
Characterising the Response of Human Breast Cancer Cells to Polyamine Modulation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Procedures
2.2.1. Cell Culture
2.2.2. Growth Determination
2.2.3. Protein Quantification
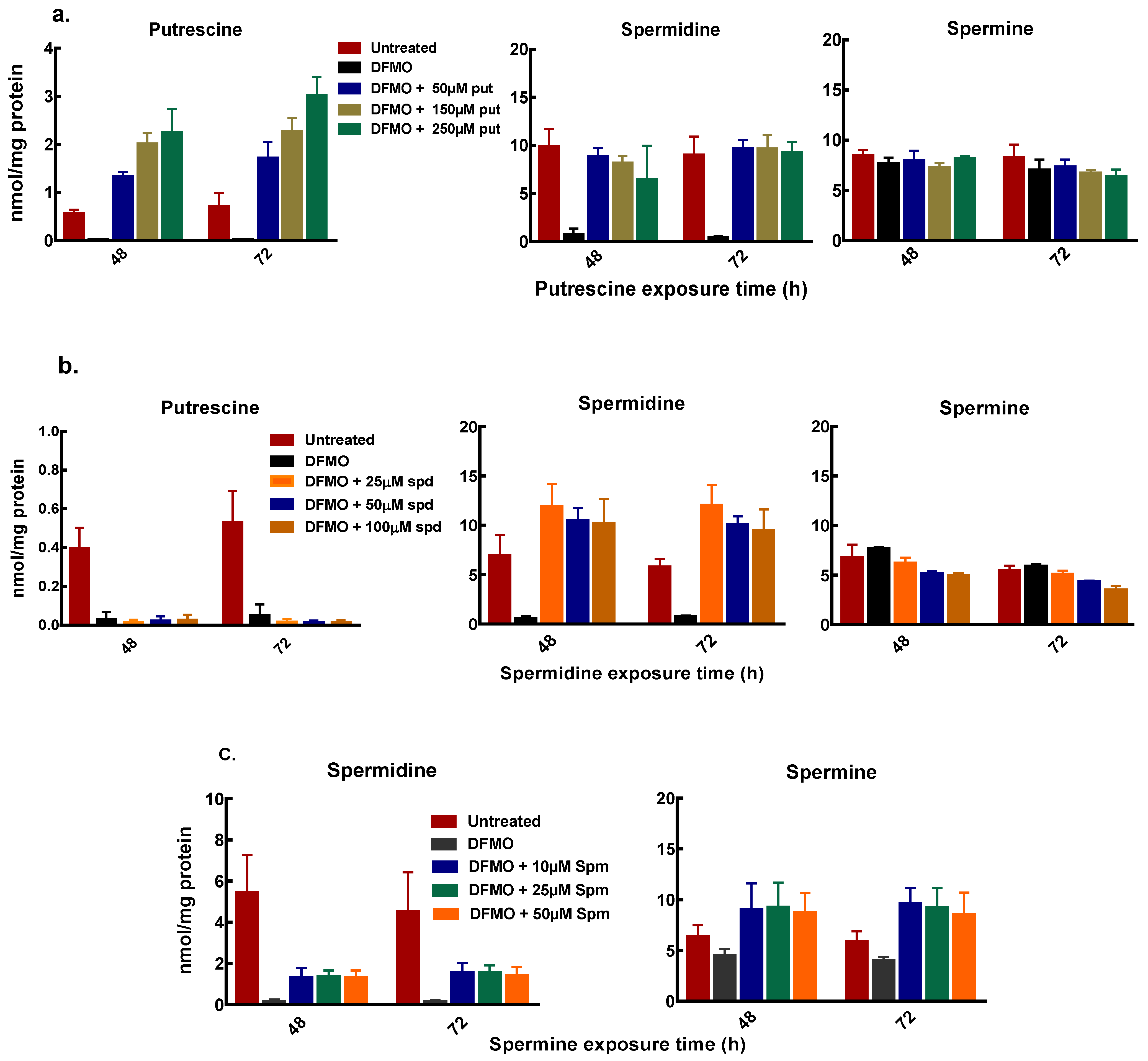
2.2.4. Polyamine Quantification
2.2.5. Quantification of Intracellular DFMO
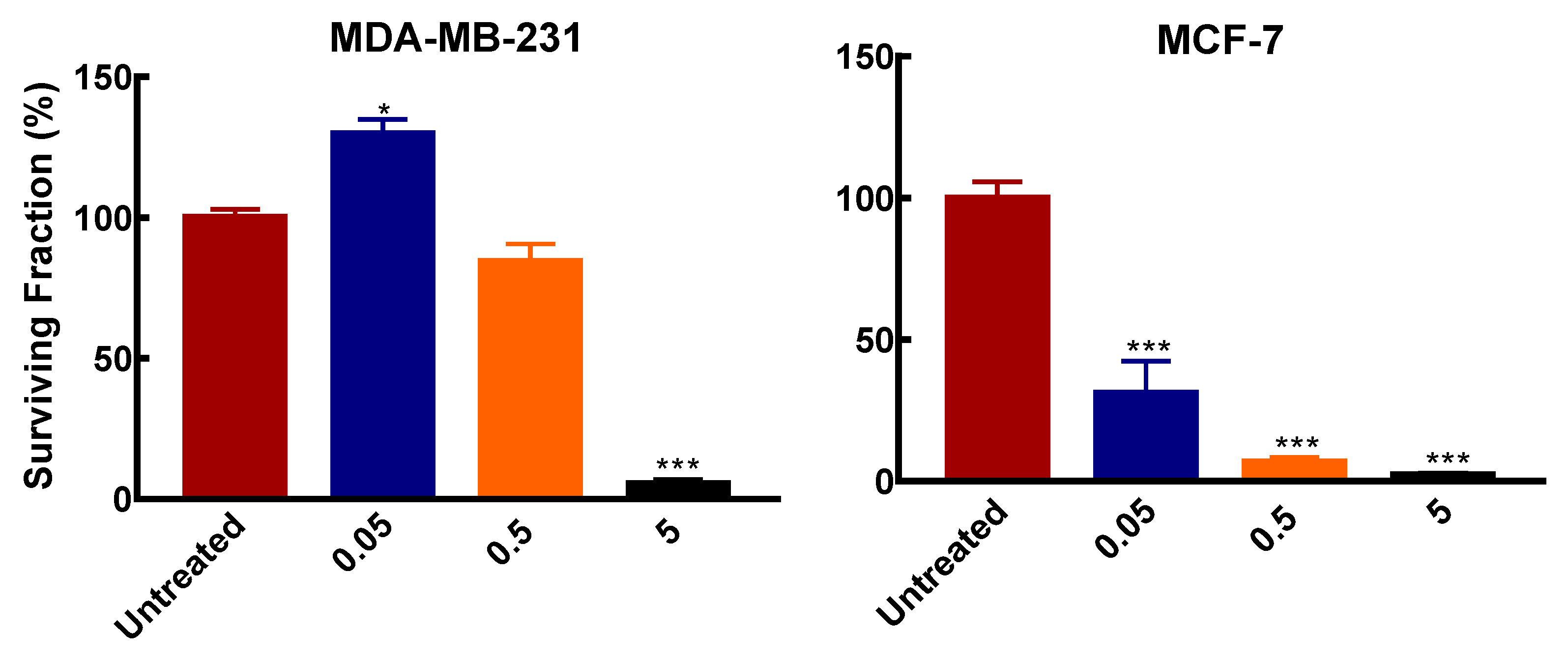
2.2.6. Colony Formation Assay
2.2.7. Western Blot
2.2.8. Polysome Analysis
2.3. Statistical Analysis
3. Results
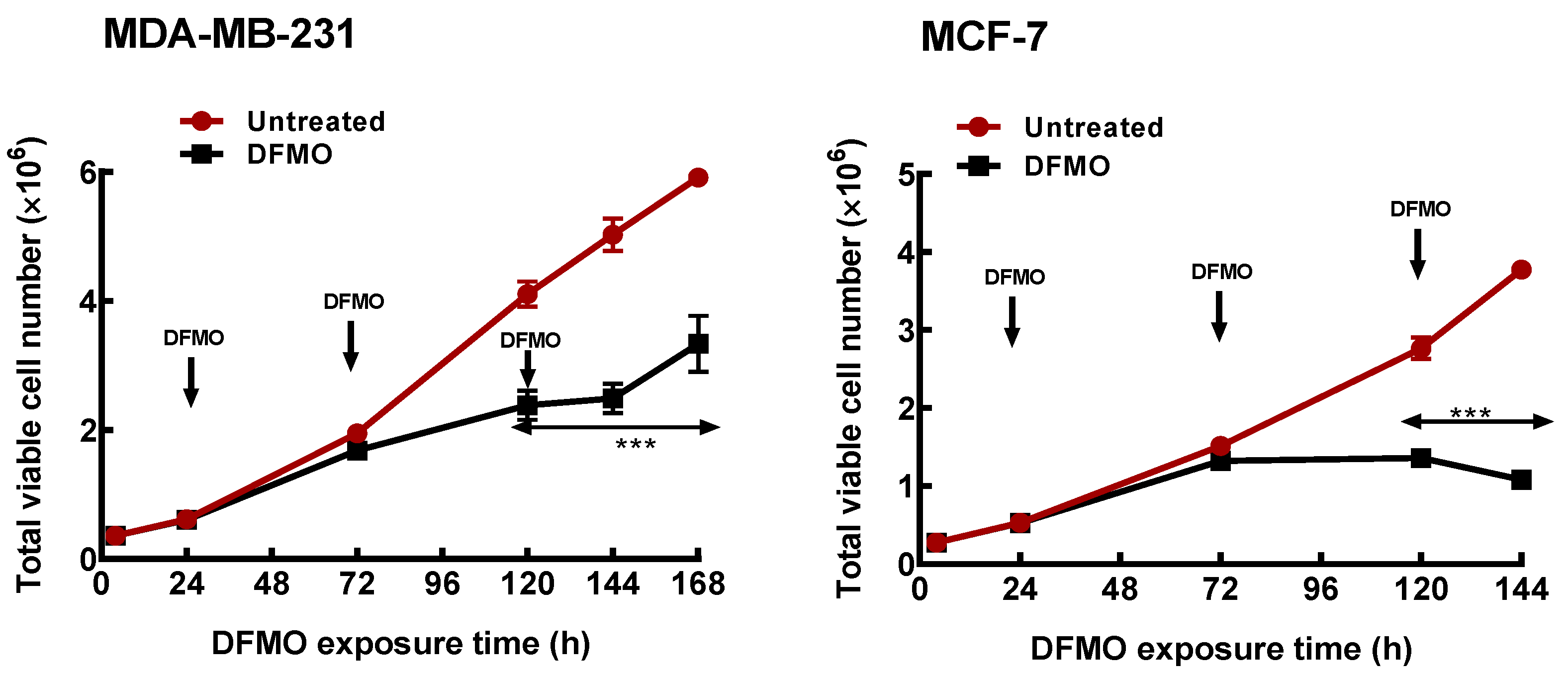
3.1. Polyamine Pathway Inhibition Affects Breast Cancer Subtypes Differentially
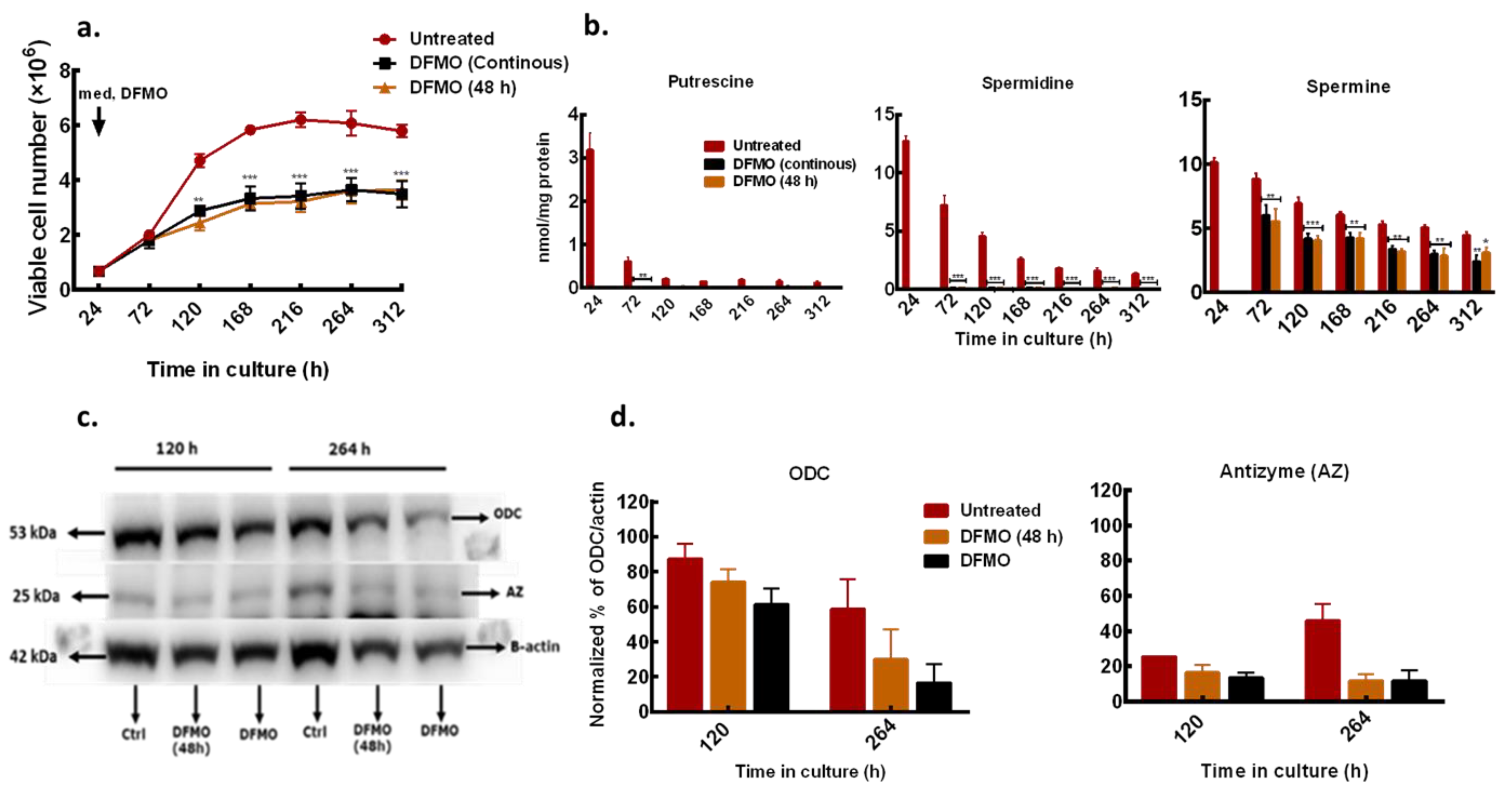
3.2. DFMO Withdrawal Fails to Rescue Cell Growth Inhibition
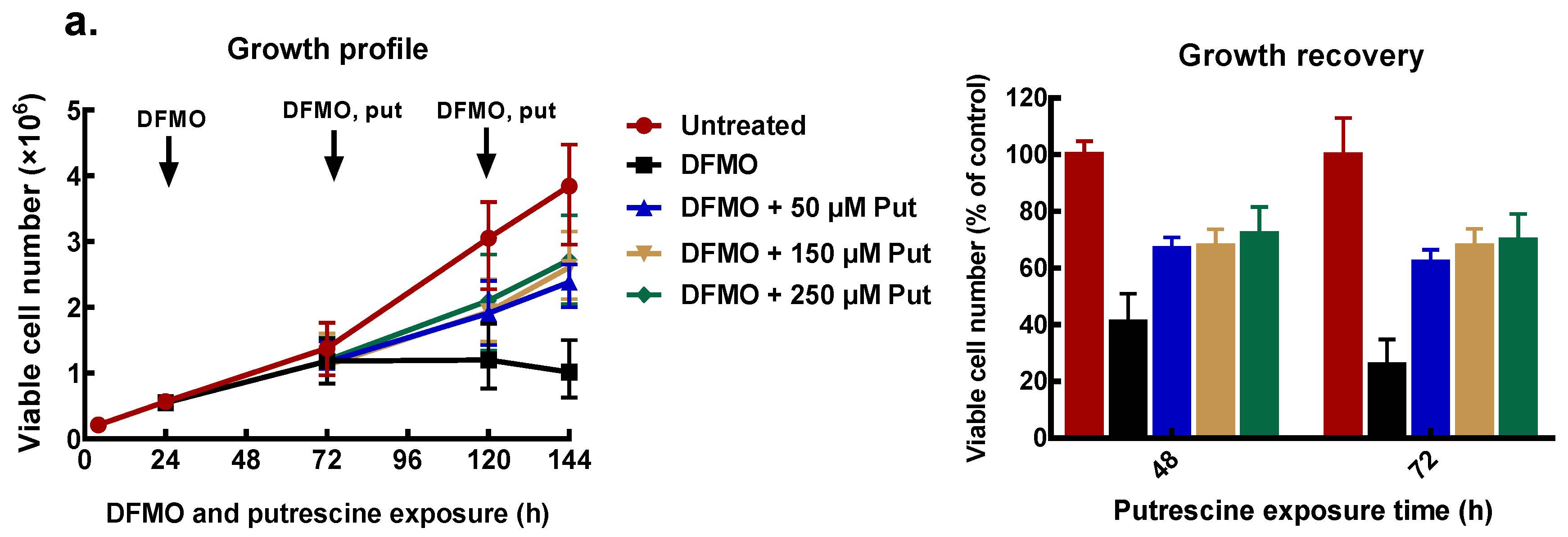
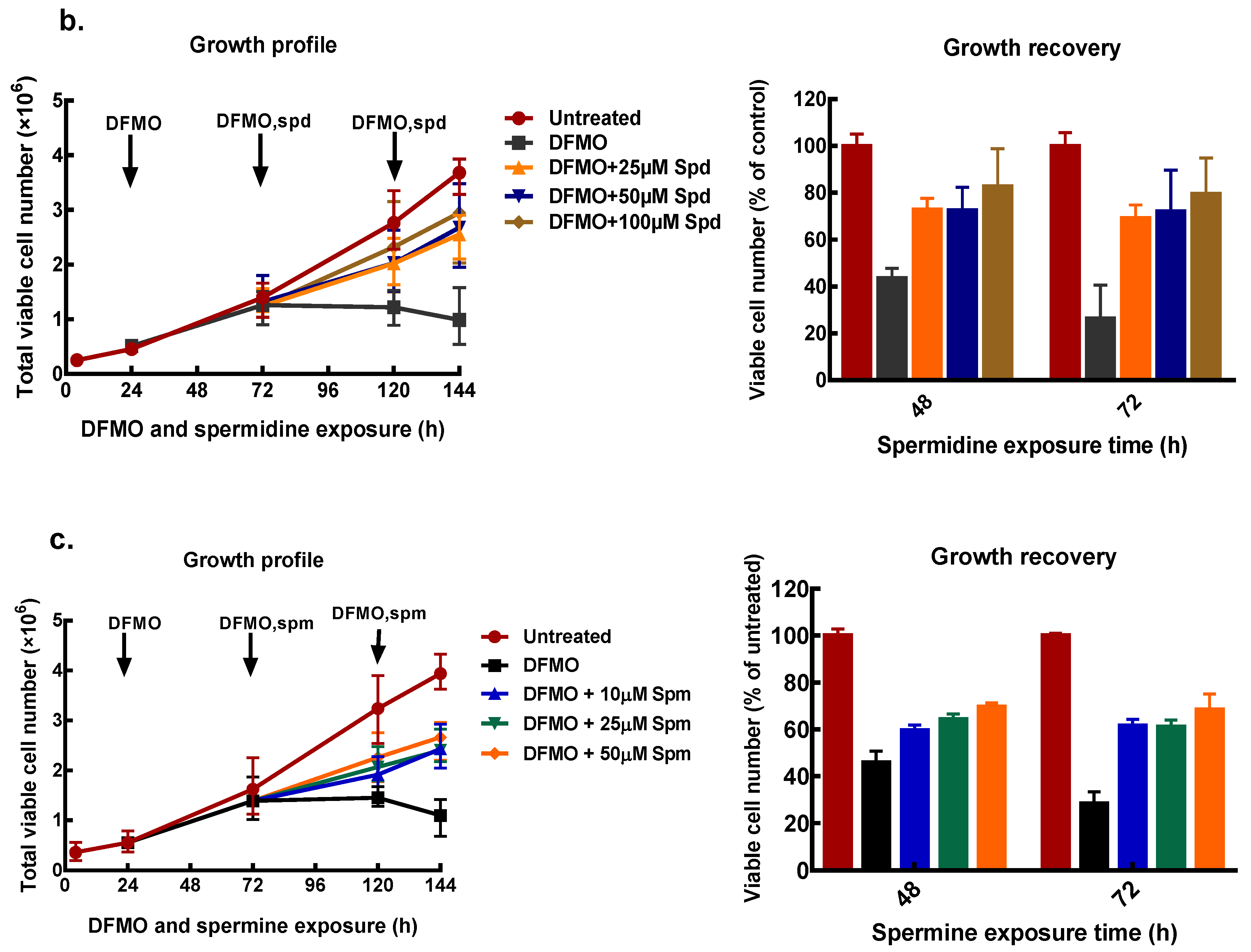
3.3. Exogenous Polyamines Reversed Growth Inhibition
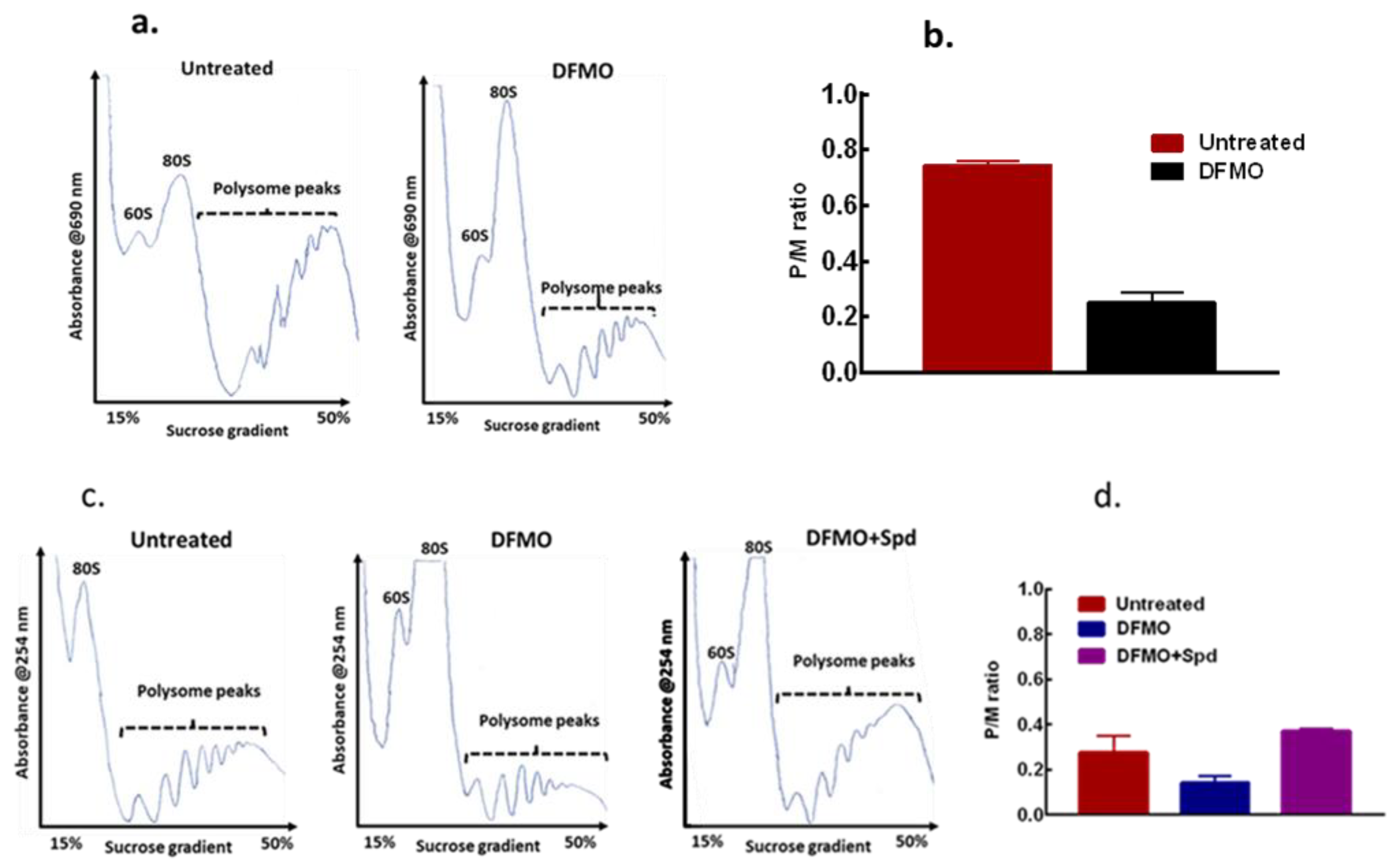
3.4. Exogenous Polyamines Reversed Translation Inhibition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, G.N.; Dave, R.; Sanadya, J.; Sharma, P.; Sharma, K.K. Various types and management of breast cancer: An overview. J. Adv. Pharm. Technol. Res. 2010, 1, 109–126. [Google Scholar]
- Gandhi, N.; Das, G.M. Metabolic Reprogramming in Breast Cancer and Its Therapeutic Implications. Cells 2019, 8, 89. [Google Scholar] [CrossRef] [Green Version]
- Ortega, M.A.; Fraile-Martínez, O.; Asúnsolo, Á.; Buján, J.; García-Honduvilla, N.; Coca, S. Signal Transduction Pathways in Breast Cancer: The Important Role of PI3K/Akt/mTOR. J. Oncol. 2020, 2020, 9258396. [Google Scholar] [CrossRef] [Green Version]
- Cha, Y.J.; Kim, E.S.; Koo, J.S. Amino Acid Transporters and Glutamine Metabolism in Breast Cancer. Int. J. Mol. Sci. 2018, 19, 907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manni, A.; Wechter, R.; Verderame, M.F.; Mauger, D. Cooperativity between the polyamine pathway and HER-2neu in transformation of human mammary epithelial cells in culture: Role of the MAPK pathway. Int. J. Cancer 1998, 76, 563–570. [Google Scholar] [CrossRef]
- Cervelli, M.; Pietropaoli, S.; Signore, F.; Amendola, R.; Mariottini, P. Polyamine metabolism and breast cancer: State of the art and perspectives. Breast Cancer Res. Treat. 2014, 148, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Thomas, T.J. Estradiol control of ornithine decarboxylase mRNA, enzyme activity, and polyamine levels in MCF-7 breast cancer cells: Therapeutic implications. Breast Cancer Res. Treat. 1994, 29, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Manni, A.; Badger, B.; Glikman, P.; Bartholomew, M.; Santner, S.; Demers, L. Individual and combined effects of alpha-difluoromethylornithine and ovariectomy on the growth and polyamine milieu of experimental breast cancer in rats. Cancer Res. 1989, 49, 3529–3534. [Google Scholar] [PubMed]
- Soda, K. The Mechanism by which Polyamines Accelerate Tumour Spread. J. Exp. Clin. Cancer Res. 2011, 30, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jänne, J.; Poso, H.; Raina, A. Polyamines in Rapid Growth and Cancer. Biochim. Biophys. Acta 1978, 473, 241–293. [Google Scholar] [CrossRef]
- Kingsnorth, A.N.; Lumsden, A.B.; Wallace, H.M. Polyamines in Colorectal Cancer. Br. J. Surg. 1984, 71, 791–794. [Google Scholar] [CrossRef] [PubMed]
- Russell, D.H. Increased polyamine concentrations in the urine of human cancer patients. Nat. New Biol. 1971, 233, 144–145. [Google Scholar] [CrossRef] [PubMed]
- Wallace, H.M.; Duthie, J.; Evans, D.M.; Lamond, S.; Nicoll, K.M.; Heys, S.D. Alterations in polyamine catabolic enzymes in human breast cancer tissue. Clin. Cancer Res. 2000, 6, 3657–3661. [Google Scholar]
- Kingsnorth, A.N.; Wallace, H.M.; Bunder, N.J.; Dixon, J.M.J. Polyamines in Breast Cancer. British J. Surg. 1984, 71, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Linsalata, M.; Caruso, M.G.; Leo, S.; Guerra, V.; D’Attoma, B.; Di Leo, A. Prognostic Value of Tissue Polyamine Levels in Human Colorectal Carcinoma. Anticancer Res. 2002, 22, 2465–2469. [Google Scholar] [PubMed]
- Manni, A.; Mauger, D.; Gimothy, P.; Badger, B. Prognostic Influence on Survival of Increased Ornithine Decarboxylase Activity in Human Breast Cancer. Clin. Cancer Res. 1999, 2, 1901–1906. [Google Scholar]
- Weiss, T.S.; Bernhardt, G.; Buschauer, A.; Wolfgang, E.T.; Dolgner, D.; Zirngibl, H.; Jauch, K. Polyamine Levels of Human Colorectal Adenocarcinomas are Correlated with Tumour Stage and Grade. Int. J. Colorectal. Dis. 2002, 17, 381–387. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Li, J.; Cameron, G.A.; Wallace, H.M. Decreased sensitivity to aspirin is associated with altered polyamine metabolism in human prostate cancer cells. Amino. Acids. 2015, 48, 1003–1012. [Google Scholar] [CrossRef]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haverman, J.; van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef]
- Wallace, H.M.; Fraser, A.V.; Hughes, A. A perspective of polyamine metabolism. Biochem. J. 2003, 376, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Childs, A.C.; Mehta, D.J.; Gerner, E.W. Polyamine-dependent gene expression. Cell. Mol. Life Sci. 2003, 60, 1394–1406. [Google Scholar] [CrossRef]
- Nitta, T.; Igarashi, K.; Yamamoto, N. Polyamine depletion induces apoptosis through mitochondria-mediated pathway. Exp. Cell Res. 2002, 276, 120–128. [Google Scholar] [CrossRef]
- Bachrach, U.; Wang, Y.C.; Tabib, A. Polyamines: New cues in cellular signal transduction. News Physiol. Sci. 2001, 16, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Miller-Fleming, L.; Olin-Sandoval, V.; Campbell, K.; Ralser, M. Remaining Mysteries of Molecular Biology: The Role of Polyamines in the Cell. J. Mol. Biol. 2015, 427, 3389–3406. [Google Scholar] [CrossRef]
- Levêque, J.; Foucher, F.; Bansard, J.Y.; Havouis, R.; Grall, J.Y.; Moulinoux, J.P. Polyamine profiles in tumor, normal tissue of the homologous breast, blood, and urine of breast cancer sufferers. Breast Cancer Res. Treat. 2000, 60, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Fabian, C.J.; Kimler, B.F.; Brady, D.A.; Mayo, M.S.; Chang, C.H.; Ferraro, J.A.; Zalles, C.M.; Stanton, A.L.; Masood, S.; Grizzle, W.E.; et al. A phase II breast cancer chemoprevention trial of oral alpha-difluoromethylornithine: Breast tissue, imaging, and serum and urine biomarkers. Clin. Cancer Res. 2002, 8, 3105–3117. [Google Scholar]
- O’Shaughnessy, J.A.; Demers, L.M.; Jones, S.E.; Arseneau, J.; Khandelwal, P.; George, T.; Gersh, R.; Mauger, D.; Manni, A. Alpha-difluoromethylornithine as treatment for metastatic breast cancer patients. Clin. Cancer Res. 1999, 5, 3438–3444. [Google Scholar]
- Horn, Y.; Schechter, P.J.; Marton, L.J. Phase I-II clinical trial with alpha-difluoromethylornithine--an inhibitor of polyamine biosynthesis. Eur. J. Cancer Clin. Oncol. 1987, 23, 1103–1107. [Google Scholar] [CrossRef]
- Hayes, C.S.; Shicora, A.C.; Keough, M.P.; Snook, A.E.; Burns, M.R.; Gilmour, S.K. Polyamine-blocking therapy reverses immunosuppression in the tumor microenvironment. Cancer Immunol. Res. 2014, 2, 274–285. [Google Scholar] [CrossRef] [Green Version]
- Kremmer, T.; Palyi, I.; Daubner, D.; Boldizsar, M.; Vincze, B.; Paulik, E.; Sugar, J.; Pokorny, E.; Tury, E. Comparative Studies on the Polyamine Metabolism and DFMO Treatment of MCF-7 and MDA-MB-231 Breast Cancer Cell Lines and Xenografts. Anticancer Res. 1991, 11, 1807–1813. [Google Scholar]
- Hoggard, C.D. Polyamines and growth regulation of cultured human breast cancer cells by 17 beta-oestradiol. Mol. Cell. Endocrinol. 1986, 46, 71–78. [Google Scholar] [CrossRef]
- Gartel, A.L.; Feliciano, C.; Tyner, A.L. A new method for determining the status of p53 in tumor cell lines of different origin. Oncol. Res. 2003, 13, 405–408. [Google Scholar] [CrossRef]
- Rodrik, V.; Gomes, E.; Hu, L.; Rockwell, P.; Foster, D.A. Myc stabilization in response to estrogen and phospholipase D in MCF-7 breast cancer cells. FEBS Lett. 2006, 580, 5647–5652. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Farrell, A.S.; Daniel, C.J.; Arnold, H.; Scanlan, C.; Laraway, B.J.; Janghorban, M.; Lum, L.; Chen, D.; Troxell, M.; et al. Mechanistic insight into Myc stabilization in breast cancer involving aberrant Axin1 expression. Proc. Natl. Acad. Sci. USA 2012, 109, 2790–2795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachmann, A.S.; Geerts, D. Polyamine synthesis as a target of MYC oncogenes. J. Biol. Chem. 2018, 293, 18757–18769. [Google Scholar] [CrossRef] [Green Version]
- Bello-Fernandez, C.; Packham, G.; Cleveland, J.L. The ornithine decarboxylase gene is a transcriptional target of c-Myc. Proc. Natl. Acad. Sci. USA 1993, 90, 7804–7808. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Ye, Q.; Deng, P.; Cao, Y.; He, D.; Zhou, Z.; Wang, C.; Zaytseva, Y.Y.; Schwartz, C.E.; Lee, E.Y.; et al. Spermine synthase and MYC cooperate to maintain colorectal cancer cell survival by repressing Bim expression. Nat. Commun. 2020, 11, 3243. [Google Scholar] [CrossRef] [PubMed]
- Dever, T.E.; Gutierrez, E.; Shin, B.S. The hypusine-containing translation factor eIF5A. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, E.; Shin, B.S.; Woolstenhulme, C.J.; Kim, J.R.; Saini, P.; Buskirk, A.R.; Dever, T.E. eIF5A promotes translation of polyproline motifs. Mol. Cell 2013, 51, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Pegg, A.E. Functions of Polyamines in Mammals. J. Biol. Chem. 2016, 291, 14904–14912. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DFMO Exposure Time (h) | Treatment | MDA-MB-231 | MCF-7 | ||||
|---|---|---|---|---|---|---|---|
| Total PA (nmol/mg Protein) | PA Decrease (% of Untreated) | Growth Inhibition (% of Untreated) | Total PA (nmol/mg Protein) | PA Decrease (% of Untreated) | Growth Inhibition (% of Untreated) | ||
| 0 | Untreated | 20.7 ± 1.7 | 0 | 0 | 22.7 ± 1.4 | 0 | 0 |
| 48 | Untreated DFMO | 18.8 ± 1.3 7.5 ± 0.5 * | 60 | 13 | 23.3 ± 1.1 9.5 ± 0.4 * | 59 | 12 |
| 96 | Untreated DFMO | 11.6 ± 0.8 5.2 ± 0.1 * | 55 | 42 | 15.1 ± 0.6 7.8 ± 0.4 * | 48 | 50 |
| 120 | Untreated DFMO | 11.3 ± 1.3 6.2 ± 0.2 * | 45 | 50 | 14.4 ± 0.9 6.6 ± 0.4 * | 54 | 70 |
| 144 | Untreated DFMO | 7.0 ± 1.2 4.3 ± 0.3 * | 39 | 43 | nd | nd | nd |
| Time in Culture (h) | DFMO Exposure Time (h) | DFMO Conc. (5 mM) | Intracellular DFMO (nmol/mg protein) |
|---|---|---|---|
| 72 | 48 | 48 | 9.9 ± 2.5 |
| 120 | 96 | Continuous 48 | 15.5 ± 2.5 3.3 ± 0.8 |
| 168 | 144 | Continuous 48 | 18.6 ± 1.3 1.4 ± 0.3 |
| 216 | 192 | Continuous 48 | 16.8 ± 1.6 0.6 ± 0.1 |
| 264 | 240 | Continuous 48 | 20.1 ± 1.0 0.3 ± 0.0 |
| 312 | 288 | Continuous 48 | 26.7 ± 6.3 0.1 ± 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akinyele, O.; Wallace, H.M. Characterising the Response of Human Breast Cancer Cells to Polyamine Modulation. Biomolecules 2021, 11, 743. https://doi.org/10.3390/biom11050743
Akinyele O, Wallace HM. Characterising the Response of Human Breast Cancer Cells to Polyamine Modulation. Biomolecules. 2021; 11(5):743. https://doi.org/10.3390/biom11050743
Chicago/Turabian StyleAkinyele, Oluwaseun, and Heather M. Wallace. 2021. "Characterising the Response of Human Breast Cancer Cells to Polyamine Modulation" Biomolecules 11, no. 5: 743. https://doi.org/10.3390/biom11050743
APA StyleAkinyele, O., & Wallace, H. M. (2021). Characterising the Response of Human Breast Cancer Cells to Polyamine Modulation. Biomolecules, 11(5), 743. https://doi.org/10.3390/biom11050743