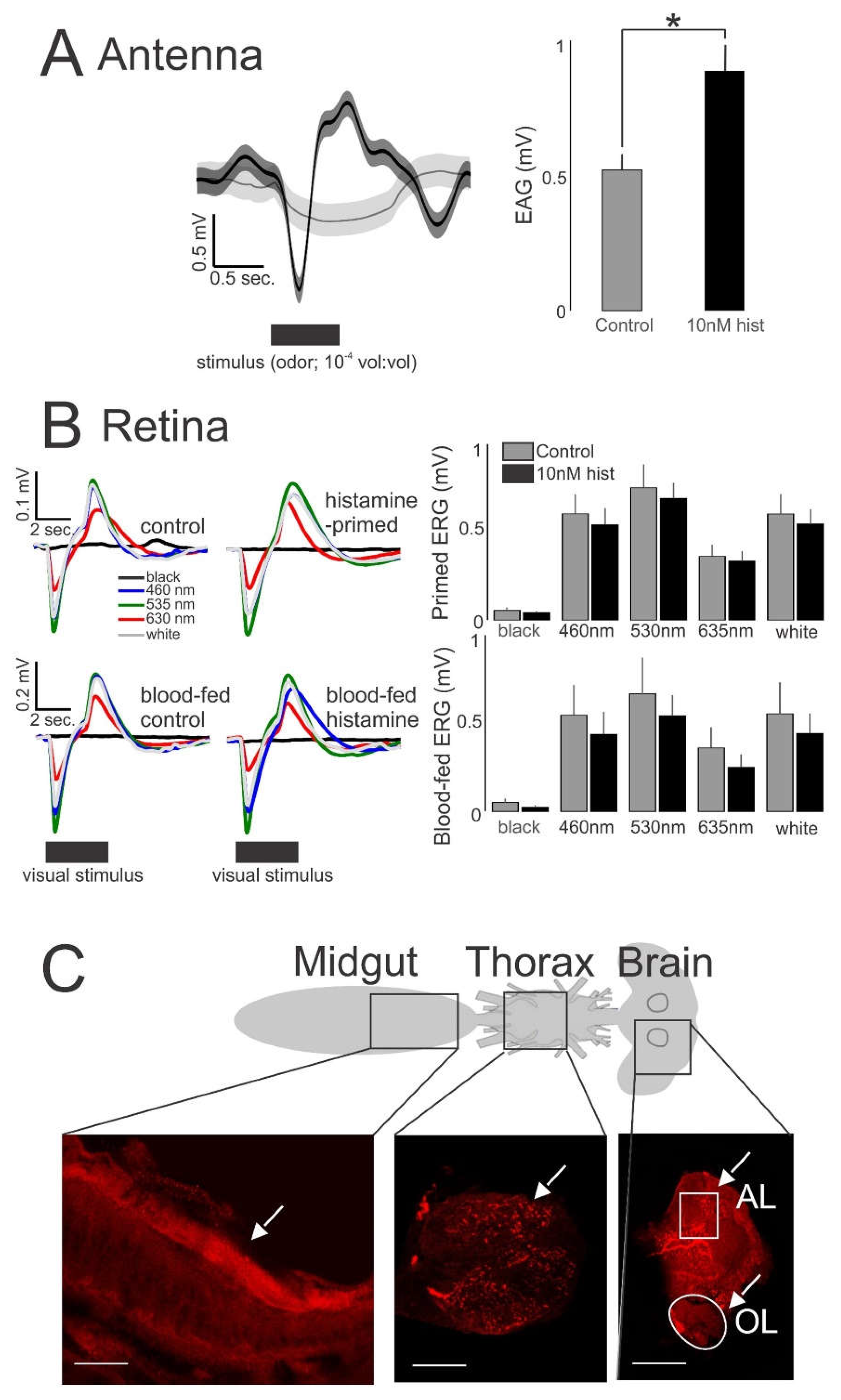
2.4.3. Electroretinogram (ERG) and Electroantennogram (EAG) Recordings of An. stephensi Response to Host-Associated Visual and Odor Stimuli
2.4.4. Histamine Immunohistochemistry
2.5. Impact of Histamine Provisioning on Lifespan and Patterns of Blood Feeding of Uninfected Female An. stephensi over Time
2.6. Impact of Histamine Provisioning on Plasmodium Infection of An. stephensi
2.6.1. P. y. yoelii 17XNL Infection of An. stephensi and Blood Feeding Behavior of Infected Female Mosquitoes
2.6.2. Impact of Histamine Treatment on P. falciparum NF54 Growth in vitro
2.6.3. P. falciparum Infection of An. stephensi
2.7. Statistical Analyses
3. Results
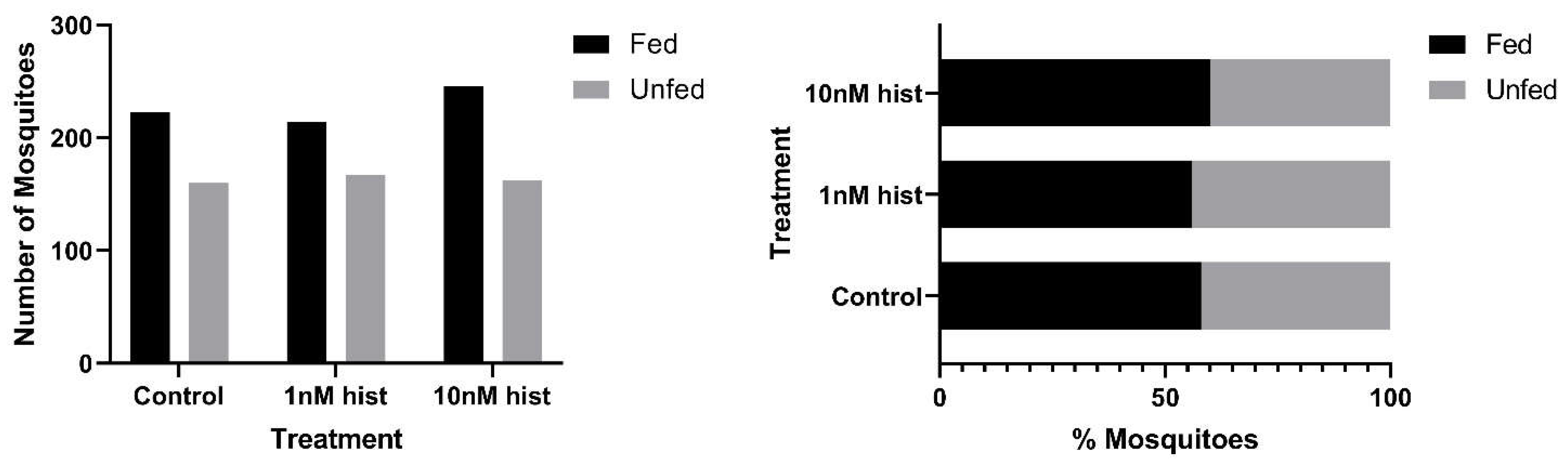
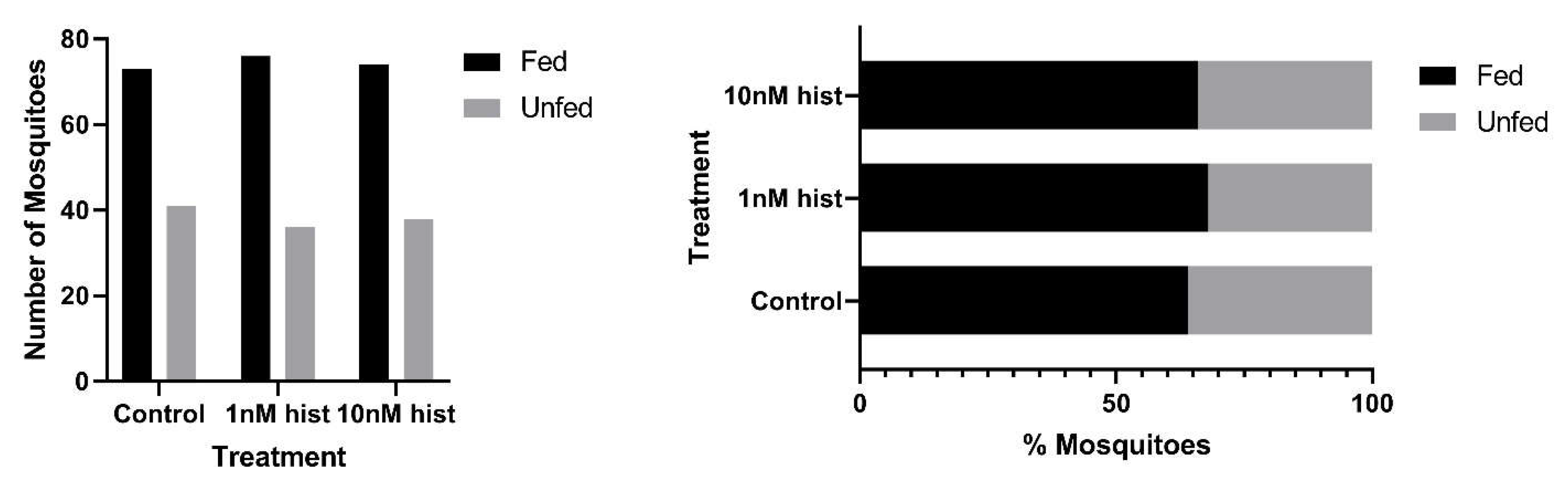
3.1. Histamine Provisioning of Uninfected An. stephensi Was Associated with Distinct Temporal Patterns in the Tendency to Take a Second Bloodmeal
3.2. Histamine Provisioning Enhanced Uninfected Female An. stephensi Flight Activity and Physiological Responses to Olfactory Cues
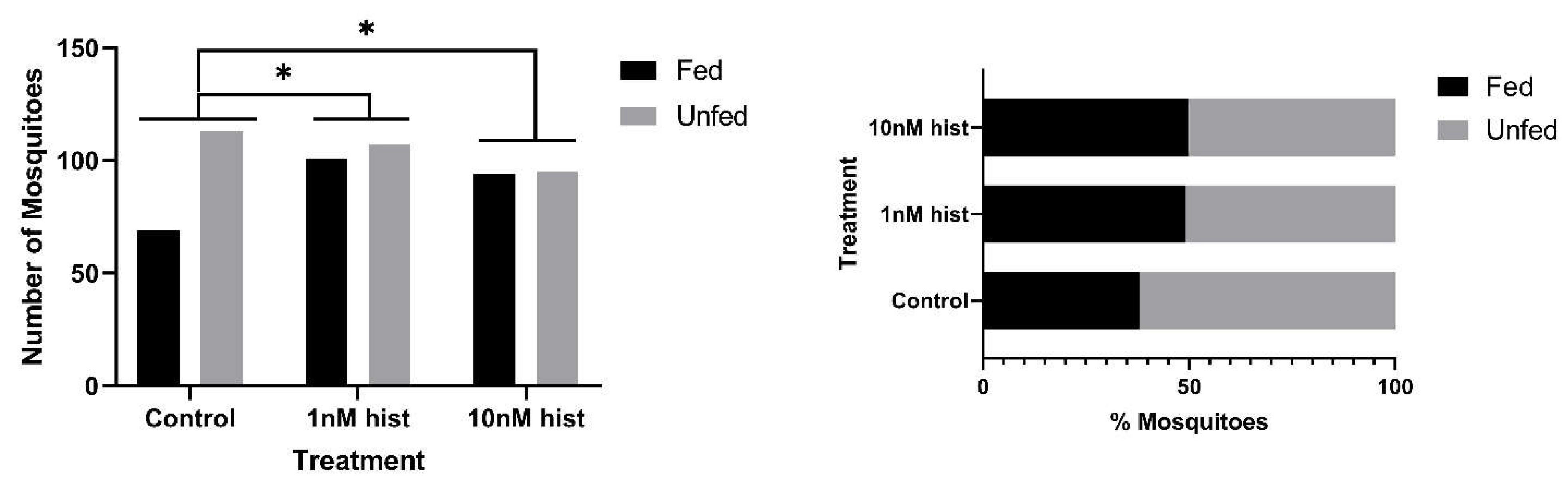
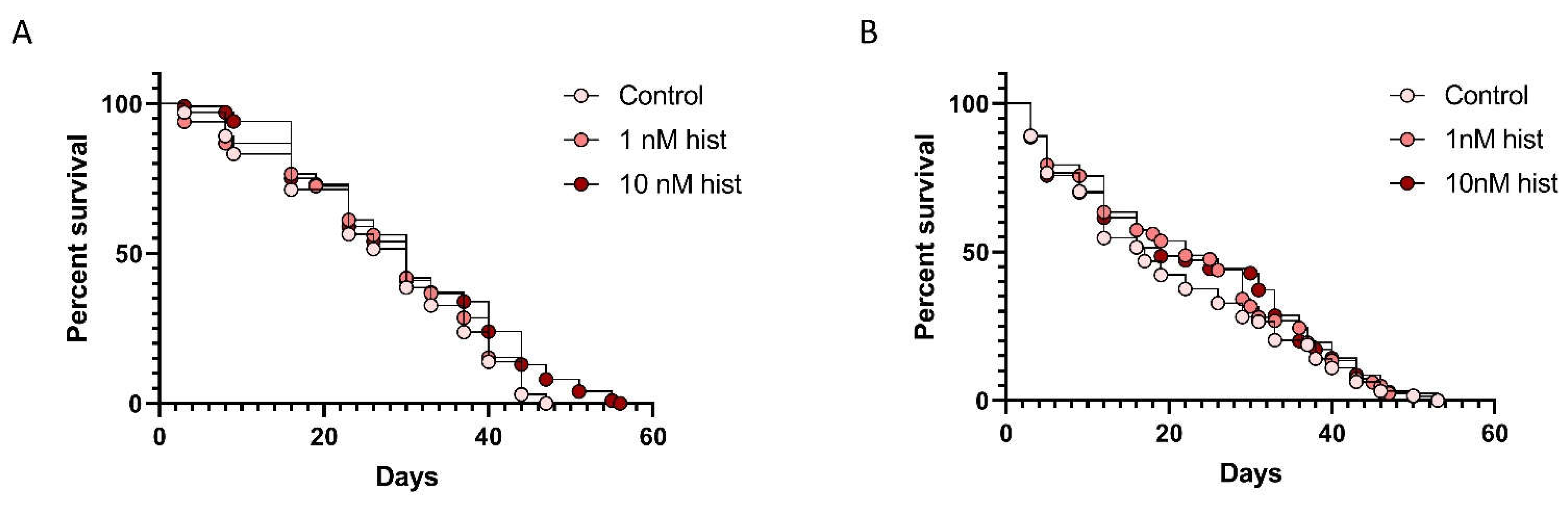
3.3. Histamine Provisioning Had Modest to No Effects on Uninfected Female An. stephensi Lifespan, But Increased the Tendency to Take Weekly Bloodmeals with Increasing Age
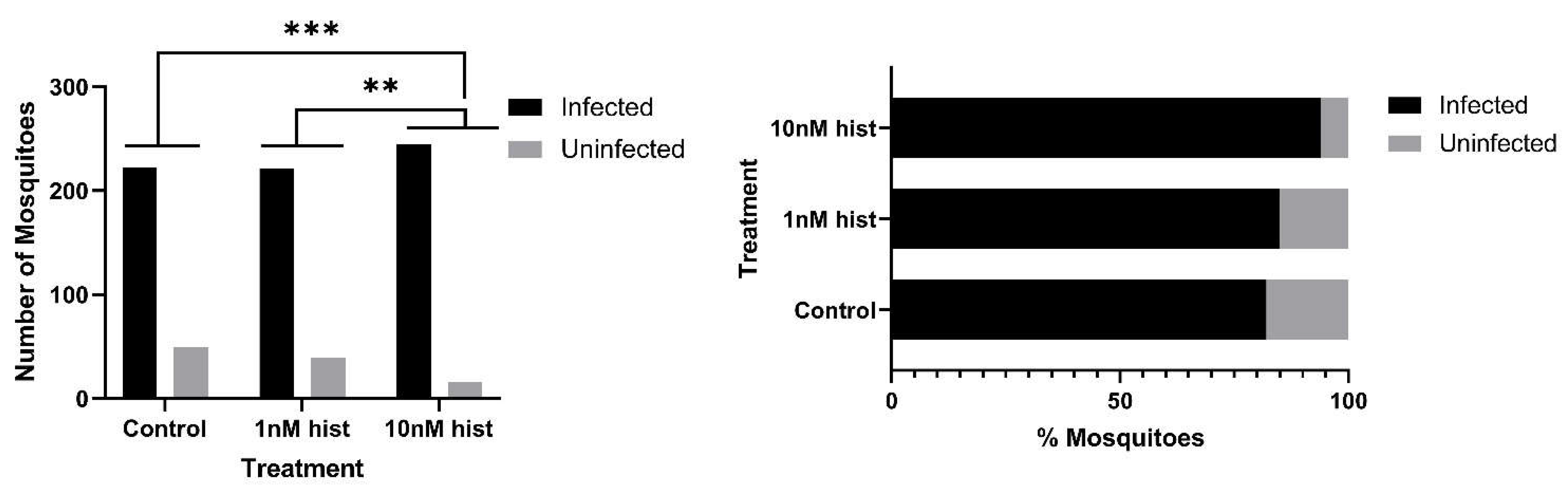
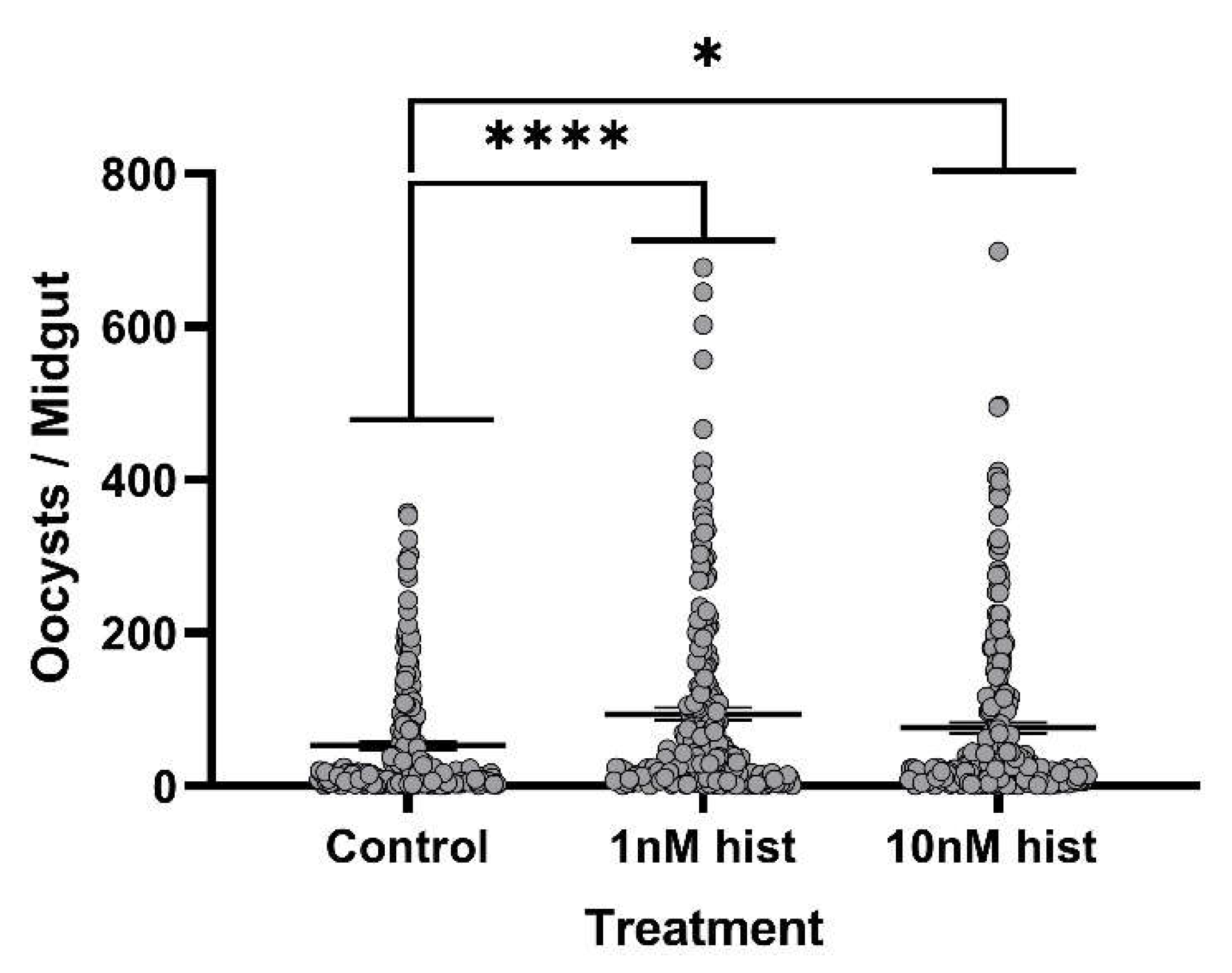
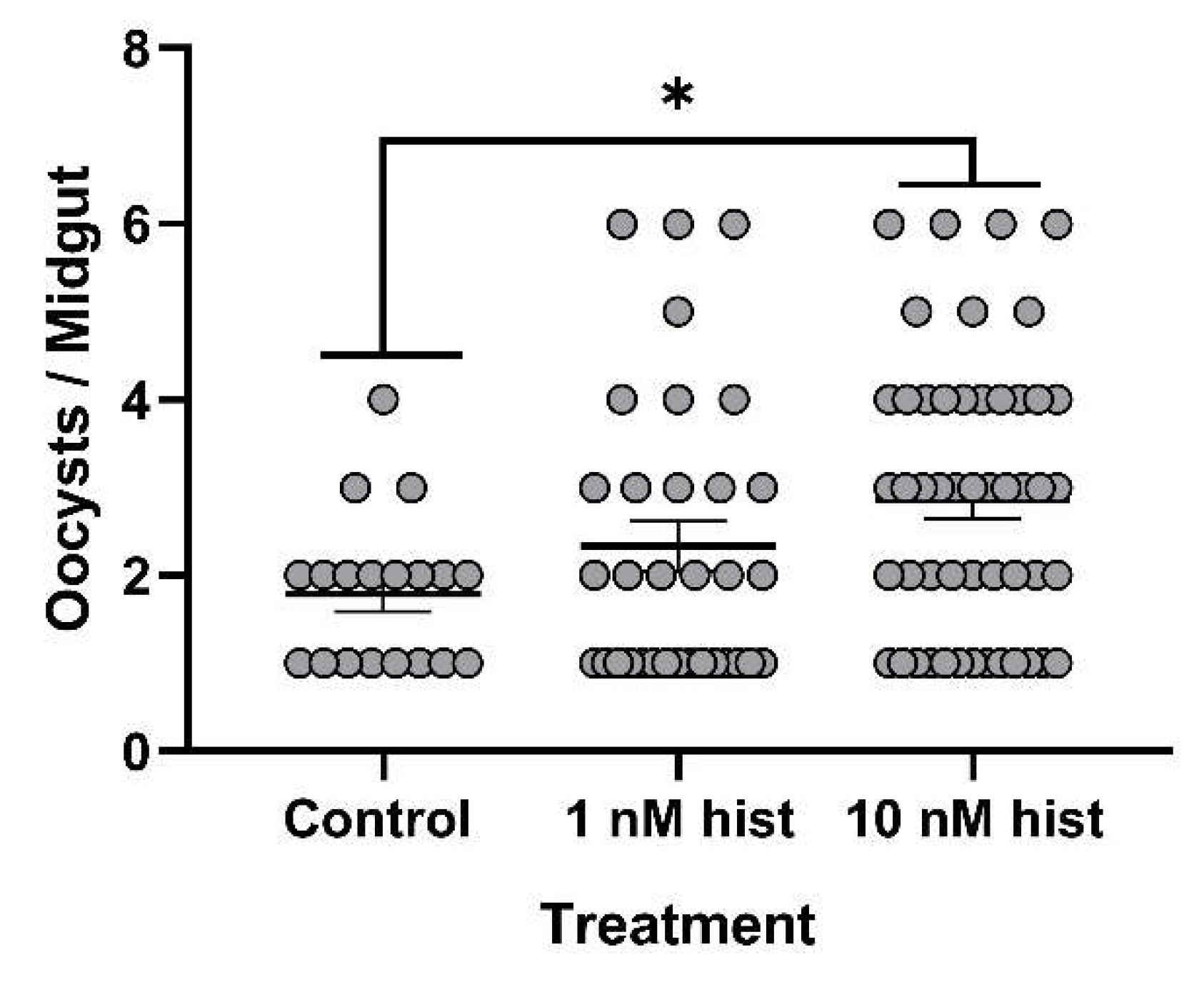
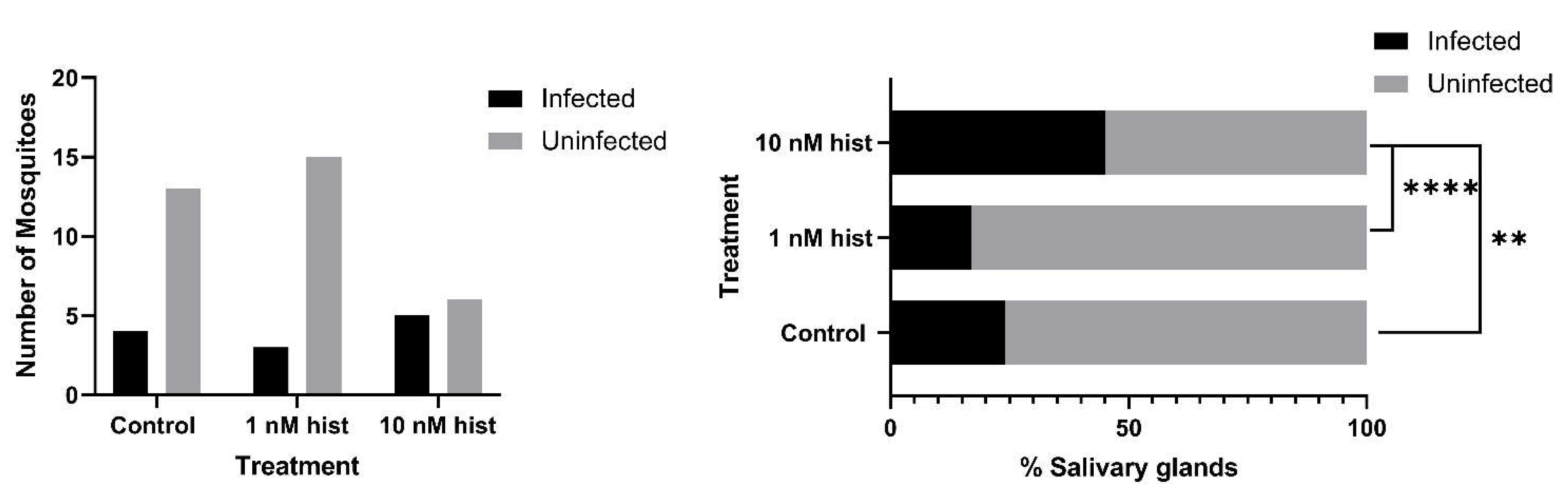
3.4. Histamine Provisioning Enhanced An. stephensi Infection with P. y. yoelii 17XNL
3.5. Histamine Provisioning Had No Effect on the Tendency of P. y. yoelii 17XNL-Infected An. stephensi to Take a Subsequent Bloodmeal
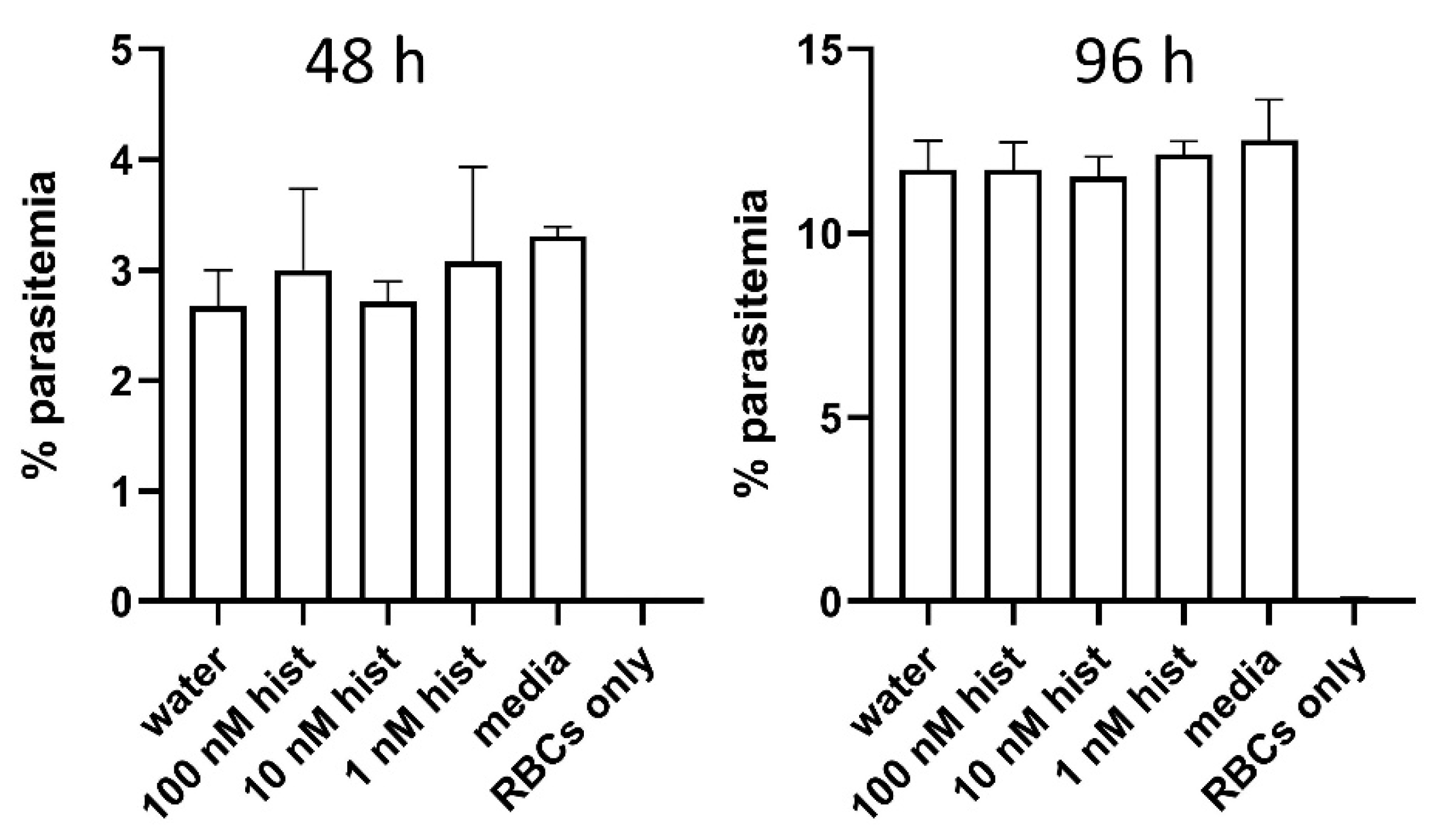
3.6. Histamine Provisioning Enhanced An. stephensi Infection with P. falciparum NF54 Strain
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. World Malaria Report 2020: 20 Years of Progress and Challenges; WHO: Geneva, Switzerland, 2020. [Google Scholar]
- Enwonwu, C.O.; Afolabi, B.M.; Salako, L.O.; Idigbe, E.O.; Bashirelahi, N. Increased Plasma Levels of Histidine and Histamine in falciparum Malaria: Relevance to Severity of Infection. J. Neural Transm. 2000, 107, 1273–1287. [Google Scholar] [CrossRef] [PubMed]
- Lopansri, B.K.; Anstey, N.M.; Stoddard, G.J.; Mwaikambo, E.D.; Boutlis, C.S.; Tjitra, E.; Maniboey, H.; Hobbs, M.R.; Levesque, M.C.; Weinberg, J.B.; et al. Elevated Plasma Phenylalanine in Severe Malaria and Implications for Pathophysiology of Neurological Complications. Infect. Immun. 2006, 75. [Google Scholar] [CrossRef] [Green Version]
- Branco, A.C.C.C.; Yoshikawa, F.S.Y.; Pietrobon, A.J.; Sato, M.N. Role of Histamine in Modulating the Immune Response and Inflammation. Mediat. Inflamm. 2018. [Google Scholar] [CrossRef]
- Giraldo-Calderón, G.I.; Emrich, S.J.; MacCallum, R.M.; Maslen, G.; Emrich, S.; Collins, F.; Dialynas, E.; Topalis, P.; Ho, N.; Gesing, S.; et al. VectorBase: An Updated Bioinformatics Resource for Invertebrate Vectors and Other Organisms Related with Human Diseases. Nucleic Acids Res. 2015, 43. [Google Scholar] [CrossRef]
- Das De, T.; Sharma, P.; Tevatiya, S.; Chauhan, C.; Kumari, S.; Singla, D.; Srivastava, V.; Rani, J.; Hasija, Y.; Pandey, K.C.; et al. Microbiome-Gut-Brain-Axis Communication Influences Metabolic Switch in the Mosquito Anopheles culicifacies. bioRxiv 2020, 774430. [Google Scholar] [CrossRef] [Green Version]
- Bradley, S.P.; Chapman, P.D.; Lizbinski, K.M.; Daly, K.C.; Dacks, A.M. A Flight Sensory-Motor to Olfactory Processing Circuit in the Moth Manduca sexta. Front. Neural Circuits 2016, 10. [Google Scholar] [CrossRef] [Green Version]
- Chapman, P.D.; Bradley, S.P.; Haught, E.J.; Riggs, K.E.; Haffar, M.M.; Daly, K.C.; Dacks, A.M. Co-Option of a Motor-to-Sensory Histaminergic Circuit Correlates with Insect Flight Biomechanics. Proc. R. Soc. B Biol. Sci. 2017, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, P.D.; Burkland, R.; Bradley, S.P.; Houot, B.; Bullman, V.; Dacks, A.M.; Daly, K.C. Flight Motor Networks Modulate Primary Olfactory Processing in the Moth Manduca sexta. Proc. Natl. Acad. Sci. USA 2018, 115, 5588–5593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchner, E.; Buchner, S.; Burg, M.G.; Hofbauer, A.; Pak, W.L.; Pollack, I. Histamine Is a Major Mechanosensory Neurotransmitter Candidate in Drosophila melanogaster. Cell Tissue Res. 1993, 273. [Google Scholar] [CrossRef]
- Borycz, J.; Vohra, M.; Tokarczyk, G.; Meinertzhagen, I.A. The Determination of Histamine in the Drosophila Head. J. Neurosci. Methods 2000, 101. [Google Scholar] [CrossRef]
- Nacher, M.; Singhasivanon, P.; Silachamroon, U.; Treeprasertsuk, S.; Tosukhowong, T.; Vannaphan, S.; Gay, F.; Mazier, D.; Looareesuwan, S. Decreased Hemoglobin Concentrations, Hyperparasitemia, and Severe Malaria Are Associated with Increased Plasmodium falciparum Gametocyte Carriage. J. Parasitol. 2002, 88. [Google Scholar] [CrossRef]
- Koepfli, C.; Robinson, L.J.; Rarau, P.; Salib, M.; Sambale, N.; Wampfler, R.; Betuela, I.; Nuitragool, W.; Barry, A.E.; Siba, P.; et al. Blood-Stage Parasitaemia and Age Determine Plasmodium falciparum and P. vivax Gametocytaemia in Papua New Guinea. PLoS ONE 2015. [Google Scholar] [CrossRef]
- Muthui, M.K.; Mogeni, P.; Mwai, K.; Nyundo, C.; Macharia, A.; Williams, T.N.; Nyangweso, G.; Wambua, J.; Mwanga, D.; Marsh, K.; et al. Gametocyte Carriage in an Era of Changing Malaria Epidemiology: A 19-Year Analysis of a Malaria Longitudinal Cohort. Wellcome Open Res. 2019, 4. [Google Scholar] [CrossRef]
- Meibalan, E.; Marti, M. Biology of Malaria Transmission. Cold Spring Harb. Perspect. Med. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepniewska, K.; Price, R.N.; Sutherland, C.J.; Drakeley, C.J.; Von Seidlein, L.; Nosten, F.; White, N.J. Plasmodium falciparum Gametocyte Dynamics in Areas of Different Malaria Endemicity. Malar. J. 2008, 7. [Google Scholar] [CrossRef] [Green Version]
- Pett, H.; Gonçalves, B.P.; Dicko, A.; Nébié, I.; Tiono, A.B.; Lanke, K.; Bradley, J.; Chen, I.; Diawara, H.; Mahamar, A.; et al. Comparison of Molecular Quantification of Plasmodium falciparum Gametocytes by Pfs25 QRT-PCR and QT-NASBA in Relation to Mosquito Infectivity. Malar. J. 2016, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adomako-Ankomah, Y.; Chenoweth, M.S.; Tocker, A.M.; Doumbia, S.; Konate, D.; Doumbouya, M.; Keita, A.S.; Anderson, J.M.; Fairhurst, R.M.; Diakite, M.; et al. Host Age and Plasmodium falciparum Multiclonality Are Associated with Gametocyte Prevalence: A 1-Year Prospective Cohort Study. Malar. J. 2017, 16. [Google Scholar] [CrossRef]
- Stone, W.; Sawa, P.; Lanke, K.; Rijpma, S.; Oriango, R.; Nyaurah, M.; Osodo, P.; Osoti, V.; Mahamar, A.; Diawara, H.; et al. A Molecular Assay to Quantify Male and Female Plasmodium falciparum Gametocytes: Results from 2 Randomized Controlled Trials Using Primaquine for Gametocyte Clearance. J. Infect. Dis. 2017, 216. [Google Scholar] [CrossRef] [Green Version]
- Eckhoff, P. P. falciparum Infection Durations and Infectiousness Are Shaped by Antigenic Variation and Innate and Adaptive Host Immunity in a Mathematical Model. PLoS ONE 2012, 7, e44950. [Google Scholar] [CrossRef] [Green Version]
- Beshir, K.B.; Sutherland, C.J.; Sawa, P.; Drakeley, C.J.; Okell, L.; Mweresa, C.K.; Omar, S.A.; Shekalaghe, S.A.; Kaur, H.; Ndaro, A.; et al. Residual Plasmodium falciparum Parasitemia in Kenyan Children after Artemisinin-Combination Therapy Is Associated with Increased Transmission to Mosquitoes and Parasite Recurrence. J. Infect. Dis. 2013, 208. [Google Scholar] [CrossRef] [PubMed]
- Nguitragool, W.; Mueller, I.; Kumpitak, C.; Saeseu, T.; Bantuchai, S.; Yorsaeng, R.; Yimsamran, S.; Maneeboonyang, W.; Sa-Angchai, P.; Chaimungkun, W.; et al. Very High Carriage of Gametocytes in Asymptomatic Low-Density Plasmodium falciparum and P. vivax Infections in Western Thailand. Parasit. Vectors 2017, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essangui, E.; Moukoko, C.E.E.; Nguedia, N.; Tchokwansi, M.; Banlanjo, U.; Maloba, F.; Fogang, B.; Donkeu, C.; Biabi, M.; Cheteug, G.; et al. Demographical, Hematological and Serological Risk Factors for Plasmodium falciparum Gametocyte Carriage in a High Stable Transmission Zone in Cameroon. PLoS ONE 2019, 14, e0216133. [Google Scholar] [CrossRef] [Green Version]
- Tadesse, F.G.; Ashine, T.; Teka, H.; Esayas, E.; Messenger, L.A.; Chali, W.; Meerstein-Kessel, L.; Walker, T.; Wolde Behaksra, S.; Lanke, K.; et al. Anopheles stephensi Mosquitoes as Vectors of Plasmodium vivax and falciparum, Horn of Africa, 2019. Emerg. Infect. Dis. 2021, 27. [Google Scholar] [CrossRef]
- Sinka, M.E.; Pironon, S.; Massey, N.C.; Longbottom, J.; Hemingway, J.; Moyes, C.L.; Willis, K.J. A New Malaria Vector in Africa: Predicting the Expansion Range of Anopheles stephensi and Identifying the Urban Populations at Risk. Proc. Natl. Acad. Sci. USA 2020, 117. [Google Scholar] [CrossRef]
- Sinka, M.E.; Bangs, M.J.; Manguin, S.; Chareonviriyaphap, T.; Patil, A.P.; Temperley, W.H.; Gething, P.W.; Elyazar, I.R.; Kabaria, C.W.; Harbach, R.E.; et al. The Dominant Anopheles Vectors of Human Malaria in the Asia-Pacific Region: Occurrence Data, Distribution Maps and Bionomic Précis. Parasites Vectors 2011, 4. [Google Scholar] [CrossRef] [Green Version]
- Faulde, M.K.; Rueda, L.M.; Khaireh, B.A. First Record of the Asian Malaria Vector Anopheles stephensi and Its Possible Role in the Resurgence of Malaria in Djibouti, Horn of Africa. Acta Trop. 2014. [Google Scholar] [CrossRef] [PubMed]
- Beghdadi, W.; Porcherie, A.; Schneider, B.S.; Dubayle, D.; Peronet, R.; Huerre, M.; Watanabe, T.; Ohtsu, H.; Louis, J.; Mécheri, S. Inhibition of Histamine-Mediated Signaling Confers Significant Protection against Severe Malaria in Mouse Models of Disease. J. Exp. Med. 2008, 205, 395–408. [Google Scholar] [CrossRef] [Green Version]
- Srichaikul, T.; Archararit, N.; Siriasawakul, T.; Virivapanich, T. Histamine Changes in Plasmodium falciparum Malaria. Trans. R. Soc. Trop. Med. Hyg 1976, 70, 36–38. [Google Scholar] [CrossRef]
- O’Donnell, A.J.; Rund, S.S.C.; Reece, S.E. Time-of-Day of Blood-Feeding: Effects on Mosquito Life History and Malaria Transmission. Parasit. Vectors 2019, 19. [Google Scholar] [CrossRef] [PubMed]
- Pakpour, N.; Corby-Harris, V.; Green, G.P.; Smithers, H.M.; Cheung, K.W.; Riehle, M.A.; Luckhart, S. Ingested Human Insulin Inhibits the Mosquito NF-ΚB-Dependent Immune Response to Plasmodium falciparum. Infect. Immun. 2012, 35, 2141–2149. [Google Scholar] [CrossRef] [Green Version]
- Souvannaseng, L.; Hun, L.V.; Baker, H.; Klyver, J.M.; Wang, B.; Pakpour, N.; Bridgewater, J.M.; Napoli, E.; Giulivi, C.; Riehle, M.A.; et al. Inhibition of JNK Signaling in the Asian Malaria Vector Anopheles stephensi Extends Mosquito Longevity and Improves Resistance to Plasmodium falciparum Infection. PLoS Pathog. 2018, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietri, J.E.; Pakpour, N.; Napoli, E.; Song, G.; Pietri, E.; Potts, R.; Cheung, K.W.; Walker, G.; Riehle, M.A.; Starcevich, H.; et al. Two Insulin-like Peptides Differentially Regulate Malaria Parasite Infection in the Mosquito through Effects on Intermediary Metabolism. Biochem. J. 2016, 3487–4503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Breugel, F.; Riffell, J.; Fairhall, A.; Dickinson, M.H. Mosquitoes Use Vision to Associate Odor Plumes with Thermal Targets. Curr. Biol. 2015, 25. [Google Scholar] [CrossRef] [Green Version]
- Stowers, J.R.; Hofbauer, M.; Bastien, R.; Griessner, J.; Higgins, P.; Farooqui, S.; Fischer, R.M.; Nowikovsky, K.; Haubensak, W.; Couzin, I.D.; et al. Virtual Reality for Freely Moving Animals. Nat. Methods 2017, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; San Alberto, D.A.; Rusch, C.; Riffell, J.A.; Montell, C. Aedes aegypti Vision-Guided Target Recognition Requires Two Redundant Rhodopsins. bioRxiv 2020. [Google Scholar] [CrossRef]
- Desjardins, R.E.; Canfield, C.J.; Haynes, J.D.; Chulay, J.D. Quantitative Assessment of Antimalarial Activity in Vitro by a Semiautomated Microdilution Technique. Antimicrob. Agents Chemother. 1979, 16. [Google Scholar] [CrossRef] [Green Version]
- Carter, R.; Ranford-Cartwright, L.; Alano, P. The Culture and Preparation of Gametocytes of Plasmodium falciparum for Immunochemical, Molecular, and Mosquito Infectivity Studies. Methods Mol. Biol. 1993, 21. [Google Scholar] [CrossRef]
- Imbahale, S.S.; Mukabana, W.R.; Orindi, B.; Githeko, A.K.; Takken, W. Variation in Malaria Transmission Dynamics in Three Different Sites in Western Kenya. J. Trop. Med. 2012, 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pombi, M.; Calzetta, M.; Guelbeogo, W.M.; Manica, M.; Perugini, E.; Pichler, V.; Mancini, E.; Sagnon, N.F.; Ranson, H.; della Torre, A. Unexpectedly High Plasmodium Sporozoite Rate Associated with Low Human Blood Index in Anopheles coluzzii from a LLIN-Protected Village in Burkina Faso. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Bousema, T.; Dinglasan, R.R.; Morlais, I.; Gouagna, L.C.; van Warmerdam, T.; Awono-Ambene, P.H.; Bonnet, S.; Diallo, M.; Coulibaly, M.; Tchuinkam, T.; et al. Mosquito Feeding Assays to Determine the Infectiousness of Naturally Infected Plasmodium falciparum Gametocyte Carriers. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Miazgowicz, K.L.; Shocket, M.S.; Ryan, S.J.; Villena, O.C.; Hall, R.J.; Owen, J.; Adanlawo, T.; Balaji, K.; Johnson, L.R.; Mordecai, E.A.; et al. Age Influences the Thermal Suitability of Plasmodium falciparum Transmission in the Asian Malaria Vector Anopheles stephensi . Proc. R. Soc. B Biol. Sci. 2020, 287. [Google Scholar] [CrossRef]
- Porter, R.J.; Laird, R.L.; Dusseau, E.M. Studies on Malarial Sporozoites. II. Effect of Age and Dosage of Sporozoites on Their Infectiousness. Exp. Parasitol. 1954, 3. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.C.; Lee, A.H.-Y.; Hancock, R.E.W. Mechanisms of the Innate Defense Regulator Peptide-1002 Anti-Inflammatory Activity in a Sterile Inflammation Mouse Model. J. Immunol. 2017, 199. [Google Scholar] [CrossRef]
- Glennon, E.K.K.; Adams, L.G.; Hicks, D.R.; Dehesh, K.; Luckhart, S. Supplementation with Abscisic Acid Reduces Malaria Disease Severity and Parasite Transmission. Am. J. Trop. Med. Hyg. 2016, 94, 1266–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Pakpour, N.; Napoli, E.; Drexler, A.; Glennon, E.K.K.; Surachetpong, W.; Cheung, K.; Aguirre, A.; Klyver, J.M.; Lewis, E.E.; et al. Anopheles stephensi P38 MAPK Signaling Regulates Innate Immunity and Bioenergetics during Plasmodium falciparum Infection. Parasit. Vectors 2015, 8. [Google Scholar] [CrossRef] [Green Version]
- Pakpour, N.; Camp, L.; Smithers, H.M.; Wang, B.; Tu, Z.; Nadler, S.A.; Luckhart, S. Protein Kinase C-Dependent Signaling Controls the Midgut Epithelial Barrier to Malaria Parasite Infection in Anopheline Mosquitoes. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Surachetpong, W.; Singh, N.; Cheung, K.W.; Luckhart, S. MAPK ERK Signaling Regulates the TGF-Beta1-Dependent Mosquito Response to Plasmodium falciparum. PLoS Pathog. 2009, 5. [Google Scholar] [CrossRef] [Green Version]
- Carlton, J.M.; Angiuoli, S.V.; Suh, B.B.; Kooij, T.W.; Pertea, M.; Silva, J.C.; Ermolaeva, M.D.; Allen, J.E.; Selengut, J.D.; Koo, H.L.; et al. Genome Sequence and Comparative Analysis of the Model Rodent Malaria Parasite Plasmodium yoelii yoelii. Nature 2002, 419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.L.; Musiime, A.K.; Maxwell, K.; Lindsay, S.W.; Kiware, S. A New Test of a Theory about Old Mosquitoes. Trends Parasitol. 2021, 37. [Google Scholar] [CrossRef]
- Kwon, H.; Reynolds, R.A.; Simões, M.L.; Dimopoulos, G.; Smith, R.C. Malaria Parasite Immune Evasion and Adaptation to Its Mosquito Host Is Influenced by the Acquisition of Multiple Blood Meals. bioRxiv 2019, 6. [Google Scholar] [CrossRef]
- Shaw, W.R.; Holmdahl, I.E.; Itoe, M.A.; Werling, K.; Marquette, M.; Paton, D.G.; Singh, N.; Buckee, C.O.; Childs, L.M.; Catteruccia, F. Multiple Blood Feeding in Mosquitoes Shortens the Plasmodium falciparum Incubation Period and Increases Malaria Transmission Potential. PLoS Pathog. 2020, 16. [Google Scholar] [CrossRef]
- Larson, D.; Cooper, P.J.; Hübner, M.P.; Reyes, J.; Vaca, M.; Chico, M.; Kong, H.H.; Mitre, E. Helminth Infection Is Associated with Decreased Basophil Responsiveness in Human Beings. J. Allergy Clin. Immunol. 2012, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, F.; Huang, S. The Roles of Mast Cells in Parasitic Protozoan Infections. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangweme, D.T.; Midzi, N.; Zinyowera-Mutapuri, S.; Mduluza, T.; Diener-West, M.; Kumar, N. Impact of Schistosome Infection on Plasmodium falciparum Malariometric Indices and Immune Correlates in School Age Children in Burma Valley, Zimbabwe. PLoS Negl. Trop. Dis. 2010, 4. [Google Scholar] [CrossRef] [Green Version]
- Nacher, M.; Singhasivanon, P.; Silachamroon, U.; Treeprasertsuk, S.; Krudsood, S.; Gay, F.; Mazier, D.; Looareesuwan, S. Association of Helminth Infections with Increased Gametocyte Carriage during Mild falciparum Malaria in Thailand. Am. J. Trop. Med. Hyg. 2001, 65. [Google Scholar] [CrossRef] [Green Version]
- Ateba-Ngoa, U.; Jones, S.; Zinsou, J.F.; Honkpehedji, J.; Adegnika, A.A.; Agobe, J.C.D.; Massinga-Loembe, M.; Mordmüller, B.; Bousema, T.; Yazdanbakhsh, M. Associations between Helminth Infections, Plasmodium falciparum Parasite Carriage and Antibody Responses to Sexual and Asexual Stage Malarial Antigens. Am. J. Trop. Med. Hyg. 2016, 95. [Google Scholar] [CrossRef] [PubMed]
- Céspedes, N.; Donnelly, E.; Garrison, S.; Haapanen, L.; van de Water, J.; Luckhart, S. Nonlethal Plasmodium yoelii Infection Drives Complex Patterns of Th2-Type Host Immunity and Mast Cell-Dependent Bacteremia. Infect. Immun. 2020, 88. [Google Scholar] [CrossRef] [PubMed]
- Chau, J.Y.; Tiffany, C.M.; Nimishakavi, S.; Lawrence, J.A.; Pakpour, N.; Mooney, J.P.; Lokken, K.L.; Caughey, G.H.; Tsolis, R.M.; Luckhart, S. Malaria-Associated l-Arginine Deficiency Induces Mast Cell-Associated Disruption to Intestinal Barrier Defenses against Nontyphoidal Salmonella Bacteremia. Infect. Immun. 2013, 81, 3515–3526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potts, R.A.; Tiffany, C.M.; Pakpour, N.; Lokken, K.L.; Tiffany, C.R.; Cheung, K.; Tsolis, R.M.; Luckhart, S. Mast Cells and Histamine Alter Intestinal Permeability during Malaria Parasite Infection. Immunobiology 2016, 221, 468–474. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Gowda, N.M.; Kawasawa, Y.I.; Gowda, D.C. A Malaria Protein Factor Induces IL-4 Production by Dendritic Cells via PI3K–Akt–NF-B Signaling Independent of MyD88/ TRIF and Promotes Th2 Response. J. Biol. Chem. 2018, 293. [Google Scholar] [CrossRef] [Green Version]
- Urban, J.F.; Fayer, R.; Sullivan, C.; Goldhill, J.; Shea-Donohue, T.; Madden, K.; Morris, S.C.; Katona, I.; Gause, W.; Ruff, M.; et al. Local TH1 and TH2 Responses to Parasitic Infection in the Intestine: Regulation by IFN-Gamma and IL-4. Vet. Immunol. Immunopathol. 1996, 54. [Google Scholar] [CrossRef]
- Westwood, M.L.; O’Donnell, A.J.; Schneider, P.; Albery, G.F.; Prior, K.F.; Reece, S.E. Testing Possible Causes of Gametocyte Reduction in Temporally Out-of-Synch Malaria Infections. Malar. J. 2020, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Naotunne, S.T.; Karunaweera, N.D.; Del Giudice, G.; Kularatne, M.U.; Grau, G.E.; Carter, R.; Mendis, K.N. Cytokines Kill Malaria Parasites during Infection Crisis: Extracellular Complementary Factors Are Essential. J. Exp. Med. 1991, 173. [Google Scholar] [CrossRef] [PubMed]
- Long, G.H.; Chan, B.H.K.; Allen, J.E.; Read, A.F.; Graham, A.L. Blockade of TNF Receptor 1 Reduces Disease Severity but Increases Parasite Transmission during Plasmodium chabaudi chabaudi Infection. Int. J. Parasitol. 2008, 38. [Google Scholar] [CrossRef] [PubMed]
- Traore, M.M.; Junnila, A.; Traore, S.F.; Doumbia, S.; Revay, E.E.; Kravchenko, V.D.; Schlein, Y.; Arheart, K.L.; Gergely, P.; De Xue, R.; et al. Large-Scale Field Trial of Attractive Toxic Sugar Baits (ATSB) for the Control of Malaria Vector Mosquitoes in Mali, West Africa. Malar. J. 2020, 19. [Google Scholar] [CrossRef] [PubMed]



















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez, A.M.; Hambly, M.G.; Jandu, S.; Simão-Gurge, R.; Lowder, C.; Lewis, E.E.; Riffell, J.A.; Luckhart, S. Histamine Ingestion by Anopheles stephensi Alters Important Vector Transmission Behaviors and Infection Success with Diverse Plasmodium Species. Biomolecules 2021, 11, 719. https://doi.org/10.3390/biom11050719
Rodriguez AM, Hambly MG, Jandu S, Simão-Gurge R, Lowder C, Lewis EE, Riffell JA, Luckhart S. Histamine Ingestion by Anopheles stephensi Alters Important Vector Transmission Behaviors and Infection Success with Diverse Plasmodium Species. Biomolecules. 2021; 11(5):719. https://doi.org/10.3390/biom11050719
Chicago/Turabian StyleRodriguez, Anna M., Malayna G. Hambly, Sandeep Jandu, Raquel Simão-Gurge, Casey Lowder, Edwin E. Lewis, Jeffrey A. Riffell, and Shirley Luckhart. 2021. "Histamine Ingestion by Anopheles stephensi Alters Important Vector Transmission Behaviors and Infection Success with Diverse Plasmodium Species" Biomolecules 11, no. 5: 719. https://doi.org/10.3390/biom11050719
APA StyleRodriguez, A. M., Hambly, M. G., Jandu, S., Simão-Gurge, R., Lowder, C., Lewis, E. E., Riffell, J. A., & Luckhart, S. (2021). Histamine Ingestion by Anopheles stephensi Alters Important Vector Transmission Behaviors and Infection Success with Diverse Plasmodium Species. Biomolecules, 11(5), 719. https://doi.org/10.3390/biom11050719