
Composition of Transcription Machinery and Its Crosstalk with Nucleoid-Associated Proteins and Global Transcription Factors

Abstract
:1. Introduction
2. The Intracellular Context
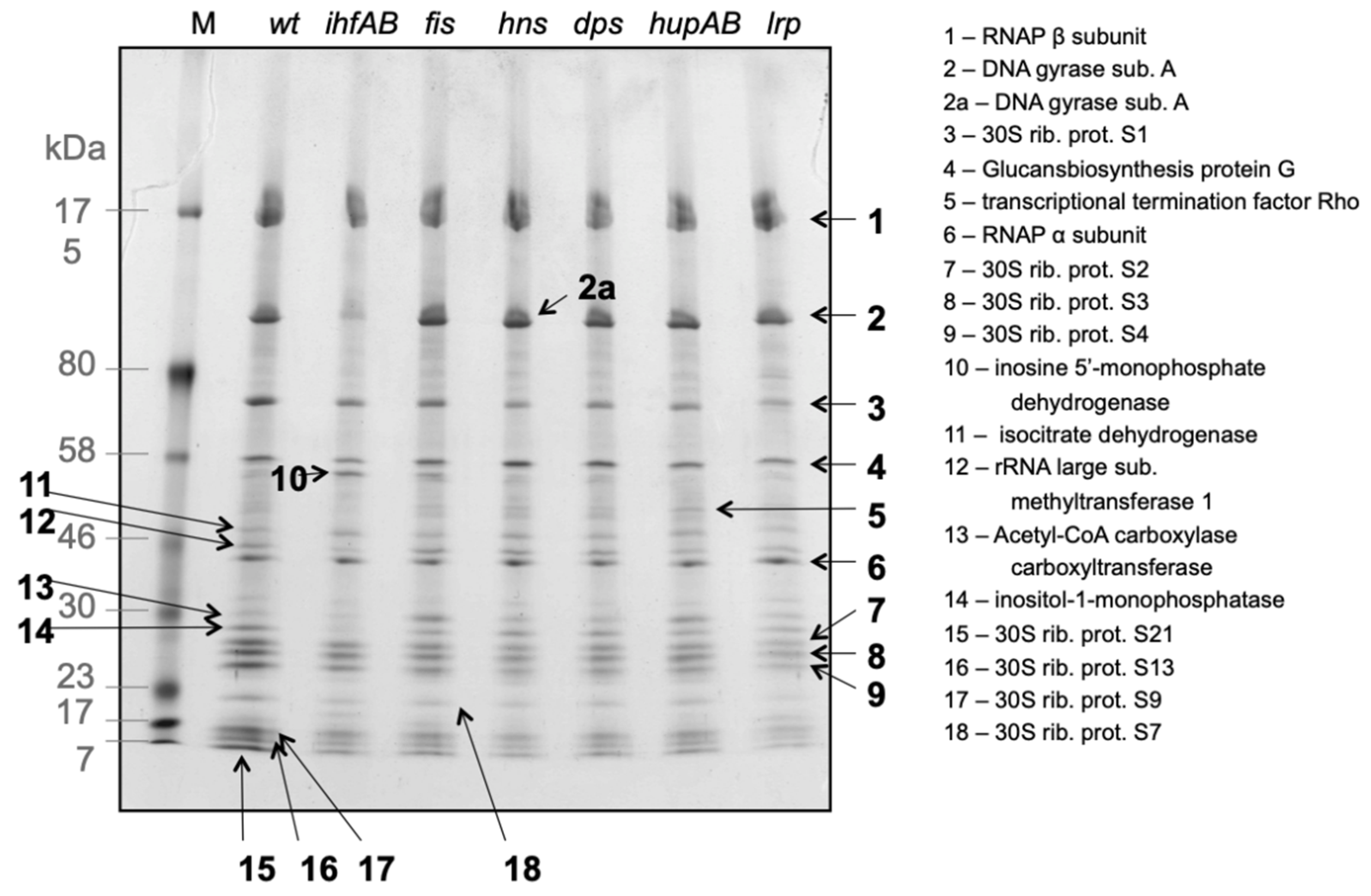
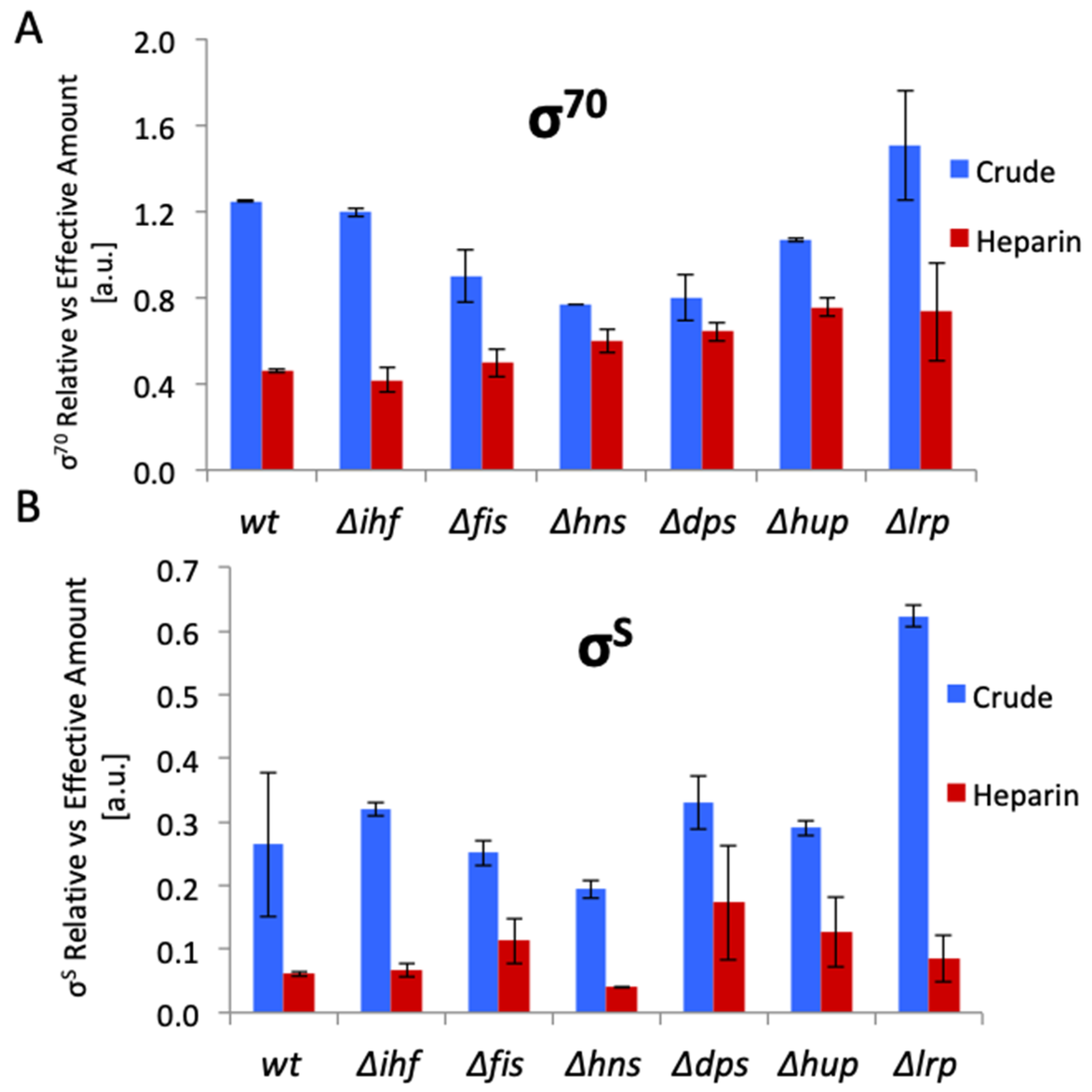
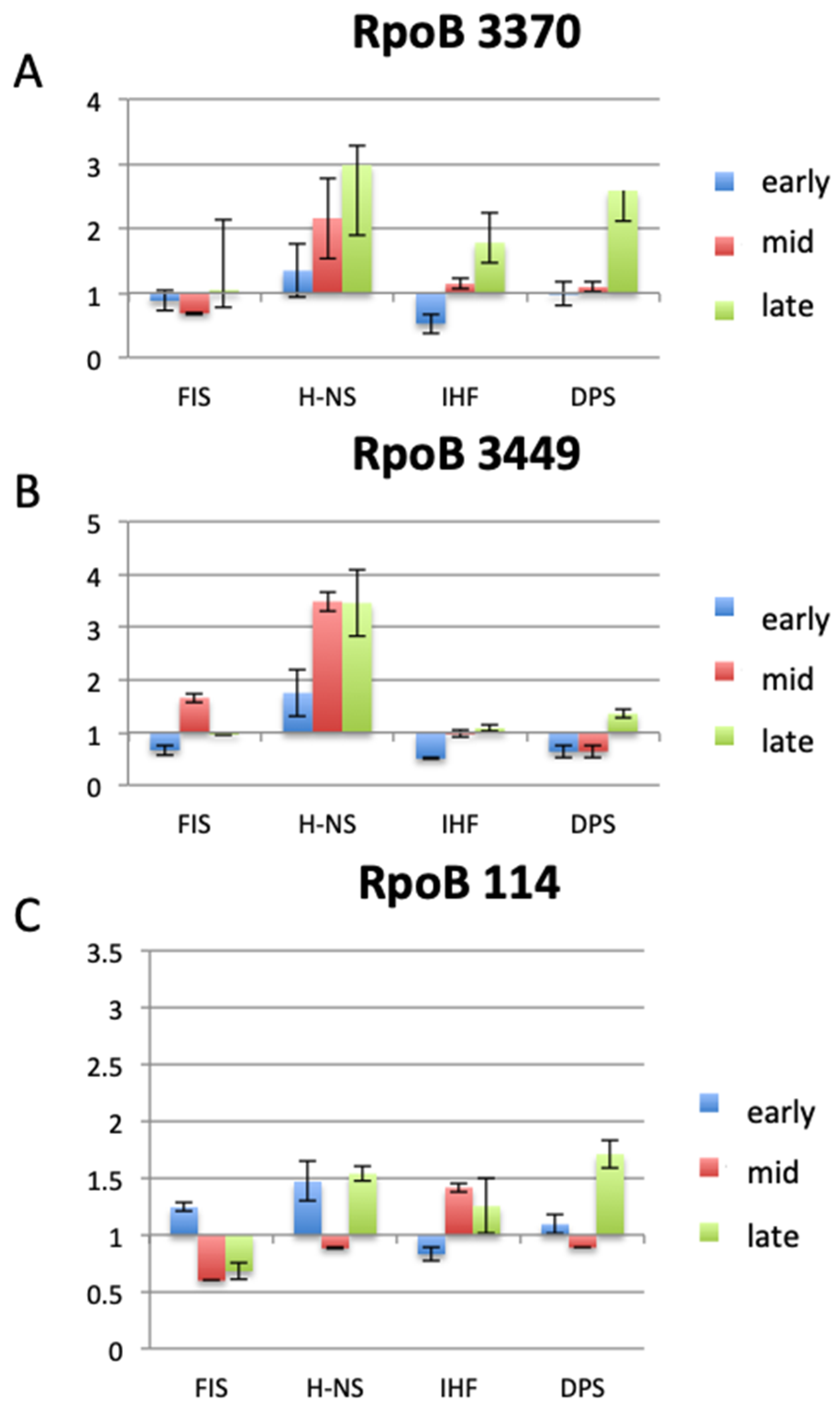
3. Modulators of RNAP
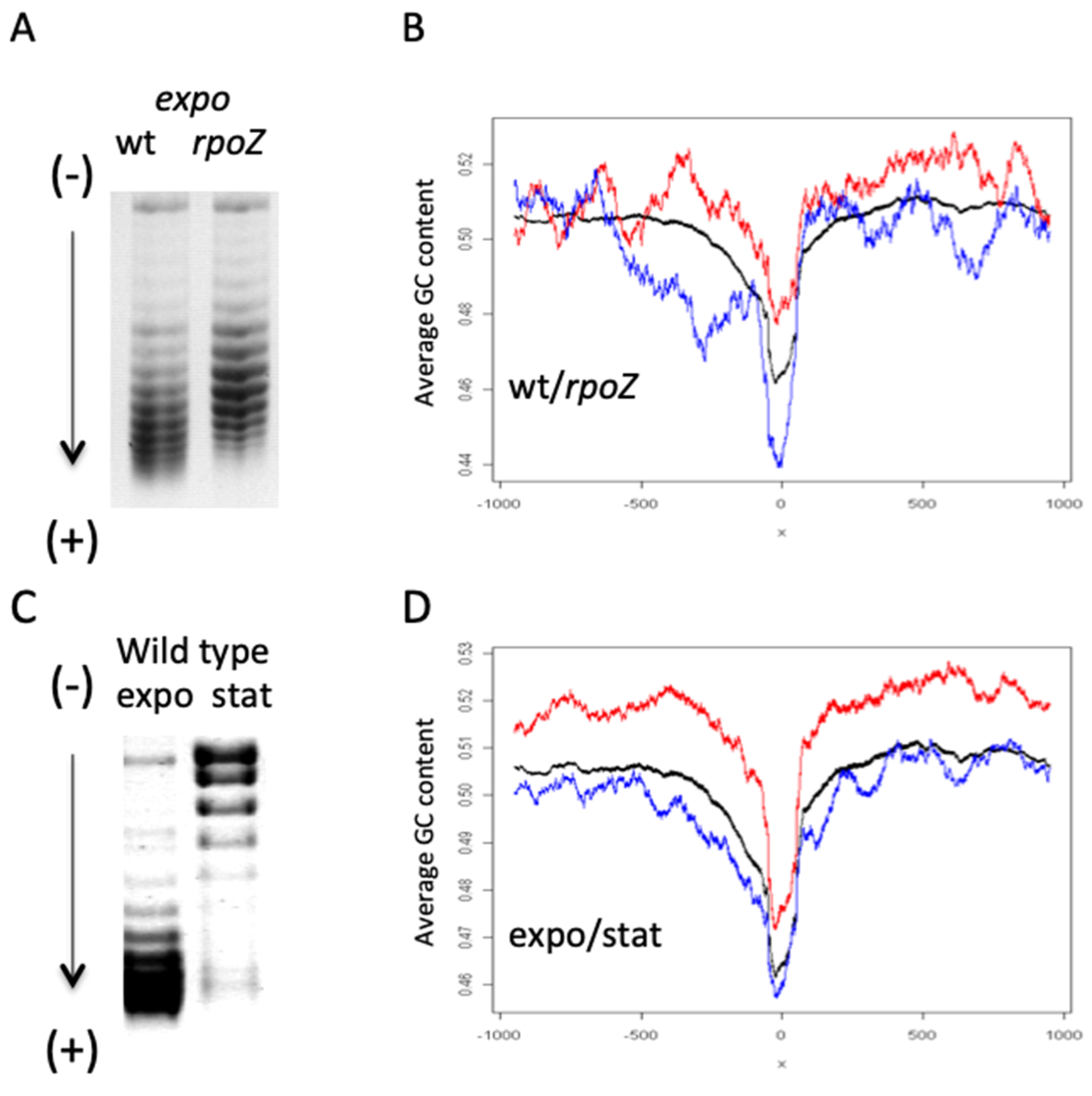
4. Role of DNA Topology and Homeostatic Regulation of Supercoiling Response
5. Interdependence of the Network Elements
6. Spatiotemporal Organization of Transcription in Genome
7. Regulons and Couplons
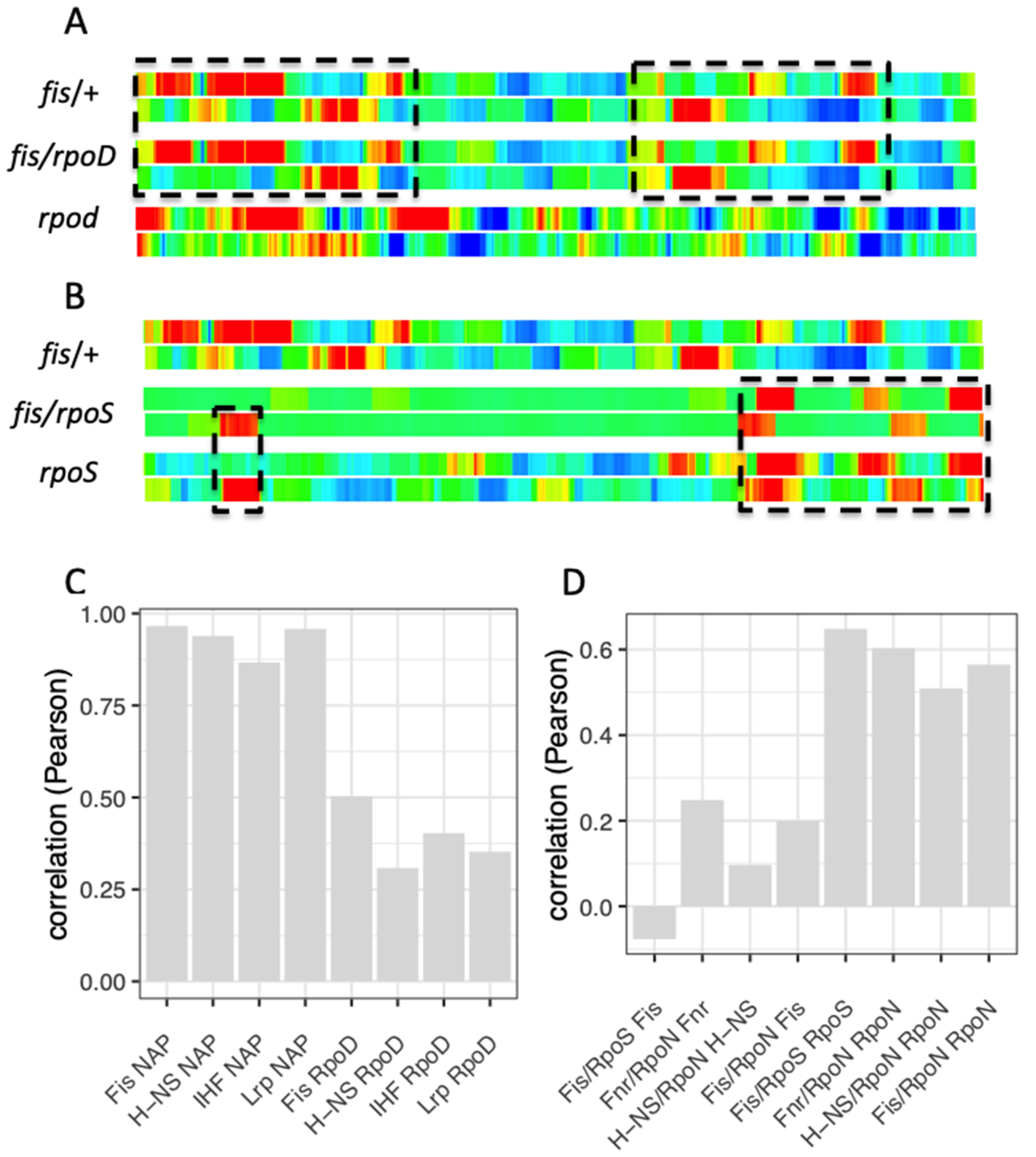
8. Spatial Organization of Regulons and Couplons
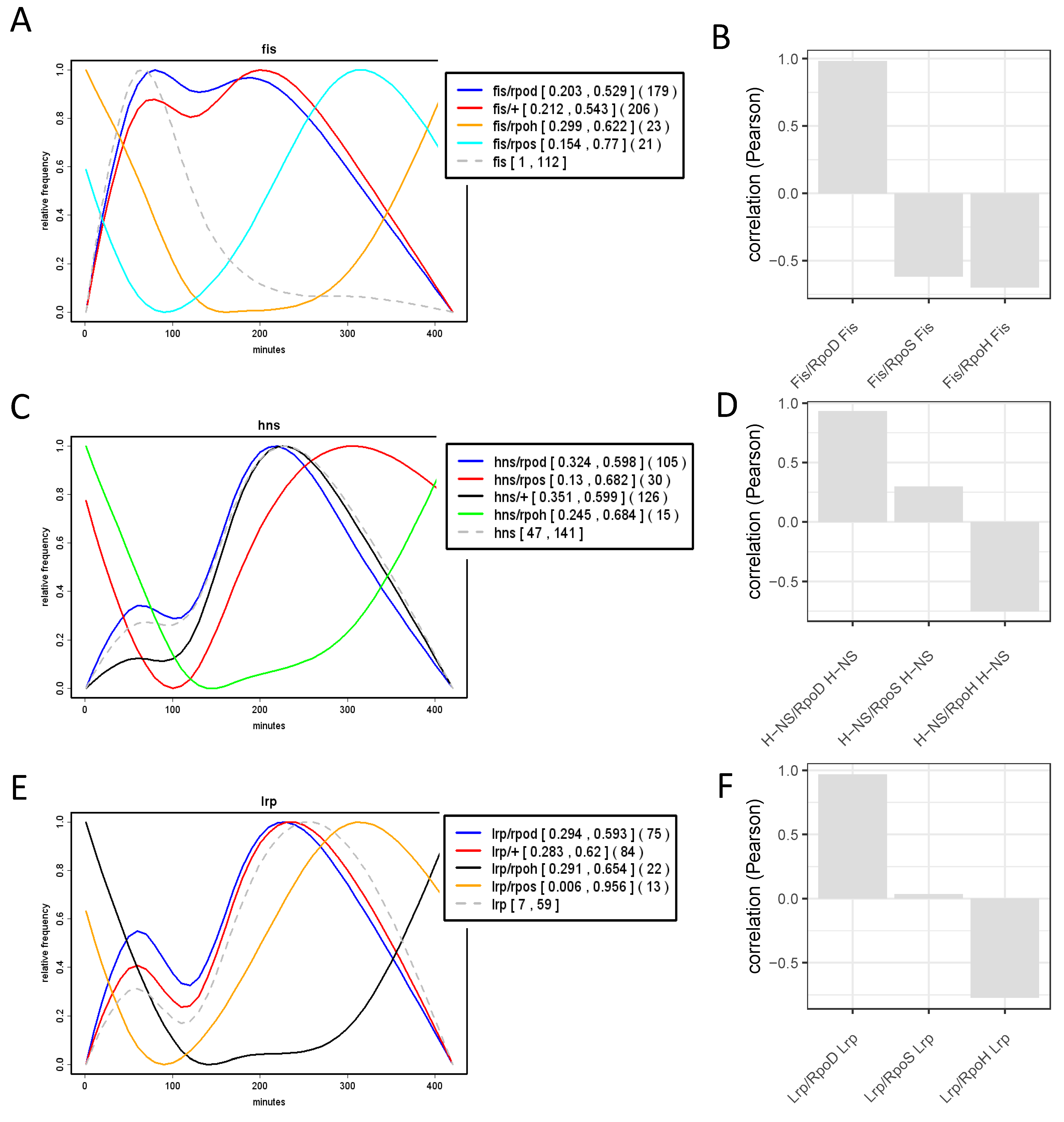
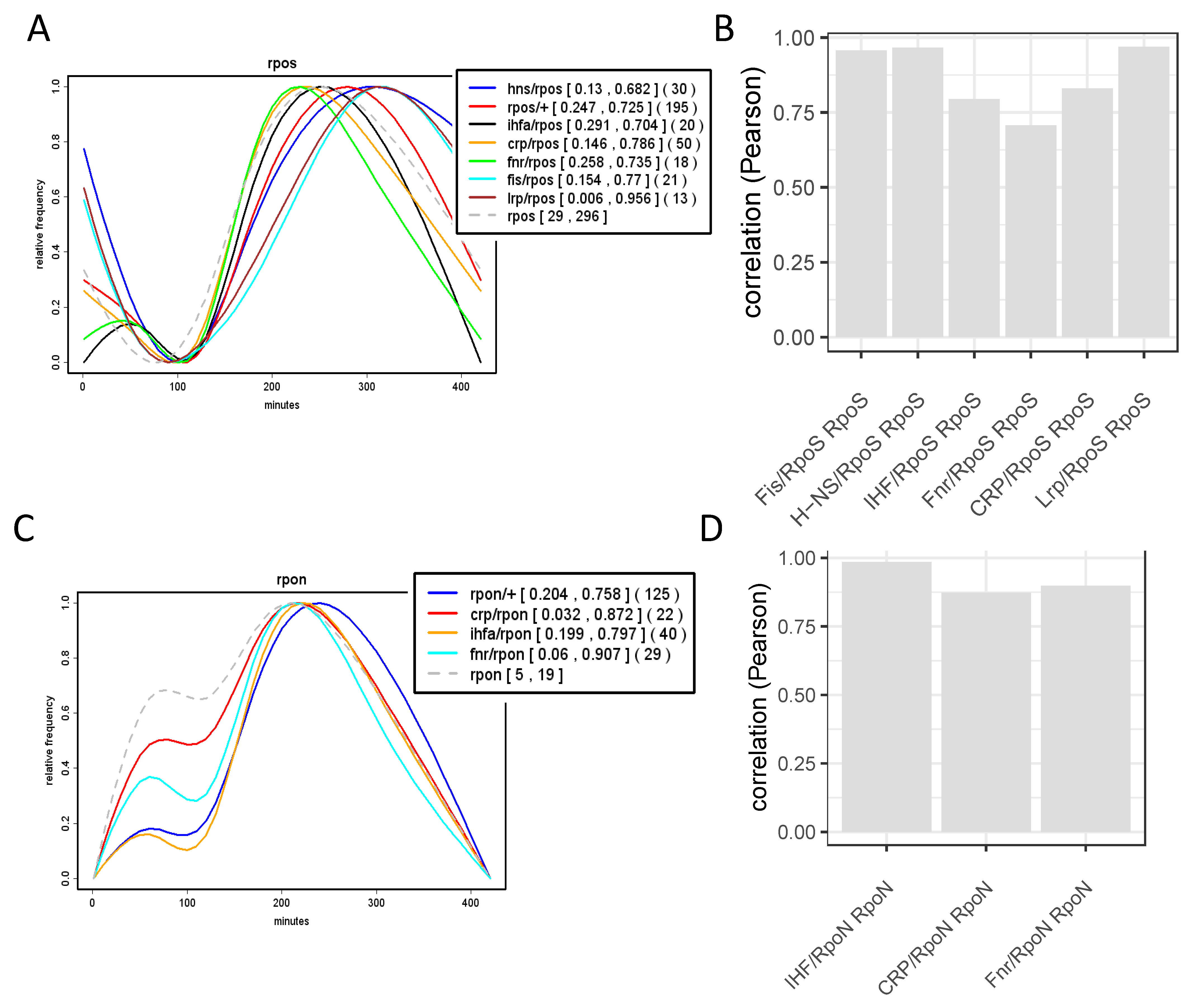
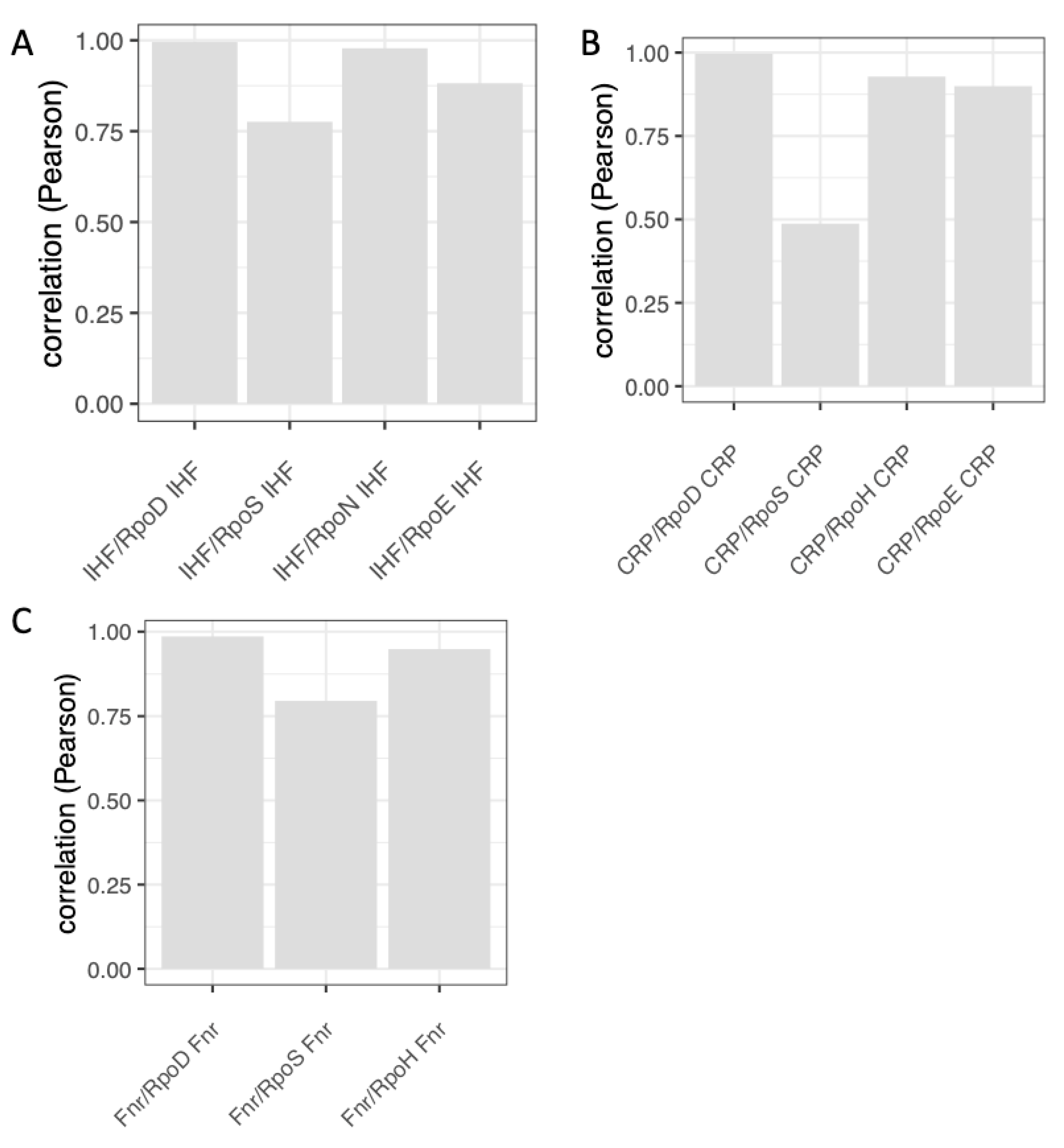
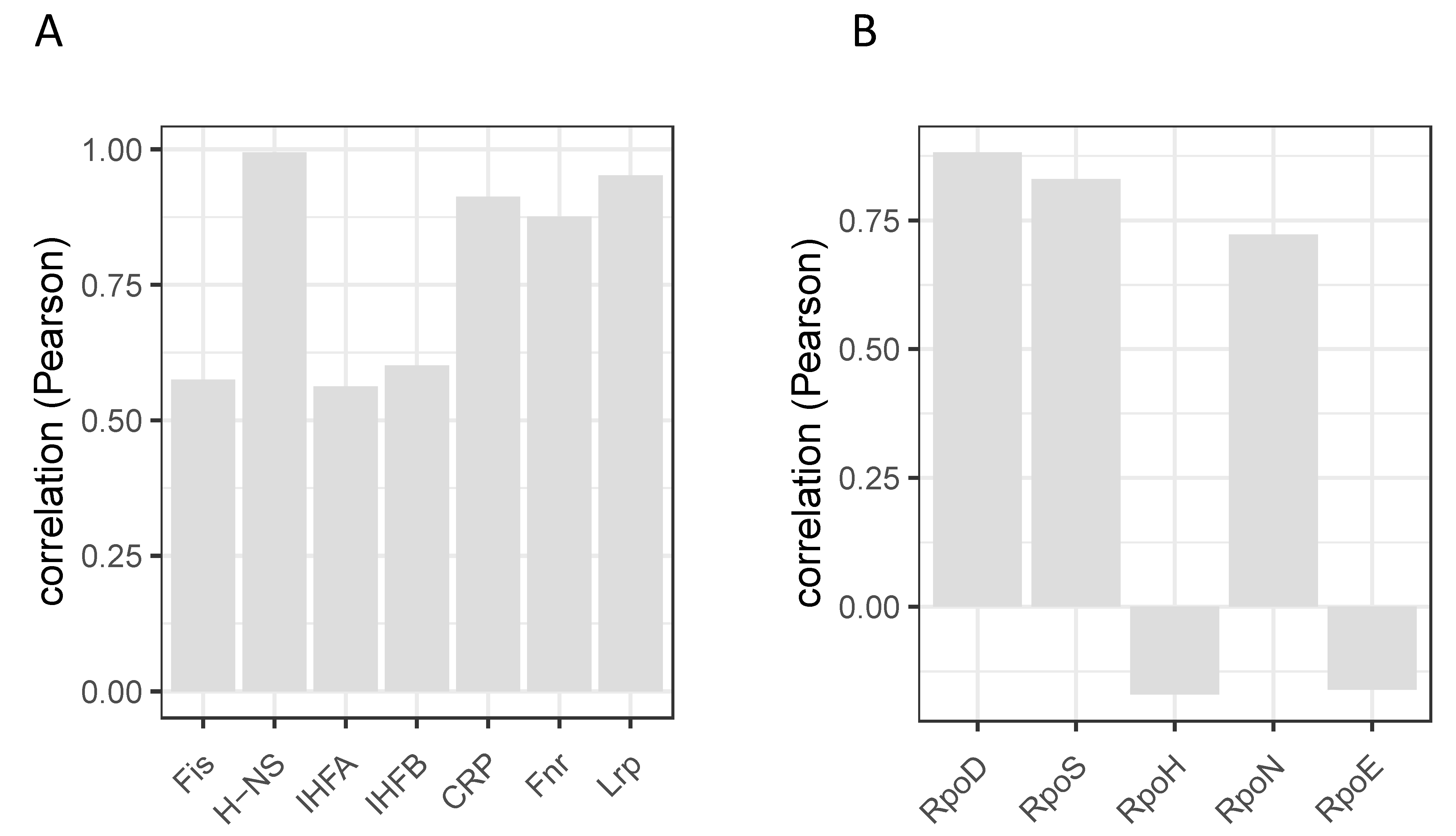
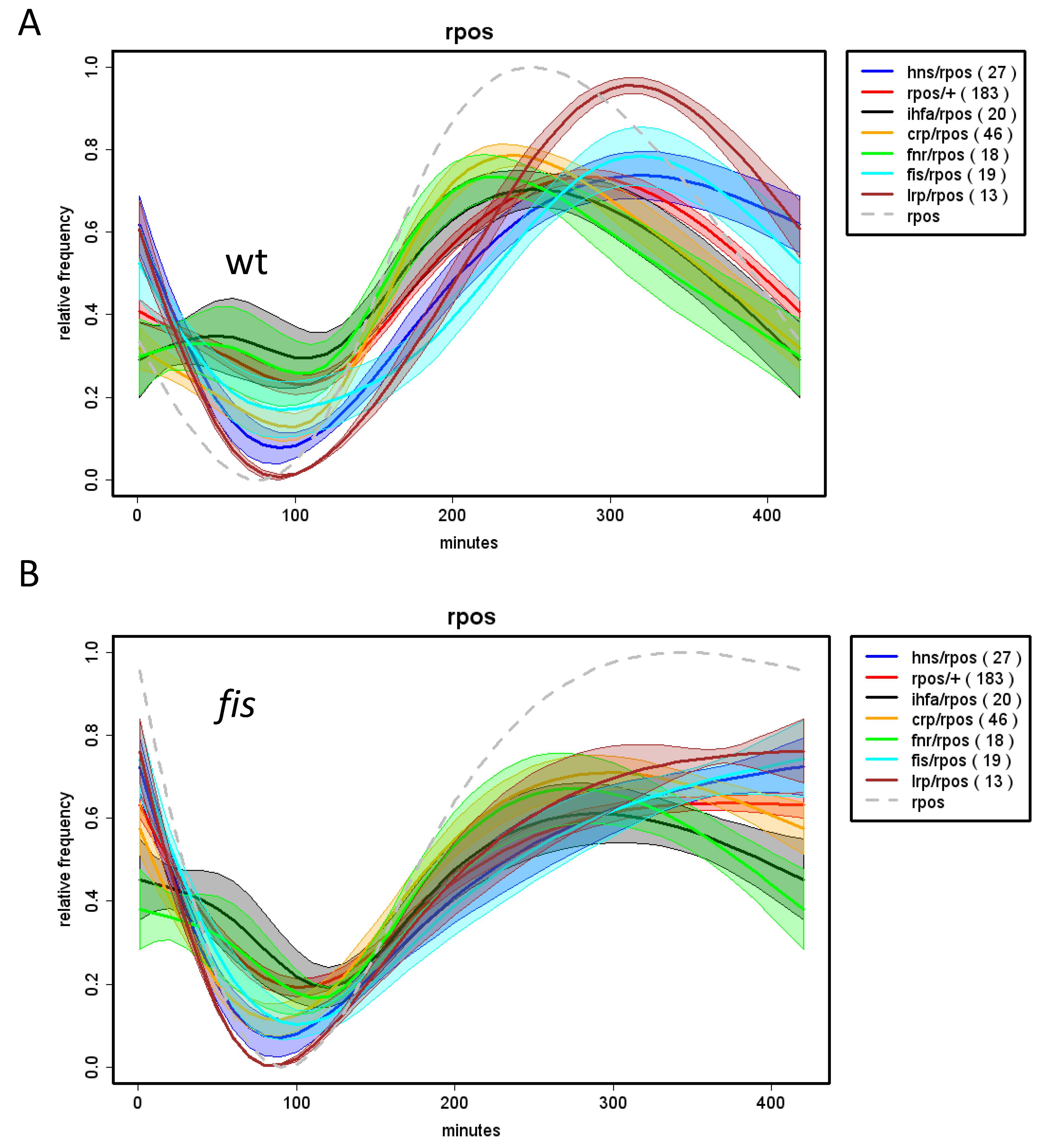
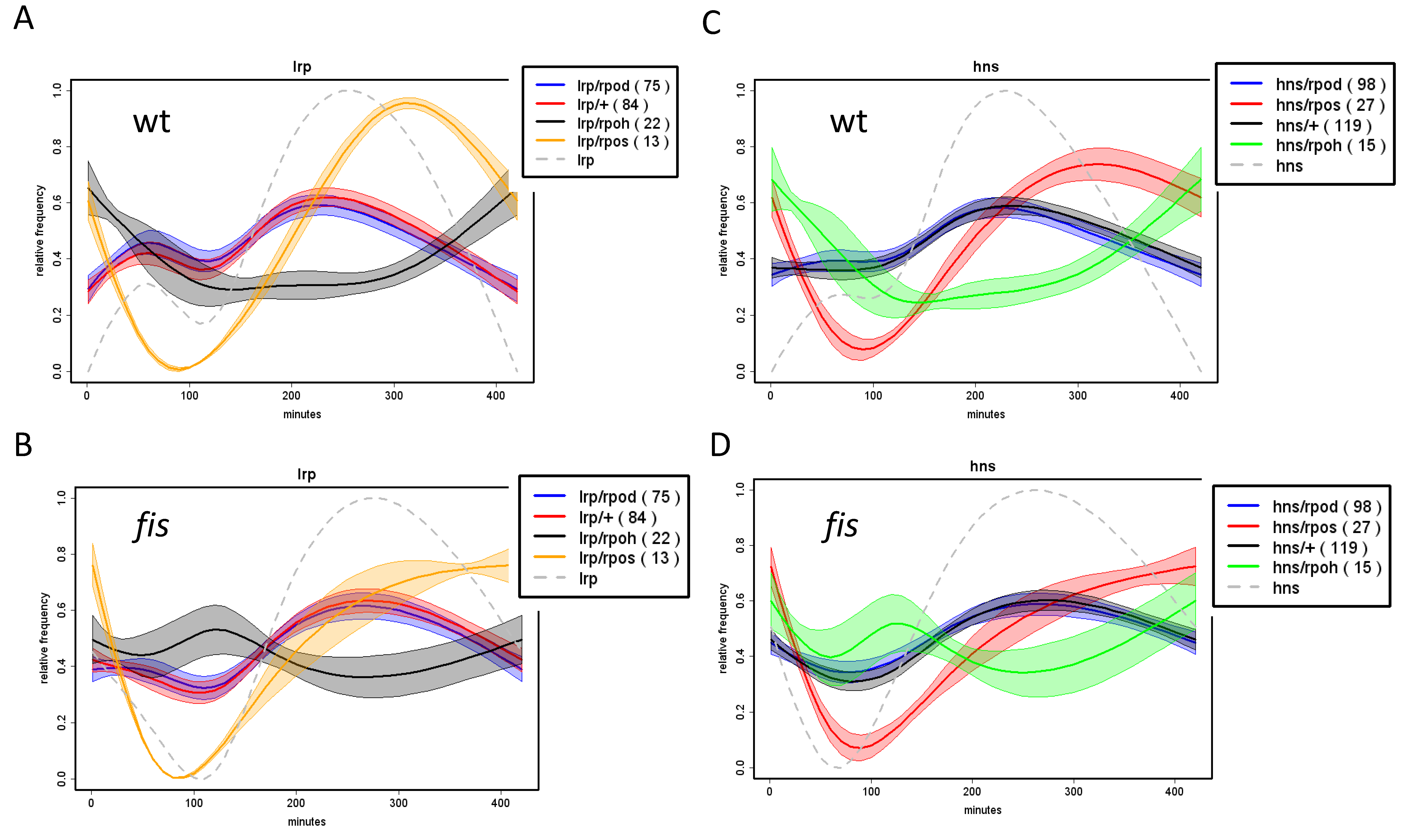
9. Temporal Expression Patterns of Regulons and Couplons
10. NAPs versus Global TFs?
11. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Travers, A.; Muskhelishvili, G. DNA supercoiling—A global transcriptional regulator for enterobacterial growth? Nat. Rev. Microbiol. 2005, 3, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Johansson, J.; Balsalobre, C.; Wang, S.-Y.; Urbonaviciene, J.; Jin, D.J.; Sondén, B.; Uhlin, B.E.; Johansson, J.; Balsalobre, C.; Wang, S.-Y.; et al. Nucleoid proteins stimulate stringently controlled bacterial promoters: A link between the cAMP-CRP and the (p)ppGpp regulons in Escherichia coli. Cell 2000, 102, 475–485. [Google Scholar] [CrossRef] [Green Version]
- Muskhelishvili, G.; Sobetzko, P.; Geertz, M.; Berger, M. General organisational principles of the transcriptional regulation system: A tree or a circle? Mol. BioSyst. 2010, 6, 662–676. [Google Scholar] [CrossRef]
- Cho, B.-K.; Kim, D.; Knight, E.M.; Zengler, K.; Palsson, B.O. Genome-scale reconstruction of the sigma factor network in Escherichia coli: Topology and functional states. BMC Biol. 2014, 12, 4. [Google Scholar] [CrossRef] [Green Version]
- Marr, C.; Geertz, M.; Hütt, M.-T.; Muskhelishvili, G. Dissecting the logical types of network control in gene expression profiles. BMC Syst. Biol. 2008, 2, 18. [Google Scholar] [CrossRef] [Green Version]
- Muskhelishvili, G.; Travers, A. Integration of syntactic and semantic properties of the DNA code reveals chromosomes as thermodynamic machines converting energy into information. Cell. Mol. Life Sci. 2013, 70, 4555–4567. [Google Scholar] [CrossRef]
- Beber, M.E.; Sobetzko, P.; Muskhelishvili, G.; Hütt, M.-T. Interplay of digital and analog control in time-resolved gene expression profiles. EPJ Nonlinear Biomed. Phys. 2016, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Salgado, H.; Peralta-Gil, M.; Gama-Castro, S.; Santos-Zavaleta, A.; Muñiz-Rascado, L.; García-Sotelo, J.S.; Weiss, V.; Solano-Lira, H.; Martínez-Flores, I.; Medina-Rivera, A.; et al. RegulonDB v8.0: Omics data sets, evolutionary conservation, regulatory phrases, cross-validated gold standards and more. Nucleic Acids Res. 2012, 41, D203–D213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnenschein, N.; Hütt, M.-T.; Stoyan, H.; Stoyan, D. Ranges of control in the transcriptional regulation of Escherichia coli. BMC Syst. Biol. 2009, 3, 119. [Google Scholar] [CrossRef] [Green Version]
- Brinza, L.; Calevro, F.; Charles, H. Genomic analysis of the regulatory elements and links with intrinsic DNA structural properties in the shrunken genome of Buchnera. BMC Genom. 2013, 14, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junier, I.; Frémont, P.; Rivoire, O. Universal and idiosyncratic characteristic lengths in bacterial genomes. Phys. Biol. 2018, 15, 035001. [Google Scholar] [CrossRef] [Green Version]
- El Houdaigui, B.; Forquet, R.; Hindré, T.; Schneider, D.; Nasser, W.; Reverchon, S.; Meyer, S. Bacterial genome architecture shapes global transcriptional regulation by DNA supercoiling. Nucleic Acids Res. 2019, 47, 5648–5657. [Google Scholar] [CrossRef] [Green Version]
- Peter, B.J.; Arsuaga, J.; Breier, A.M.; Khodursky, A.B.; Brown, P.O.; Cozzarelli, N.R. Genomic transcriptional response to loss of chromosomal supercoiling in Escherichia coli. Genome Biol. 2004, 5, R87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, K.S.; Ahn, J.; Khodursky, A.B. Spatial patterns of transcriptional activity in the chromosome of Escherichia coli. Genome Biol. 2004, 5, R86. [Google Scholar] [CrossRef] [Green Version]
- Blot, N.; Mavathur, R.; Geertz, M.; Travers, A.; Muskhelishvili, G. Homeostatic regulation of supercoiling sensitivity coordinates transcription of the bacterial genome. EMBO Rep. 2006, 7, 710–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Sobetzko, P.; Nasser, W.; Reverchon, S.; Muskhelishvili, G. Chromosomal “Stress-Response” Domains Govern the Spatiotemporal Expression of the Bacterial Virulence Program. mBio 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Martis, B.S.; Forquet, R.; Reverchon, S.; Nasser, W.; Meyer, S. DNA Supercoiling: An Ancestral Regulator of Gene Expression in Pathogenic Bacteria? Comput. Struct. Biotechnol. J. 2019, 17, 1047–1055. [Google Scholar] [CrossRef]
- Hsieh, L.S.; Rouviere-Yaniv, J.; Drlica, K. Bacterial DNA supercoiling and [ATP]/[ADP] ratio: Changes associated with salt shock. J. Bacteriol. 1991, 173, 3914–3917. [Google Scholar] [CrossRef] [Green Version]
- McClellan, J.A.; Boublikova, P.; Palecek, E.; Lilley, D.M. Superhelical torsion in cellular DNA responds directly to environmental and genetic factors. Proc. Natl. Acad. Sci. USA 1990, 87, 8373–8377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Workum, M.; Dooren, S.J.M.; Oldenburg, N.; Molenaar, D.; Jensen, P.R.; Snoep, J.L.; Westerhoff, H. DNA supercoiling depends on the phosphorylation potential in Escherichia coli. Mol. Microbiol. 1996, 20, 351–360. [Google Scholar] [CrossRef]
- Sonnenschein, N.; Geertz, M.; Muskhelishvili, G.; Hütt, M.-T. Analog regulation of metabolic demand. BMC Syst. Biol. 2011, 5, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthoumieux, S.; De Jong, H.; Baptist, G.; Pinel, C.; Ranquet, C.; Ropers, D.; Geiselmann, J. Shared control of gene expression in bacteria by transcription factors and global physiology of the cell. Mol. Syst. Biol. 2013, 9, 634. [Google Scholar] [CrossRef]
- Dillon, S.C.; Dorman, C. Bacterial nucleoid-associated proteins, nucleoid structure and gene expression. Nat. Rev. Microbiol. 2010, 8, 185–195. [Google Scholar] [CrossRef]
- Luijsterburg, M.S.; White, M.; Van Driel, R.; Dame, R.T. The Major Architects of Chromatin: Architectural Proteins in Bacteria, Archaea and Eukaryotes. Crit. Rev. Biochem. Mol. Biol. 2008, 43, 393–418. [Google Scholar] [CrossRef] [PubMed]
- Rimsky, S.; Travers, A. Pervasive regulation of nucleoid structure and function by nucleoid-associated proteins. Curr. Opin. Microbiol. 2011, 14, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Schultz, S.G.; Epstein, W.; Solomon, A.K. Cation Transport in Escherichia coli. J. Gen. Physiol. 1963, 47, 329–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinnbier, U.; Limpinsel, E.; Schmid, R.; Bakker, E.P. Transient accumulation of potassium glutamate and its replacement by trehalose during adaptation of growing cells of Escherichia coli K-12 to elevated sodium chloride concentrations. Arch. Microbiol. 1988, 150, 348–357. [Google Scholar] [CrossRef]
- McLaggan, D.; Naprstek, J.; Buurman, E.; Epstein, W. Interdependence of K+ and glutamate accumulation during osmotic adaptation of Escherichia coli. J. Biol. Chem. 1994, 269, 1911–1917. [Google Scholar] [CrossRef]
- Sobetzko, P.; Glinkowska, M.; Muskhelishvili, G. GSE65244: Temporal Gene Expression in Escherichia coli. Gene Expression Omnibus. 2017. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE65244 (accessed on 20 June 2021).
- Sobetzko, P.; Glinkowska, M.; Travers, A.; Muskhelishvili, G. DNA thermodynamic stability and supercoil dynamics determine the gene expression program during the bacterial growth cycle. Mol. BioSyst. 2013, 9, 1643–1651. [Google Scholar] [CrossRef]
- Stock, C.; Hielkema, L.; Tascón, I.; Wunnicke, D.; Oostergetel, G.T.; Azkargorta, M.; Paulino, C.; Hänelt, I. Cryo-EM structures of KdpFABC suggest a K+ transport mechanism via two inter-subunit half-channels. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Cheng, X.; Guinn, E.J.; Buechel, E.; Wong, R.; Sengupta, R.; Shkel, I.A.; Record, M.T. Basis of Protein Stabilization by K Glutamate: Unfavorable Interactions with Carbon, Oxygen Groups. Biophys. J. 2016, 111, 1854–1865. [Google Scholar] [CrossRef] [Green Version]
- Blundell, T.; Barlow, D.; Borkakoti, N.; Thornton, J. Solvent-induced distortions and the curvature of α-helices. Nature 1983, 306, 281–283. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.; Bauer, W. Supercoiling in closed circular DNA: Dependence upon ion type and concentration. Biochemistry 1978, 17, 594–601. [Google Scholar] [CrossRef]
- Rybenkov, V.V.; Vologodskii, A.V.; Cozzarelli, N.R. The effect of ionic conditions on DNA helical repeat, effective diameter and free energy of supercoiling. Nucleic Acids Res. 1997, 25, 1412–1418. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Travers, A. Modelling and DNA topology of compact 2-start and 1-start chromatin fibres. Nucleic Acids Res. 2019, 47, 9902–9924. [Google Scholar] [CrossRef]
- Ohlsen, K.L.; Gralla, J.D. Melting during steady-state transcription of the rrnB P1 promoter in vivo and in vitro. J. Bacteriol. 1992, 174, 6071–6075. [Google Scholar] [CrossRef] [Green Version]
- Travers, A.; Muskhelishvili, G. Chromosomal Organization and Regulation of Genetic Function in Escherichia coli Integrates the DNA Analog and Digital Information. EcoSal Plus 2020, 9. [Google Scholar] [CrossRef]
- Geertz, M.; Travers, A.; Mehandziska, S.; Sobetzko, P.; Janga, S.C.; Shimamoto, N.; Muskhelishvili, G. Structural coupling between RNA polymerase composition and DNA supercoiling in coordinating transcription: A global role for the omega subunit? mBio 2011, 2, e00034-11. [Google Scholar] [CrossRef] [Green Version]
- Vinella, D.; Potrykus, K.; Murphy, H.; Cashel, M. Effects on growth by changes of the balance between GreA, GreB, and DksA suggest mutual competition and functional redundancy in Escherichia coli. J. Bacteriol. 2012, 194, 261–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, B.; Zhu, C.-X.; Ji, C.; Ahumada, A.; Tse-Dinh, Y.-C. Direct interaction between Escherichia coli RNA polymerase and the zinc ribbon domains of DNA topoisomerase I. J. Biol. Chem. 2003, 278, 30705–30710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.; China, A.; Manjunatha, U.H.; Ponnanna, N.; Nagaraja, V. A complex of DNA gyrase and RNA polymerase fosters transcription in Mycobacterium smegmatis. Biochem. Biophys. Res. Commun. 2006, 343, 1141–1145. [Google Scholar] [CrossRef]
- Verma, S.; Xiong, Y.; Mayer, M.U.; Squier, T.C. Remodeling of the bacterial RNA polymerase supramolecular complex in response to environmental conditions. Biochemistry 2007, 46, 3023–3035. [Google Scholar] [CrossRef]
- Banda, S.; Cao, N.; Tse-Dinh, Y.-C. Distinct Mechanism Evolved for Mycobacterial RNA Polymerase and Topoisomerase I Protein-Protein Interaction. J. Mol. Biol. 2017, 429, 2931–2942. [Google Scholar] [CrossRef]
- Mehandziska, S.; Petrescu, A.M.; Muskhelishvili, G. Isolation and Analysis of RNA Polymerase Supramolecular Complex with Associated Proteins. Methods Mol. Biol. 2017, 1624, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Eason, I.R.; Kaur, H.P.; Alexander, K.A.; Sukhodolets, M.V. Growth phase-specific changes in the composition of E. coli transcription complexes. J. Chromatogr. B 2019, 1109, 155–165. [Google Scholar] [CrossRef]
- Kohler, R.; Mooney, R.A.; Mills, D.J.; Landick, R.; Cramer, P. Architecture of a transcribing-translating expressome. Science 2017, 356, 194–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, G.E.; Lalanne, J.-B.; Peters, M.L.; Li, G.-W. Functionally uncoupled transcription–translation in Bacillus subtilis. Nature 2020, 585, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Bubunenko, M.; Smith, C.; Abbott, D.M.; Stringer, A.M.; Shi, R.; Court, D.L.; Wade, J.T. SuhB Associates with Nus Factors to Facilitate 30S Ribosome Biogenesis in Escherichia coli. mBio 2016, 7, e00114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudenhoeffer, B.R.; Schneider, H.; Schweimer, K.; Knauer, S.H. SuhB is an integral part of the ribosomal antitermination complex and interacts with NusA. Nucleic Acids Res. 2019, 47, 6504–6518. [Google Scholar] [CrossRef]
- Hedstrom, L. IMP Dehydrogenase: Structure, mechanism, and inhibition. Chem. Rev. 2009, 109, 2903–2928. [Google Scholar] [CrossRef] [Green Version]
- McLean, J.E.; Hamaguchi, N.; Belenky, P.; Mortimer, S.E.; Stanton, M.; Hedstrom, L. Inosine 5′-monophosphate dehydrogenase binds nucleic acids in vitro and in vivo. Biochem. J. 2004, 379, 243–251. [Google Scholar] [CrossRef]
- Kozhevnikova, E.N.; van der Knaap, J.A.; Pindyurin, A.V.; Ozgur, Z.; van Ijcken, W.; Moshkin, Y.M.; Verrijzer, C.P. Metabolic enzyme IMPDH is also a transcription factor regulated by cellular state. Mol. Cell 2012, 47, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Pimkin, M.; Pimkina, J.; Markham, G.D. A regulatory role of the Bateman domain of IMP dehydrogenase in adenylate nucleotide biosynthesis. J. Biol. Chem. 2009, 284, 7960–7969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snoep, J.L.; van der Weijden, C.C.; Andersen, H.W.; Westerhoff, H.V.; Jensen, P.R. DNA supercoiling in Escherichia coli is under tight and subtle homeostatic control, involving gene-expression and metabolic regulation of both topoisomerase I and DNA gyrase. Eur. J. Biochem. 2002, 269, 1662–1669. [Google Scholar] [CrossRef] [PubMed]
- Riles, L.; Shaw, R.J.; Johnston, M.; Reines, D. Large-scale screening of yeast mutants for sensitivity to the IMP dehydrogenase inhibitor 6-azauracil. Yeast 2004, 21, 241–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurkiewicz, A.; Leśniewska, E.; Cieśla, M.; Gorjão, N.; Kantidakis, T.; White, R.J.; Boguta, M.; Graczyk, D. Inhibition of tRNA Gene Transcription by the Immunosuppressant Mycophenolic Acid. Mol. Cell. Biol. 2019, 40, e00294-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortimer, S.E.; Xu, D.; McGrew, D.; Hamaguchi, N.; Lim, H.C.; Bowne, S.J.; Daiger, S.P.; Hedstrom, L. IMP dehydrogenase type 1 associates with polyribosomes translating rhodopsin mRNA. J. Biol. Chem. 2008, 283, 36354–36360. [Google Scholar] [CrossRef] [Green Version]
- Böhringer, J.; Fischer, D.; Mosler, G.; Hengge-Aronis, R. UDP-glucose is a potential intracellular signal molecule in the control of expression of sigma S and sigma S-dependent genes in Escherichia coli. J. Bacteriol. 1995, 177, 413–422. [Google Scholar] [CrossRef] [Green Version]
- Krause, K.; Maciąg-Dorszyńska, M.; Wosinski, A.; Gaffke, L.; Morcinek-Orłowska, J.; Rintz, E.; Bielańska, P.; Szalewska-Pałasz, A.; Muskhelishvili, G.; Węgrzyn, G. The Role of Metabolites in the Link between DNA Replication and Central Carbon Metabolism in Escherichia coli. Genes 2020, 11, 447. [Google Scholar] [CrossRef] [Green Version]
- Berger, M.; Farcas, A.; Geertz, M.; Zhelyazkova, P.; Brix, K.; Travers, A.; Muskhelishvili, G. Coordination of genomic structure and transcription by the main bacterial nucleoid-associated protein HU. EMBO Rep. 2009, 11, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Weng, X.; Bohrer, C.H.; Bettridge, K.; Lagda, A.C.; Cagliero, C.; Jin, D.J.; Xiao, J. Spatial organization of RNA polymerase and its relationship with transcription in Escherichia coli. Proc. Natl. Acad. Sci. USA 2019, 116, 20115–20123. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.F.; Wang, J.C. Supercoiling of the DNA template during transcription. Proc. Natl. Acad. Sci. USA 1987, 84, 7024–7027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutormin, D.; Rubanova, N.; Logacheva, M.; Ghilarov, D.; Severinov, K. Single-nucleotide-resolution mapping of DNA gyrase cleavage sites across the Escherichia coli genome. Nucleic Acids Res. 2018, 47, 1373–1388. [Google Scholar] [CrossRef] [Green Version]
- Muskhelishvili, G.; Travers, A. Transcription factor as a topological homeostat. Front. Biosci. 2003, 8, d279–285. [Google Scholar] [CrossRef] [Green Version]
- Maurer, S.; Fritz, J.; Muskhelishvili, G.; Travers, A. RNA polymerase and an activator form discrete subcomplexes in a transcription initiation complex. EMBO J. 2006, 25, 3784–3790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muskhelishvili, G.; Travers, A. The regulatory role of DNA supercoiling in nucleoprotein complex assembly and genetic activity. Biophys. Rev. 2016, 8 (Suppl. 1), 5–22. [Google Scholar] [CrossRef] [Green Version]
- Menzel, R.; Gellert, M. Regulation of the genes for E. coli DNA gyrase: Homeostatic control of DNA supercoiling. Cell 1983, 34, 105–113. [Google Scholar] [CrossRef]
- Gaal, T.; Mandel, M.J.; Silhavy, T.J.; Gourse, R.L.; Gaal, T.; Mandel, M.J.; Silhavy, T.J.; Gourse, R.L. Crl facilitates RNA polymerase holoenzyme formation. J. Bacteriol. 2006, 188, 7966–7970. [Google Scholar] [CrossRef] [Green Version]
- Typas, A.; Barembruch, C.; Possling, A.; Hengge, R. Stationary phase reorganisation of the Escherichia coli transcription machinery by Crl protein, a fine-tuner of σs activity and levels. EMBO J. 2007, 26, 1569–1578. [Google Scholar] [CrossRef]
- Banta, A.; Chumanov, R.S.; Yuan, A.H.; Lin, H.; Campbell, E.; Burgess, R.R.; Gourse, R.L. Key features of S required for specific recognition by Crl, a transcription factor promoting assembly of RNA polymerase holoenzyme. Proc. Natl. Acad. Sci. USA 2013, 110, 15955–15960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minakhin, L.; Bhagat, S.; Brunning, A.; Campbell, E.; Darst, S.A.; Ebright, R.; Severinov, K. Bacterial RNA polymerase subunit omega and eukaryotic RNA polymerase subunit RPB6 are sequence, structural, and functional homologs and promote RNA polymerase assembly. Proc. Natl. Acad. Sci. USA 2001, 98, 892–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artsimovitch, I.; Landick, R. Pausing by bacterial RNA polymerase is mediated by mechanistically distinct classes of signals. Proc. Natl. Acad. Sci. USA 2000, 97, 7090–7095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burmann, B.M.; Schweimer, K.; Luo, X.; Wahl, M.; Stitt, B.L.; Gottesman, M.E.; Rösch, P.; Burmann, B.M.; Schweimer, K.; Luo, X.; et al. A NusE:NusG complex links transcription and translation. Science 2010, 328, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Webster, M.W.; Takacs, M.; Zhu, C.; Vidmar, V.; Eduljee, A.; Abdelkareem, M.; Weixlbaumer, A. Structural basis of transcription-translation coupling and collision in bacteria. Science 2020, 369, 1355–1359. [Google Scholar] [CrossRef]
- Wang, C.; Molodtsov, V.; Firlar, E.; Kaelber, J.T.; Blaha, G.; Su, M.; Ebright, R.H. Structural basis of transcription-translation coupling. Science 2020, 369, 1359–1365. [Google Scholar] [CrossRef] [PubMed]
- Washburn, R.S.; Zuber, P.K.; Sun, M.; Hashem, Y.; Shen, B.; Li, W.; Harvey, S.; Reyes, F.J.A.; Gottesman, M.E.; Knauer, S.H.; et al. Escherichia coli NusG Links the Lead Ribosome with the Transcription Elongation Complex. iScience 2020, 23, 101352. [Google Scholar] [CrossRef]
- Dutta, D.; Shatalin, K.; Epshtein, V.; Gottesman, M.E.; Nudler, E. Linking RNA polymerase backtracking to genome instability in E. coli. Cell 2011, 146, 533–543. [Google Scholar] [CrossRef] [Green Version]
- Cardinale, C.J.; Washburn, R.S.; Tadigotla, V.R.; Brown, L.M.; Gottesman, M.E.; Nudler, E. Termination factor Rho and its cofactors NusA and NusG silence foreign DNA in E. coli. Science 2008, 320, 935–938. [Google Scholar] [CrossRef] [Green Version]
- Peters, J.M.; Mooney, R.A.; Kuan, P.F.; Rowland, J.L.; Keleş, S.; Landick, R. Rho directs widespread termination of intragenic and stable RNA transcription. Proc. Natl. Acad. Sci. USA 2009, 106, 15406–15411. [Google Scholar] [CrossRef] [Green Version]
- Said, N.; Hilal, T.; Sunday, N.D.; Khatri, A.; Bürger, J.; Mielke, T.; Belogurov, G.A.; Loll, B.; Sen, R.; Artsimovitch, I.; et al. Steps toward translocation-independent RNA polymerase inactivation by terminator ATPase ρ. Science 2020, 371, eabd1673. [Google Scholar] [CrossRef]
- Leela, J.K.; Syeda, A.H.; Anupama, K.; Gowrishankar, J. Rho-dependent transcription termination is essential to prevent excessive genome-wide R-loops in Escherichia coli. Proc. Natl. Acad. Sci. USA 2012, 110, 258–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghunathan, N.; Kapshikar, R.M.; Leela, J.K.; Mallikarjun, J.; Bouloc, P.; Gowrishankar, J. Genome-wide relationship between R-loop formation and antisense transcription in Escherichia coli. Nucleic Acids Res. 2018, 46, 3400–3411. [Google Scholar] [CrossRef]
- Rowley, G.; Spector, M.; Kormanec, J.; Roberts, M. Pushing the envelope: Extracytoplasmic stress responses in bacterial pathogens. Nat. Rev. Microbiol. 2006, 4, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Paget, M.S. Bacterial Sigma Factors and Anti-Sigma Factors: Structure, Function and Distribution. Biomolecules 2015, 5, 1245–1265. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Coll, L.; Maciag-Dorszynska, M.; Tailor, K.; Vadia, S.; Levin, P.A.; Szalewska-Palasz, A.; Cashel, M. The Absence of (p)ppGpp Renders Initiation of Escherichia coli Chromosomal DNA Synthesis Independent of Growth Rates. mBio 2020, 11, e03223-19. [Google Scholar] [CrossRef] [Green Version]
- Rochman, M.; Aviv, M.; Glaser, G.; Muskhelishvili, G. Promoter protection by a transcription factor acting as a local topological homeostat. EMBO Rep. 2002, 3, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Potrykus, K.; Vinella, D.; Murphy, H.; Szalewska-Palasz, A.; D’Ari, R.; Cashel, M. Antagonistic regulation of Escherichia coli ribosomal RNA rrnB P1 promoter activity by GreA and DksA. J. Biol. Chem. 2006, 281, 15238–15248. [Google Scholar] [CrossRef] [Green Version]
- Ueshima, R.; Fujita, N.; Ishihama, A. DNA supercoiling and temperature shift affect the promoter activity of the Escherichia coli rpoH gene encoding the heat-shock sigma subunit of RNA polymerase. Mol. Genet. Genom. 1989, 215, 185–189. [Google Scholar] [CrossRef]
- Sudzinová, P.; Kambová, M.; Ramaniuk, O.; Benda, M.; Šanderová, H.; Krásný, L. Effects of DNA Topology on Transcription from rRNA Promoters in Bacillus subtilis. Microorganisms 2021, 9, 87. [Google Scholar] [CrossRef] [PubMed]
- Kusano, S.; Ding, Q.; Fujita, N.; Ishihama, A. Promoter selectivity of Escherichia coli RNA polymerase E sigma 70 and E sigma 38 holoenzymes. Effect of DNA supercoiling. J. Biol. Chem. 1996, 271, 1998–2004. [Google Scholar] [CrossRef] [Green Version]
- Schneider, R.; Travers, A.; Muskhelishvili, G. The expression of the Escherichia coli fis gene is strongly dependent on the superhelical density of DNA. Mol. Microbiol. 2000, 38, 167–175. [Google Scholar] [CrossRef]
- González-Gil, G.; Kahmann, R.; Muskhelishvili, G. Regulation of crp transcription by oscillation between distinct nucleoprotein complexes. EMBO J. 1998, 17, 2877–2885. [Google Scholar] [CrossRef] [PubMed]
- Levanon, S.S.; San, K.-Y.; Bennett, G.N. Effect of oxygen on the Escherichia coli ArcA and FNR regulation systems and metabolic responses. Biotechnol. Bioeng. 2005, 89, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Berger, M.; Gerganova, V.; Berger, P.; Rapiteanu, R.; Lisicovas, V.; Dobrindt, U. Genes on a Wire: The Nucleoid-Associated Protein HU Insulates Transcription Units in Escherichia coli. Sci. Rep. 2016, 6, 31512. [Google Scholar] [CrossRef] [Green Version]
- Claret, L.; Rouviere-Yaniv, J. Variation in HU composition during growth of Escherichia coli: The heterodimer is required for long term survival. J. Mol. Biol. 1997, 273, 93–104. [Google Scholar] [CrossRef]
- Zhou, Y.N.; Jin, D.J. The rpoB mutants destabilizing initiation complexes at stringently controlled promoters behave like “stringent” RNA polymerases in Escherichia coli. Proc. Natl. Acad. Sci. USA 1998, 95, 2908–2913. [Google Scholar] [CrossRef] [Green Version]
- Balke, V.L.; Gralla, J.D. Changes in the linking number of supercoiled DNA accompany growth transitions in Escherichia coli. J. Bacteriol. 1987, 169, 4499–4506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, M.B.; Roth, J.R. Gene location affects expression level in Salmonella typhimurium. J. Bacteriol. 1987, 169, 2872–2875. [Google Scholar] [CrossRef] [Green Version]
- Sousa, C.; de Lorenzo, V.; Cebolla, A. Modulation of gene expression through chromosomal positioning in Escherichia coli. Microbiology 1997, 143, 2071–2078. [Google Scholar] [CrossRef] [Green Version]
- Rocha, E.P.; Danchin, A. Gene essentiality determines chromosome organisation in bacteria. Nucleic Acids Res. 2003, 31, 6570–6577. [Google Scholar] [CrossRef] [Green Version]
- Fang, G.; Rocha, E.P.; Danchin, A. Persistence drives gene clustering in bacterial genomes. BMC Genom. 2008, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Sobetzko, P.; Travers, A.; Muskhelishvili, G. Gene order and chromosome dynamics coordinate spatiotemporal gene expression during the bacterial growth cycle. Proc. Natl. Acad. Sci. USA 2011, 109, E42–E50. [Google Scholar] [CrossRef] [Green Version]
- Kosmidis, K.; Jablonski, K.P.; Muskhelishvili, G.; Hütt, M.-T. Chromosomal origin of replication coordinates logically distinct types of bacterial genetic regulation. NPJ Syst. Biol. Appl. 2020, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gerganova, V.; Berger, M.F.; Zaldastanishvili, E.; Sobetzko, P.; Lafon, C.; Mourez, M.; Travers, A.; Muskhelishvili, G. Chromosomal position shift of a regulatory gene alters the bacterial phenotype. Nucleic Acids Res. 2015, 43, 8215–8226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soler-Bistué, A.; Timmermans, M.; Mazel, D. The Proximity of Ribosomal Protein Genes to oriC Enhances Vibrio cholerae Fitness in the Absence of Multifork Replication. mBio 2017, 8, e00097-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogue, M.M.; Mogre, A.; Beckett, M.C.; Thomson, N.R.; Dorman, C.J. Network Rewiring: Physiological Consequences of Reciprocally Exchanging the Physical Locations and Growth-Phase-Dependent Expression Patterns of the Salmonella fis and dps Genes. mBio 2020, 11, e02128-20. [Google Scholar] [CrossRef]
- Nigatu, D.; Henkel, W.; Sobetzko, P.; Muskhelishvili, G. Relationship between digital information and thermodynamic stability in bacterial genomes. EURASIP J. Bioinform. Syst. Biol. 2016, 2016, 4555. [Google Scholar] [CrossRef] [Green Version]
- Travers, A.A.; Muskhelishvili, G. DNA thermodynamics shape chromosome organization and topology. Biochem. Soc. Trans. 2013, 41, 548–553. [Google Scholar] [CrossRef] [Green Version]
- Meyer, S.; Reverchon, S.; Nasser, W.; Muskhelishvili, G. Chromosomal organization of transcription: In a nutshell. Curr. Genet. 2017, 64, 555–565. [Google Scholar] [CrossRef]
- Muskhelishvili, G.; Forquet, R.; Reverchon, S.; Meyer, S.; Nasser, W. Coherent Domains of Transcription Coordinate Gene Expression During Bacterial Growth and Adaptation. Microorganisms 2019, 7, 694. [Google Scholar] [CrossRef] [Green Version]
- Reverchon, S.; Meyer, S.; Forquet, R.; Hommais, F.; Muskhelishvili, G.; Nasser, W. The nucleoid-associated protein IHF acts as a ‘transcriptional domainin’ protein coordinating the bacterial virulence traits with global transcription. Nucleic Acids Res. 2020, 49, 776–790. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Tanaka, K.; Ishihama, A. The whole set of the constitutive promoters recognized by four minor sigma subunits of Escherichia coli RNA polymerase. PLoS ONE 2017, 12, e0179181. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, M.; Elliott, T.; Hirsch, M.; Elliott, T. Fis regulates transcriptional induction of RpoS in Salmonella enterica. J. Bacteriol. 2005, 187, 1568–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, P.A. Making DNA do a U-turn: IHF and related proteins. Curr. Opin. Struct. Biol. 1997, 7, 86–93. [Google Scholar] [CrossRef]
- Pagel, J.M.; Winkelman, J.W.; Adams, C.W.; Hatfield, G. DNA topology-mediated regulation of transcription initiation from the tandem promoters of the ilvGMEDA operon of Escherichia coli. J. Mol. Biol. 1992, 224, 919–935. [Google Scholar] [CrossRef]
- Travers, A. DNA-protein interactions: IHF—The master bender. Curr. Biol. 1997, 7, R252–R254. [Google Scholar] [CrossRef] [Green Version]
- Ellenberger, T.; Landy, A. A good turn for DNA: The structure of integration host factor bound to DNA. Structure 1997, 5, 153–157. [Google Scholar] [CrossRef] [Green Version]
- Green, J.; Scott, C.; Guest, J.R. Functional versatility in the CRP-FNR superfamily of transcription factors: FNR and FLP. Adv. Microb. Physiol. 2001, 44, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Amouyal, M.; Buc, H. Topological unwinding of strong and weak promoters by RNA polymerase: A comparison between the lac wild-type and the UV5 sites of Escherichia coli. J. Mol. Biol. 1987, 195, 795–808. [Google Scholar] [CrossRef]
- Shin, M.; Song, M.; Rhee, J.H.; Hong, Y.; Kim, Y.-J.; Seok, Y.-J.; Ha, K.-S.; Jung, S.-H.; Choy, H.E. DNA looping-mediated repression by histone-like protein H-NS: Specific requirement of Esigma70 as a cofactor for looping. Genes Dev. 2005, 19, 2388–2398. [Google Scholar] [CrossRef] [Green Version]
- Cellai, S.; Mangiarotti, L.; Vannini, N.; Naryshkin, N.; Kortkhonjia, E.; Ebright, R.H.; Rivetti, C. Upstream promoter sequences and alphaCTD mediate stable DNA wrapping within the RNA polymerase–promoter open complex. EMBO Rep. 2007, 8, 271–278. [Google Scholar] [CrossRef]
- Dorman, C.J.; Schumacher, M.A.; Bush, M.; Brennan, R.G.; Buttner, M.J. When is a transcription factor a NAP? Curr. Opin. Microbiol. 2020, 55, 26–33. [Google Scholar] [CrossRef]
- Harman, J.G. Allosteric regulation of the cAMP receptor protein. Biochim. Biophys. Acta. 2001, 1547, 1–17. [Google Scholar] [CrossRef]
- Mettert, E.L.; Kiley, P.J. Reassessing the Structure and Function Relationship of the O2 Sensing Transcription Factor FNR. Antioxid. Redox Signal. 2018, 29, 1830–1840. [Google Scholar] [CrossRef]
- Alba, B.M.; Gross, C.A. Regulation of the Escherichia coli sigma-dependent envelope stress response. Mol. Microbiol. 2004, 52, 613–619. [Google Scholar] [CrossRef]
- Rhodius, V.A.; Suh, W.C.; Nonaka, G.; West, J.; Gross, C.A. Conserved and variable Functions of the sigmaE stress response in related genomes. PLoS Biol. 2005, 4, e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hengge-Aronis, R. Signal transduction and regulatory mechanisms involved in control of the sigma(S) (RpoS) subunit of RNA polymerase. Microbiol. Mol. Biol. Rev. 2002, 66, 373–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordes, P.; Conter, A.; Morales, V.; Bouvier, J.; Kolb, A.; Gutierrez, C. DNA supercoiling contributes to disconnect sigmaS accumulation from sigmaS-dependent transcription in Escherichia coli. Mol. Microbiol. 2003, 48, 561–571. [Google Scholar] [CrossRef]
- Janga, S.C.; Salgado, H.; Martínez-Antonio, A. Transcriptional regulation shapes the organization of genes on bacterial chromosomes. Nucleic Acids Res. 2009, 37, 3680–3688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llopis, P.M.; Jackson, A.F.; Sliusarenko, O.; Surovtsev, I.; Heinritz, J.; Emonet, T.; Jacobs-Wagner, C. Spatial organization of the flow of genetic information in bacteria. Nature 2010, 466, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Kuhlman, T.E.; Cox, E.C. Gene location and DNA density determine transcription factor distributions in Escherichia coli. Mol. Syst. Biol. 2012, 8, 610. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, M.; Li, S.; Heermann, D.W.; Wiggins, P.A. A model for Escherichia coli chromosome packaging supports transcription factor-induced DNA domain formation. Nucleic Acids Res. 2011, 40, 972–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, C.D.; Cozzarelli, N.R. A genetic selection for supercoiling mutants of Escherichia coli reveals proteins implicated in chromosome structure. Mol. Microbiol. 2005, 57, 1636–1652. [Google Scholar] [CrossRef]
- Wu, F.; Japaridze, A.; Zheng, X.; Wiktor, J.; Kerssemakers, J.W.J.; Dekker, C. Direct imaging of the circular chromosome in a live bacterium. Nat. Commun. 2019, 10, 2194. [Google Scholar] [CrossRef] [Green Version]
- Képès, F.; Jester, B.C.; Lepage, T.; Rafiei, N.; Rosu, B.; Junier, I. The layout of a bacterial genome. FEBS Lett. 2012, 586, 2043–2048. [Google Scholar] [CrossRef]
- Képès, F. Periodic epi-organization of the yeast genome revealed by the distribution of promoter sites. J. Mol. Biol. 2003, 329, 859–865. [Google Scholar] [CrossRef]
- Couturier, E.; Rocha, E.P.C. Replication-associated gene dosage effects shape the genomes of fast-growing bacteria but only for transcription and translation genes. Mol. Microbiol. 2006, 59, 1506–1518. [Google Scholar] [CrossRef]
- Norris, V.; Blaauwen, T.D.; Cabin-Flaman, A.; Doi, R.H.; Harshey, R.; Janniere, L.; Jimenez-Sanchez, A.; Jin, D.J.; Levin, P.A.; Mileykovskaya, E.; et al. Functional taxonomy of bacterial hyperstructures. Microbiol. Mol. Biol. Rev. 2007, 71, 230–253. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Hypernegative Supercoiling | DNA relaxation | ||||||
|---|---|---|---|---|---|---|---|---|
| Strain, Background | LZ54WT | ∆fis | ∆hns | ∆fis/hns | LZ41WT | ∆fis | ∆hns | ∆fis/hns |
| Genes of NAPs and global TFs | fis | lrp | lrp hupB ihfA stpA | hupA | fnr ihfB | fnr | fnr crp | |
| Genes of RNAP modulators | crl nusG nusB | crl rho | crl nusG | nusG rho | greA | greA | ||
| Genes of the RNAP sigma and anti-sigma factors | rpoE rseAB | rpoE | rseA | rpoH | rpoD | rpoH rpoD fecI | rpoH rpoD fliA | |
| Topoisomerase genes | yrdD parC | yrdD parC | yrdD parC | yrdD parE | gyrA gyrB | gyrB | gyrB | gyrA gyrB |
| Strain, Condition | CF1943 Wild Type | CF2790 rpoZ | CF2790 rpoZ + rpoD * | CF2790 rpoZ |
|---|---|---|---|---|
| Impacts of NAPs & global TFs | FIS, IHF | CRP | IHF, CRP, Fnr | H-NS, FIS, Lrp |
| Impacts of RNAP sigma subunits | RpoD | RpoS | RpoD | RpoS |
| Regulons up | RpoS | CRP, Fnr | RpoS | |
| Couplons up | H-NS/RpoS | CRP/RpoD Fnr/RpoD | CRP/RpoS FIS/RpoS Lrp/RpoS IHF/RpoS Fnr/RpoS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muskhelishvili, G.; Sobetzko, P.; Mehandziska, S.; Travers, A. Composition of Transcription Machinery and Its Crosstalk with Nucleoid-Associated Proteins and Global Transcription Factors. Biomolecules 2021, 11, 924. https://doi.org/10.3390/biom11070924
Muskhelishvili G, Sobetzko P, Mehandziska S, Travers A. Composition of Transcription Machinery and Its Crosstalk with Nucleoid-Associated Proteins and Global Transcription Factors. Biomolecules. 2021; 11(7):924. https://doi.org/10.3390/biom11070924
Chicago/Turabian StyleMuskhelishvili, Georgi, Patrick Sobetzko, Sanja Mehandziska, and Andrew Travers. 2021. "Composition of Transcription Machinery and Its Crosstalk with Nucleoid-Associated Proteins and Global Transcription Factors" Biomolecules 11, no. 7: 924. https://doi.org/10.3390/biom11070924
APA StyleMuskhelishvili, G., Sobetzko, P., Mehandziska, S., & Travers, A. (2021). Composition of Transcription Machinery and Its Crosstalk with Nucleoid-Associated Proteins and Global Transcription Factors. Biomolecules, 11(7), 924. https://doi.org/10.3390/biom11070924