
Impact of a Single Nucleotide Change or Non-Nucleoside Modifications in G-Rich Region on the Quadruplex–Duplex Hybrid Formation

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
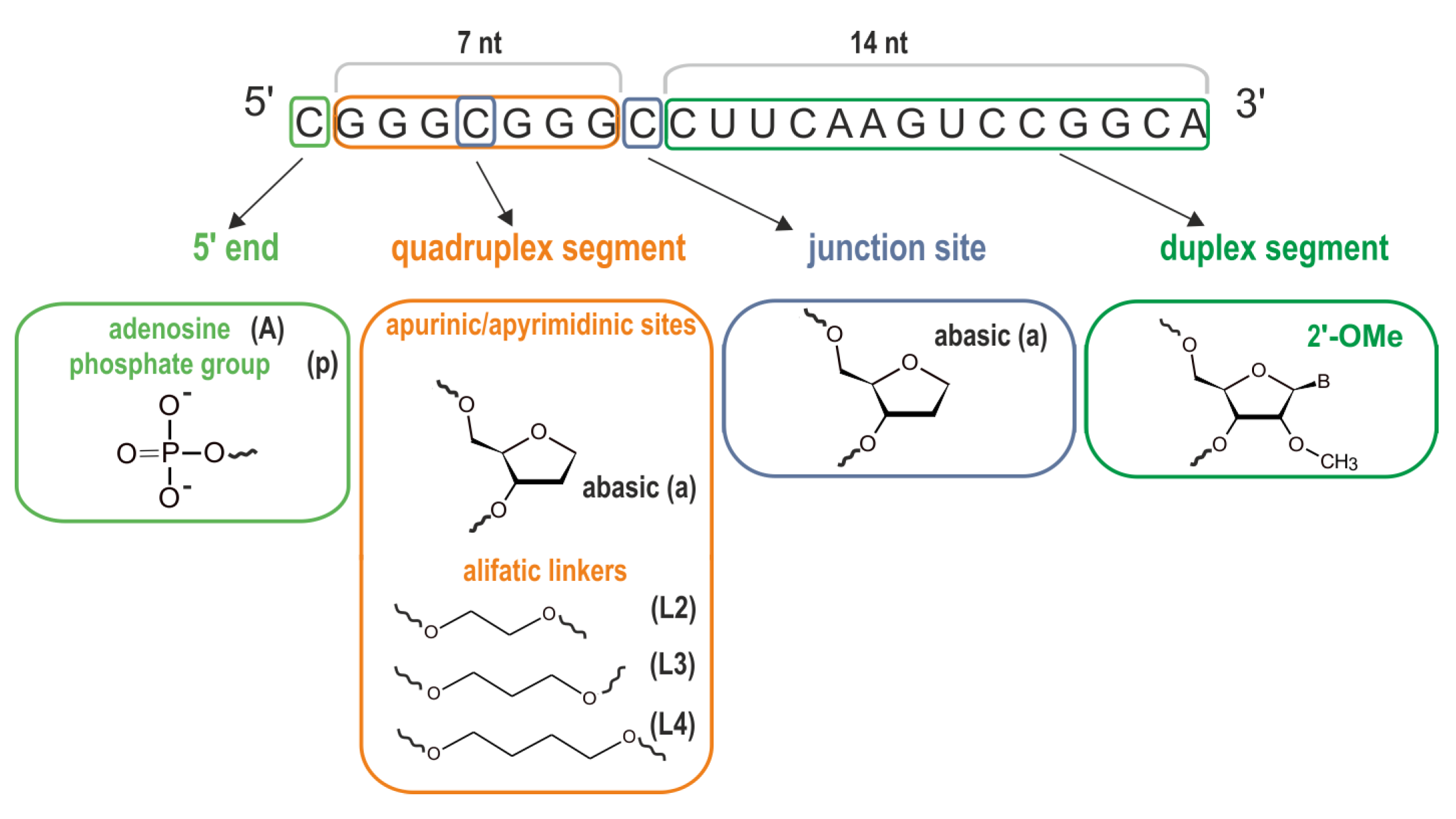
2.1. Synthesis and Purification of DNA and RNA Oligonucleotides
2.2. NH2-CCC Labeling with o-BMVC-C3
2.3. Denaturing Electrophoresis of RNA Oligo-o-BMVC-C3
2.4. Non-Denaturing Electrophoresis of Oligonucleotides
2.5. NMR Experiments
2.6. UV Thermal Denaturation Curves
2.7. Fluorescence Measurements
2.8. CD Measurements
3. Results
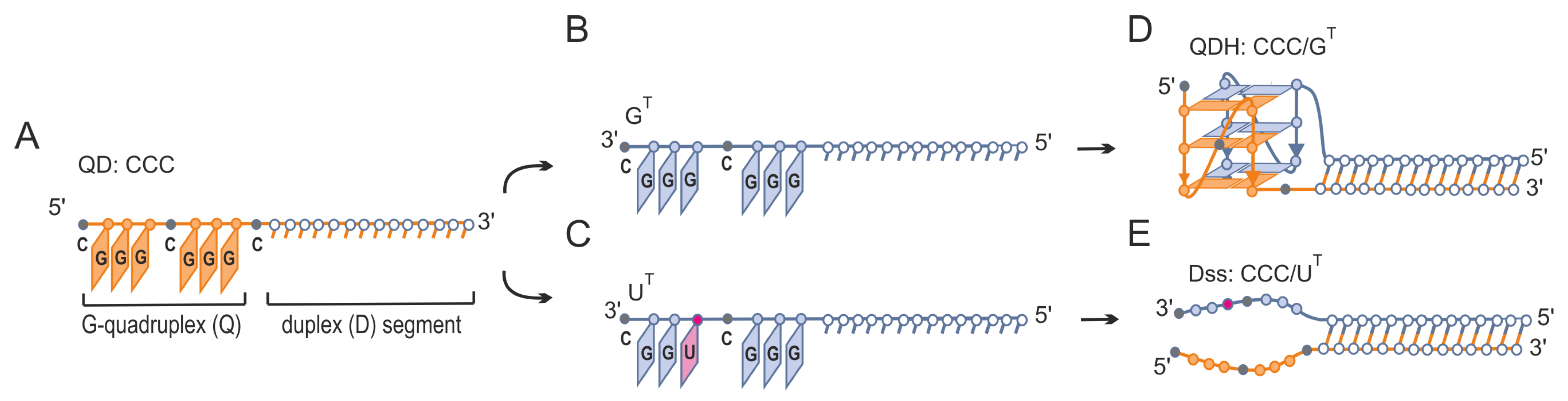
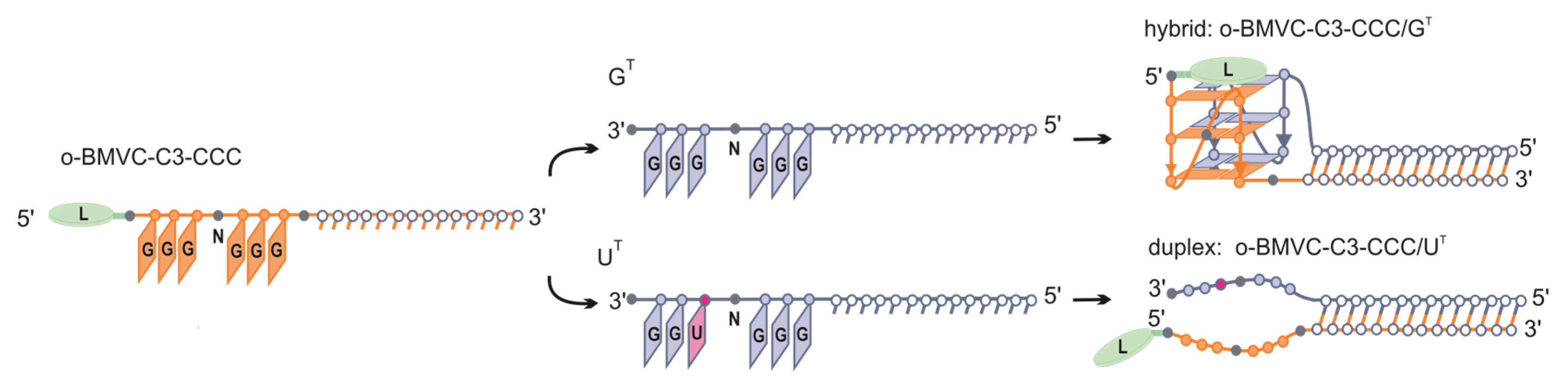
3.1. Formation of Alternative Structures by Hybridization of G-Rich Oligonucleotide to Target Sequences That Differ by a Single Nucleotide Residue
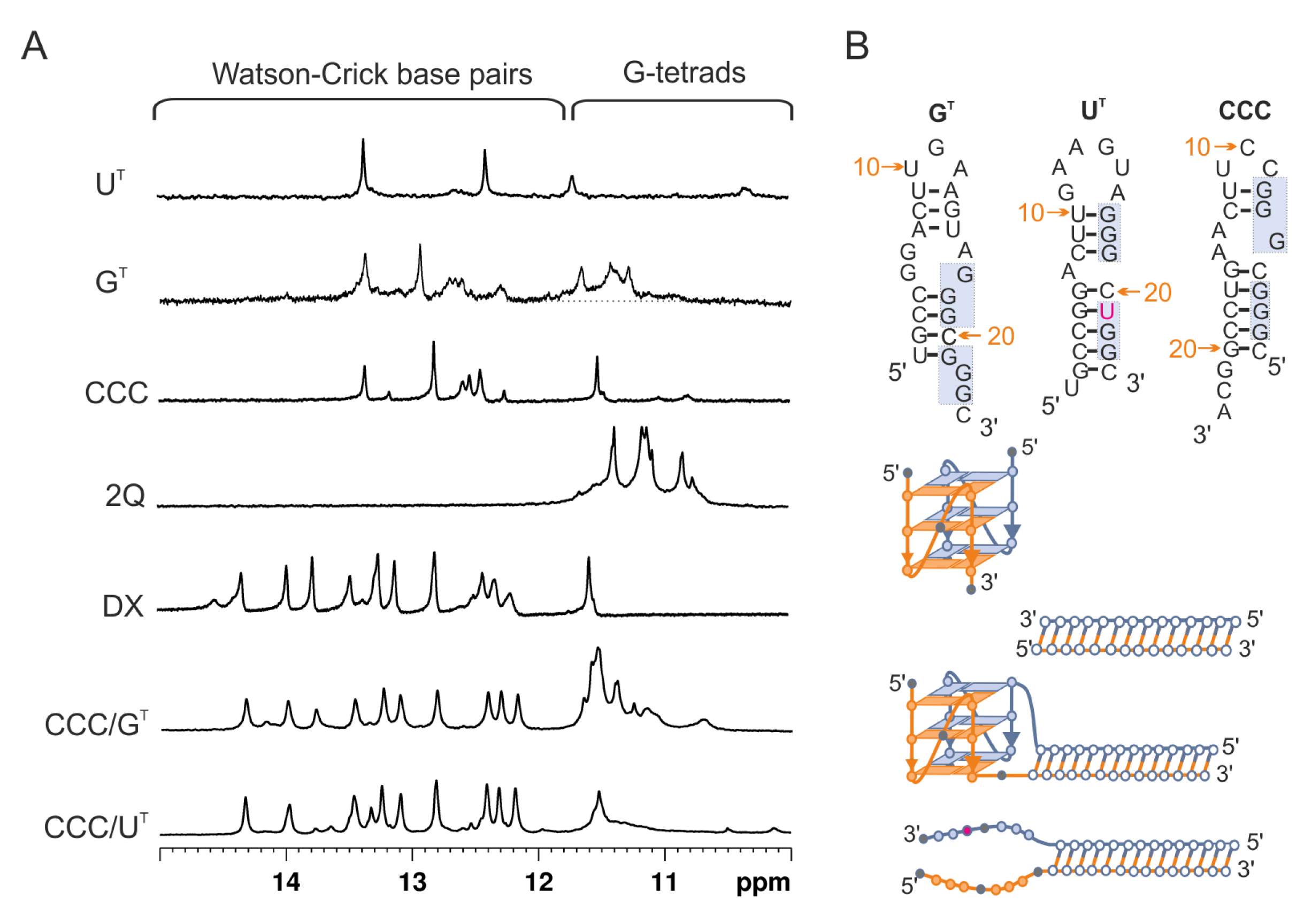
3.2. Evaluation of Secondary Structures of CCC/GT and CCC/UT RNA:RNA Complexes
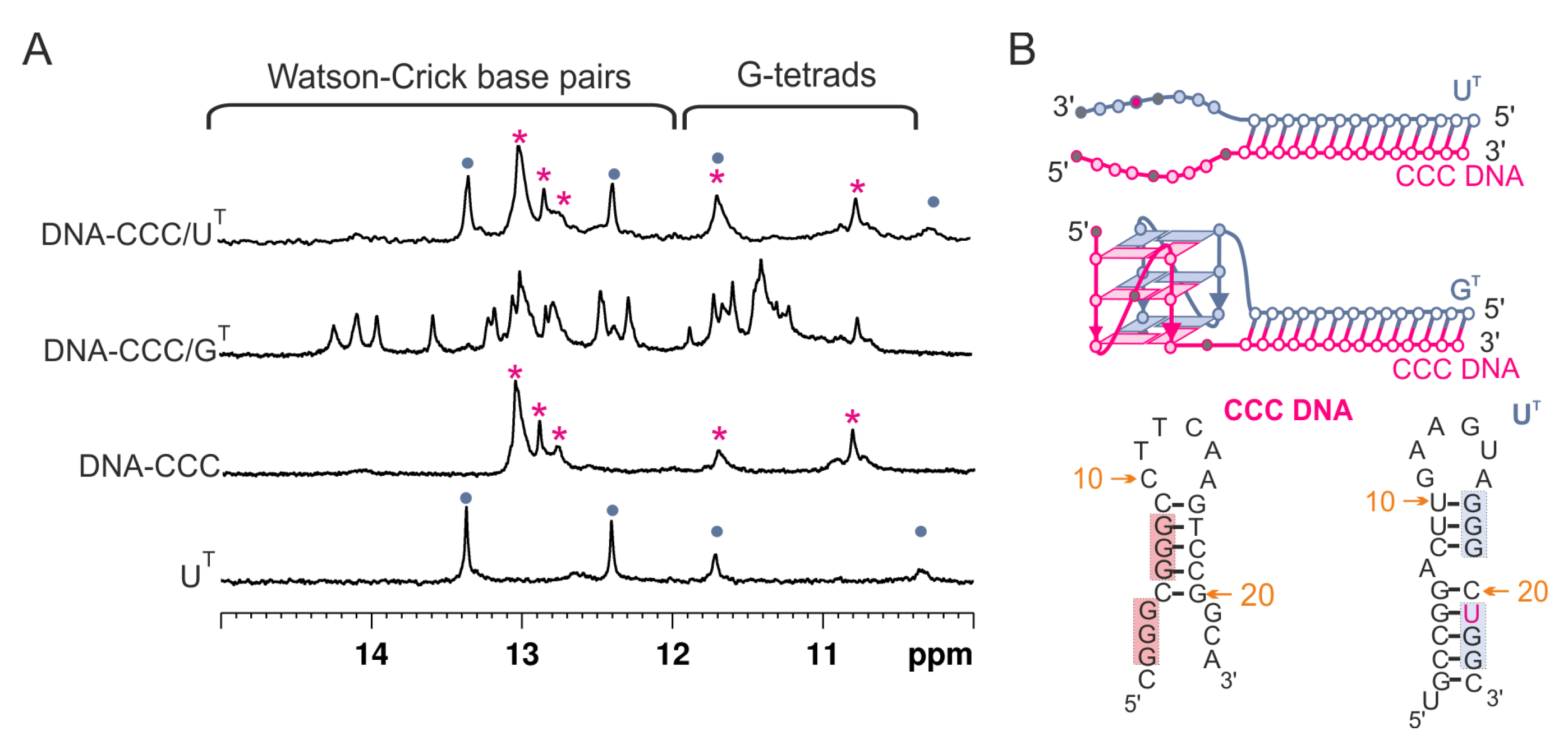
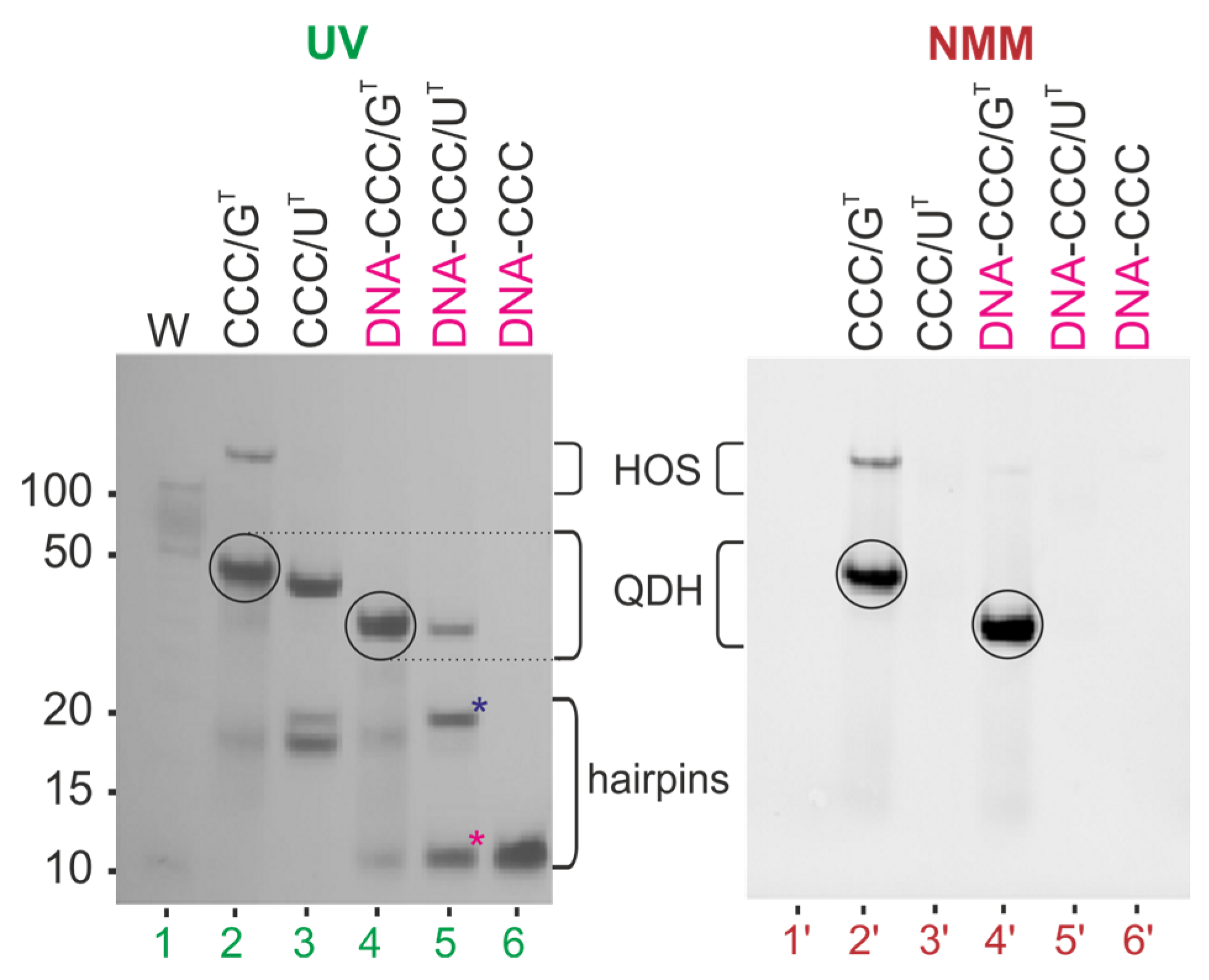
3.3. Evaluation of Secondary Structures of DNA-CCC/GT and DNA-CCC/UT Complexes
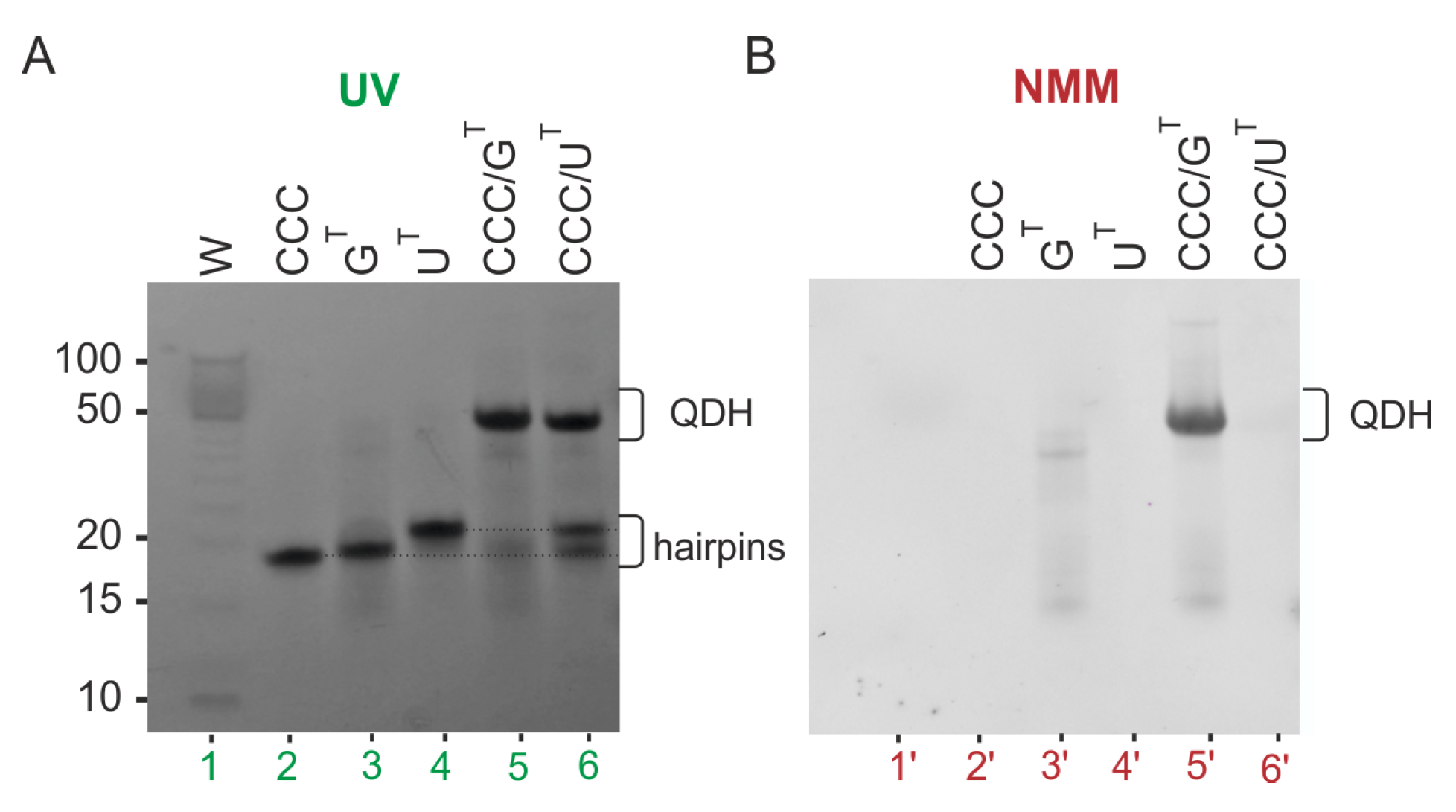
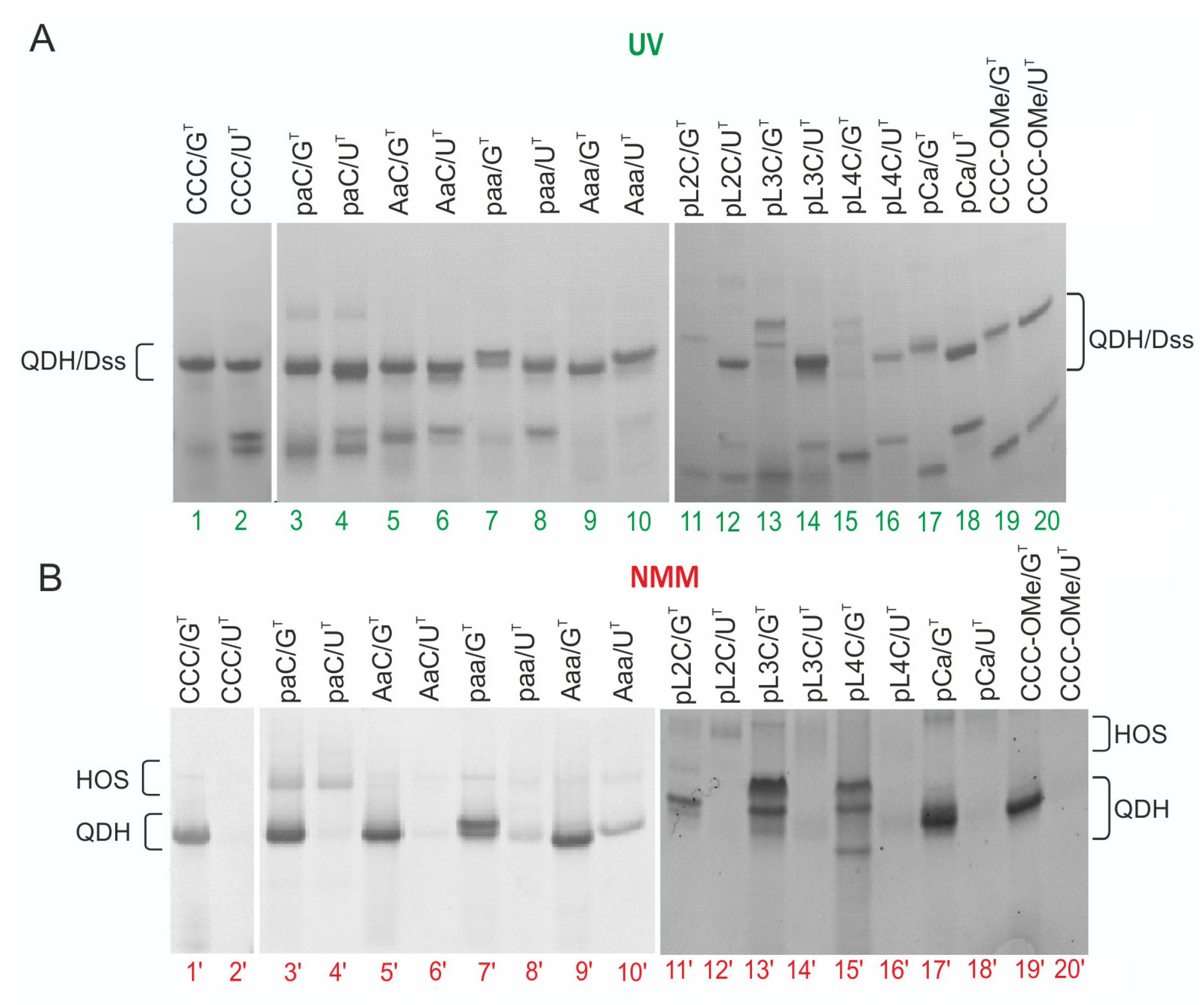
3.4. Impact of the Chemical Modifications on the Ability to Form Alternate QDH and Dss Structures
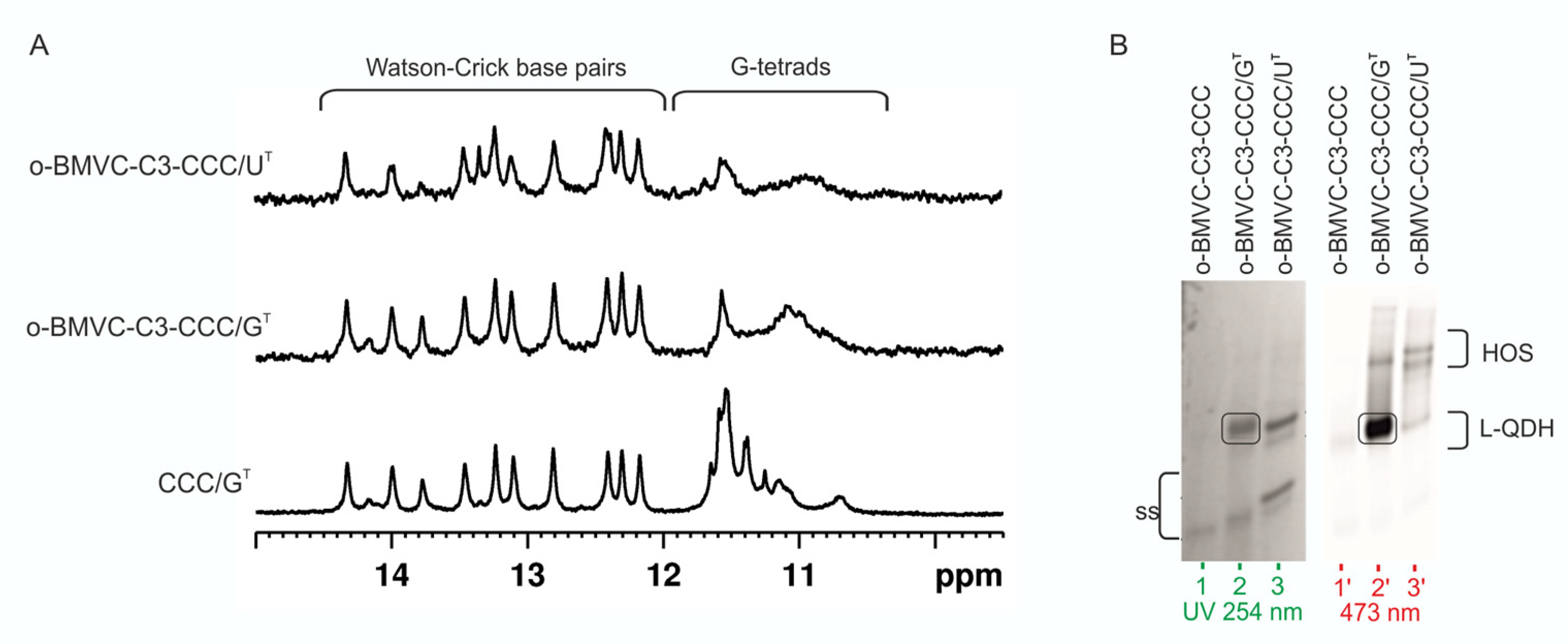
3.5. Influence of CCC Oligonucleotide with Covalently Attached G4 Ligand on the Formation and Stability of QDH and Dss Structures
4. Discussion
4.1. Quadruplex–Duplex Hybrid Structures (RNA vs. DNA)
4.2. Stability of QDH and Dss Structures
4.3. Formation of Non-Canonical Hybrid Structures
4.4. Application Potential of G-Rich Oligonucleotides
4.5. G4 Ligand Conjugated to Oligonucleotide
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Burge, S.; Parkinson, G.N.; Hazel, P.; Todd, A.K.; Neidle, S. Quadruplex DNA: Sequence, topology and structure. Nucleic Acids Res. 2006, 34, 5402–5415. [Google Scholar] [CrossRef] [Green Version]
- Parkinson, G.N. Fundamentals of Quadruplex Structures. In Quadruplex Nucleic Acids; Neidle, S., Balasubramanian, S., Eds.; Royal Society of Chemistry: London, UK, 2007; pp. 1–30. [Google Scholar]
- Han, J.H.; Kim, J.H.; Kim, S.K.; Jang, Y.J. Conformational change of a G-quadruplex under molecular crowding conditions. J. Biomol. Struct. Dyn. 2020, 38, 2575–2581. [Google Scholar] [CrossRef]
- Chilka, P.; Desai, N.; Datta, B. Small Molecule Fluorescent Probes for G- Quadruplex Visualization as Potential Cancer Theranostic Agents. Molecules 2019, 24, 752. [Google Scholar] [CrossRef] [Green Version]
- Kwok, C.K.; Merrick, C.J. G-Quadruplexes: Prediction, Characterization, and Biological Application. Trends Biotechnol. 2017, 35, 997–1013. [Google Scholar] [CrossRef] [PubMed]
- Dumas, L.; Herviou, P.; Dassi, E.; Cammas, A.; Millevoi, S. G-Quadruplexes in RNA Biology: Recent Advances and Future Directions. Trends Biochem. Sci. 2021, 46, 270–283. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.-D.; Shibata, T.; Yamamoto, Y.; Xu, Y. An intramolecular antiparallel G-quadruplex formed by human telomere RNA. Chem. Commun. 2018, 54, 3944–3946. [Google Scholar] [CrossRef] [PubMed]
- Banco, M.T.; Ferré-D’Amaré, A.R. The emerging structural complexity of G-quadruplex RNAs. RNA 2021, 27, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Trachman, R.J.; Autour, A.; Jeng, S.C.Y.; Abdolahzadeh, A.; Andreoni, A.; Cojocaru, R.; Garipov, R.; Dolgosheina, E.V.; Knutson, J.R.; Ryckelynck, M.; et al. Structure and functional reselection of the Mango-III fluorogenic RNA aptamer. Nat. Chem. Biol. 2019, 15, 472–479. [Google Scholar] [CrossRef]
- Trachman, R.J.; Demeshkina, N.A.; Lau, M.W.L.; Panchapakesan, S.S.S.; Jeng, S.C.Y.; Unrau, P.J.; Ferré-D’Amaré, A.R. Structural basis for high-affinity fluorophore binding and activation by RNA Mango. Nat. Chem. Biol. 2017, 13, 807–813. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Suslov, N.B.; Li, N.-S.; Shelke, S.A.; Evans, M.E.; Koldobskaya, Y.; Rice, P.A.; Piccirilli, J.A. A G-quadruplex–containing RNA activates fluorescence in a GFP-like fluorophore. Nat. Chem. Biol. 2014, 10, 686–691. [Google Scholar] [CrossRef]
- Haase, L.; Karg, B.; Weisz, K. Manipulating DNA G-Quadruplex Structures by Using Guanosine Analogues. ChemBioChem 2019, 20, 985–993. [Google Scholar] [CrossRef]
- Sagi, J. G-quadruplexes incorporating modified constituents: A review. J. Biomol. Struct. Dyn. 2014, 32, 477–511. [Google Scholar] [CrossRef] [PubMed]
- Lech, C.J.; Li, Z.; Heddi, B.; Phan, A.T. 2′-F-ANA-guanosine and 2′-F-guanosine as powerful tools for structural manipulation of G-quadruplexes. Chem. Commun. 2012, 48, 11425. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lech, C.J.; Phan, A.T. Sugar-modified G-quadruplexes: Effects of LNA-, 2′F-RNA– and 2′F-ANA-guanosine chemistries on G-quadruplex structure and stability. Nucleic Acids Res. 2014, 42, 4068–4079. [Google Scholar] [CrossRef] [PubMed]
- Risitano, A. Influence of loop size on the stability of intramolecular DNA quadruplexes. Nucleic Acids Res. 2004, 32, 2598–2606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rachwal, P.A.; Brown, T.; Fox, K.R. Sequence effects of single base loops in intramolecular quadruplex DNA. FEBS Lett. 2007, 581, 1657–1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gudanis, D.; Popenda, L.; Szpotkowski, K.; Kierzek, R.; Gdaniec, Z. Structural characterization of a dimer of RNA duplexes composed of 8-bromoguanosine modified CGG trinucleotide repeats: A novel architecture of RNA quadruplexes. Nucleic Acids Res. 2016, 44, 2409–2416. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.; Sun, H.; Zhou, H.; Xiang, J.; Tang, Y.; Zhao, C. Research progress of RNA quadruplex. Nucleic Acid Ther. 2011, 21, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Bhavesh, N.S.; Patel, P.K.; Karthikeyan, S.; Hosur, R.V. Distinctive features in the structure and dynamics of the DNA repeat sequence GGCGGG. Biochem. Biophys. Res. Commun. 2004, 317, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Mergny, J.L.; De Cian, A.; Amrane, S.; da Silva, M.W. Kinetics of double-chain reversals bridging contiguous quartets in tetramolecular quadruplexes. Nucleic Acids Res. 2006, 34, 2386–2397. [Google Scholar] [CrossRef] [Green Version]
- Malgowska, M.; Gudanis, D.; Kierzek, R.; Wyszko, E.; Gabelica, V.; Gdaniec, Z. Distinctive structural motifs of RNA G-quadruplexes composed of AGG, CGG and UGG trinucleotide repeats. Nucleic Acids Res. 2014, 42, 10196–10207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lightfoot, H.L.; Hagen, T.; Tatum, N.J.; Hall, J. The diverse structural landscape of quadruplexes. FEBS Lett. 2019, 593, 2083–2102. [Google Scholar] [CrossRef] [PubMed]
- Malgowska, M.; Czajczynska, K.; Gudanis, D.; Tworak, A.; Gdaniec, Z. Overview of the RNA G-quadruplex structures. Acta Biochim. Pol. 2016, 63, 609–621. [Google Scholar] [CrossRef]
- Andrałojć, W.; Małgowska, M.; Sarzyńska, J.; Pasternak, K.; Szpotkowski, K.; Kierzek, R.; Gdaniec, Z. Unraveling the structural basis for the exceptional stability of RNA G-quadruplexes capped by a uridine tetrad at the 3′ terminus. RNA 2019, 25, 121–134. [Google Scholar] [CrossRef]
- Andrałojć, W.; Pasternak, K.; Sarzyńska, J.; Zielińska, K.; Kierzek, R.; Gdaniec, Z. The origin of the high stability of 3′-terminal uridine tetrads: Contributions of hydrogen bonding, stacking interactions, and steric factors evaluated using modified oligonucleotide analogs. RNA 2020, 26, 2000–2016. [Google Scholar] [CrossRef]
- Esposito, V.; Martino, L.; Citarella, G.; Virgilio, A.; Mayol, L.; Giancola, C.; Galeone, A. Effects of abasic sites on structural, thermodynamic and kinetic properties of quadruplex structures. Nucleic Acids Res. 2009, 38, 2069–2080. [Google Scholar] [CrossRef] [PubMed]
- Švehlová, K.; Lawrence, M.S.; Bednárová, L.; Curtis, E.A. Altered biochemical specificity of G-quadruplexes with mutated tetrads. Nucleic Acids Res. 2016, 44, 10789–10803. [Google Scholar] [CrossRef]
- Endoh, T.; Kawasaki, Y.; Sugimoto, N. Stability of RNA quadruplex in open reading frame determines proteolysis of human estrogen receptor α. Nucleic Acids Res. 2013, 41, 6222–6231. [Google Scholar] [CrossRef] [Green Version]
- Endoh, T.; Sugimoto, N. Conformational Dynamics of the RNA G-Quadruplex and its Effect on Translation Efficiency. Molecules 2019, 24, 1613. [Google Scholar] [CrossRef] [Green Version]
- Chambers, V.S.; Marsico, G.; Boutell, J.M.; Di Antonio, M.; Smith, G.P.; Balasubramanian, S. High-throughput sequencing of DNA G-quadruplex structures in the human genome. Nat. Biotechnol. 2015, 33, 877–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huppert, J.L. Prevalence of quadruplexes in the human genome. Nucleic Acids Res. 2005, 33, 2908–2916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwok, C.K.; Marsico, G.; Sahakyan, A.B.; Chambers, V.S.; Balasubramanian, S. rG4-seq reveals widespread formation of G-quadruplex structures in the human transcriptome. Nat. Methods 2016, 13, 841–844. [Google Scholar] [CrossRef] [PubMed]
- Kharel, P.; Becker, G.; Tsvetkov, V.; Ivanov, P. Properties and biological impact of RNA G-quadruplexes: From order to turmoil and back. Nucleic Acids Res. 2020, 1–22. [Google Scholar] [CrossRef]
- Tassinari, M.; Richter, S.N.; Gandellini, P. Biological relevance and therapeutic potential of G-quadruplex structures in the human noncoding transcriptome. Nucleic Acids Res. 2021, 49, 3617–3633. [Google Scholar] [CrossRef]
- Kharel, P.; Balaratnam, S.; Beals, N.; Basu, S. The role of RNA G-quadruplexes in human diseases and therapeutic strategies. Wiley Interdiscip. Rev. RNA 2020, 11, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Lyons, S.M.; Gudanis, D.; Coyne, S.M.; Gdaniec, Z.; Ivanov, P. Identification of functional tetramolecular RNA G-quadruplexes derived from transfer RNAs. Nat. Commun. 2017, 8, 1127. [Google Scholar] [CrossRef] [Green Version]
- Jackowiak, P.; Hojka-Osinska, A.; Gasiorek, K.; Stelmaszczuk, M.; Gudanis, D.; Gdaniec, Z.; Figlerowicz, M. Effects of G-quadruplex topology on translational inhibition by tRNA fragments in mammalian and plant systems in vitro. Int. J. Biochem. Cell Biol. 2017, 92, 148–154. [Google Scholar] [CrossRef]
- Lim, K.W.; Jenjaroenpun, P.; Low, Z.J.; Khong, Z.J.; Ng, Y.S.; Kuznetsov, V.A.; Phan, A.T. Duplex stem-loop-containing quadruplex motifs in the human genome: A combined genomic and structural study. Nucleic Acids Res. 2015, 43, 5630–5646. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.Q.N.; Lim, K.W.; Phan, A.T. Folding Kinetics of G-Quadruplexes: Duplex Stem Loops Drive and Accelerate G-Quadruplex Folding. J. Phys. Chem. B 2020, 124, 5122–5130. [Google Scholar] [CrossRef]
- Lim, K.W.; Phan, A.T. Structural basis of DNA quadruplex-duplex junction formation. Angew. Chemie Int. Ed. 2013, 52, 8566–8569. [Google Scholar] [CrossRef]
- Lim, K.W.; Khong, Z.J.; Phan, A.T. Thermal stability of DNA quadruplex-duplex hybrids. Biochemistry 2014, 53, 247–257. [Google Scholar] [CrossRef]
- Butovskaya, E.; Heddi, B.; Bakalar, B.; Richter, S.N.; Phan, A.T. Major G-Quadruplex Form of HIV-1 LTR Reveals a (3 + 1) Folding Topology Containing a Stem-Loop. J. Am. Chem. Soc. 2018, 140, 13654–13662. [Google Scholar] [CrossRef] [Green Version]
- Lim, K.W.; Nguyen, T.Q.N.; Phan, A.T. Joining of Multiple Duplex Stems at a Single Quadruplex Loop. J. Am. Chem. Soc. 2014, 136, 17969–17973. [Google Scholar] [CrossRef] [PubMed]
- Ngoc Nguyen, T.Q.; Lim, K.W.; Phan, A.T. Duplex formation in a G-quadruplex bulge. Nucleic Acids Res. 2020, 1–9. [Google Scholar] [CrossRef]
- Vianney, Y.M.; Preckwinkel, P.; Mohr, S.; Weisz, K. Quadruplex–Duplex Junction: A High-Affinity Binding Site for Indoloquinoline Ligands. Chem. A Eur. J. 2020, 26, 16910–16922. [Google Scholar] [CrossRef]
- Karg, B.; Mohr, S.; Weisz, K. Duplex-Guided Refolding into Novel G-Quadruplex (3+1) Hybrid Conformations. Angew. Chemie Int. Ed. 2019, 58, 11068–11071. [Google Scholar] [CrossRef]
- Greco, M.L.; Kotar, A.; Rigo, R.; Cristofari, C.; Plavec, J.; Sissi, C. Coexistence of two main folded G-quadruplexes within a single G-rich domain in the EGFR promoter. Nucleic Acids Res. 2017, 45, 10132–10142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, D.J.Y.; Winnerdy, F.R.; Lim, K.W.; Phan, A.T. Coexistence of two quadruplex–duplex hybrids in the PIM1 gene. Nucleic Acids Res. 2020, 48, 11162–11171. [Google Scholar] [CrossRef] [PubMed]
- Hagihara, M.; Yamauchi, L.; Seo, A.; Yoneda, K.; Senda, M.; Nakatani, K. Antisense-induced guanine quadruplexes inhibit reverse transcription by HIV-1 reverse transcriptase. J. Am. Chem. Soc. 2010, 132, 11171–11178. [Google Scholar] [CrossRef]
- Bhattacharyya, D.; Nguyen, K.; Basu, S. Rationally Induced RNA:DNA G-Quadruplex Structures Elicit an Anticancer Effect by Inhibiting Endogenous eIF-4E Expression. Biochemistry 2014, 53, 5461–5470. [Google Scholar] [CrossRef]
- Gudanis, D.; Kaniowski, D.; Kulik, K.; Baranowski, D.; Gdaniec, Z.; Nawrot, B. Formation of an RNA Quadruplex-Duplex Hybrid in Living Cells between mRNA of the Epidermal Growth Factor Receptor (EGFR) and a G-Rich Antisense Oligoribonucleotide. Cells 2020, 9, 2375. [Google Scholar] [CrossRef]
- Tan, D.J.Y.; Das, P.; Winnerdy, F.R.; Lim, K.W.; Phan, A.T. Guanine anchoring: A strategy for specific targeting of a G-quadruplex using short PNA, LNA and DNA molecules. Chem. Commun. 2020, 56, 5897–5900. [Google Scholar] [CrossRef] [PubMed]
- Russo Krauss, I.; Ramaswamy, S.; Neidle, S.; Haider, S.; Parkinson, G.N. Structural Insights into the Quadruplex-Duplex 3′ Interface Formed from a Telomeric Repeat: A Potential Molecular Target. J. Am. Chem. Soc. 2016, 138, 1226–1233. [Google Scholar] [CrossRef] [Green Version]
- Asamitsu, S.; Obata, S.; Phan, A.T.; Hashiya, K.; Bando, T.; Sugiyama, H. Simultaneous Binding of Hybrid Molecules Constructed with Dual DNA-Binding Components to a G-Quadruplex and Its Proximal Duplex. Chem. A Eur. J. 2018, 24, 4428–4435. [Google Scholar] [CrossRef]
- Nguyen, T.Q.N.; Lim, K.W.; Phan, A.T. A Dual-Specific Targeting Approach Based on the Simultaneous Recognition of Duplex and Quadruplex Motifs. Sci. Rep. 2017, 7, 11969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Li, Q.; Xiang, J.; Yang, Q.; Sun, H.; Guan, A.; Wang, L.; Liu, Y.; Yu, L.; Shi, Y.; et al. Directly lighting up RNA G-quadruplexes from test tubes to living human cells. Nucleic Acids Res. 2015, 43, 9575–9586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murat, P.; Marsico, G.; Herdy, B.; Ghanbarian, A.; Portella, G.; Balasubramanian, S. RNA G-quadruplexes at upstream open reading frames cause DHX36- and DHX9-dependent translation of human mRNAs. Genome Biol. 2018, 19, 229. [Google Scholar] [CrossRef]
- Benhalevy, D.; Gupta, S.K.; Danan, C.H.; Ghosal, S.; Sun, H.-W.; Kazemier, H.G.; Paeschke, K.; Hafner, M.; Juranek, S.A. The Human CCHC-type Zinc Finger Nucleic Acid-Binding Protein Binds G-Rich Elements in Target mRNA Coding Sequences and Promotes Translation. Cell Rep. 2017, 18, 2979–2990. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.-C.; Chen, S.-B.; Dai, J.; Yuan, J.-H.; Ou, T.-M.; Huang, Z.-S.; Tan, J.-H. Tracking the Dynamic Folding and Unfolding of RNA G-Quadruplexes in Live Cells. Angew. Chemie Int. Ed. 2018, 57, 4702–4706. [Google Scholar] [CrossRef]
- Biffi, G.; Di Antonio, M.; Tannahill, D.; Balasubramanian, S. Visualization and selective chemical targeting of RNA G-quadruplex structures in the cytoplasm of human cells. Nat. Chem. 2013, 6, 75–80. [Google Scholar] [CrossRef]
- Lyu, K.; Chen, S.B.; Chan, C.Y.; Tan, J.H.; Kwok, C.K. Structural analysis and cellular visualization of APP RNA G-quadruplex. Chem. Sci. 2019, 10, 11095–11102. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.-B.; Hu, M.-H.; Liu, G.-C.; Wang, J.; Ou, T.-M.; Gu, L.-Q.; Huang, Z.-S.; Tan, J.-H. Visualization of NRAS RNA G-Quadruplex Structures in Cells with an Engineered Fluorogenic Hybridization Probe. J. Am. Chem. Soc. 2016, 138, 10382–10385. [Google Scholar] [CrossRef] [PubMed]
- Collie, G.W.; Parkinson, G.N.; Moyzis, R.K.; Buckingham, J.M.; Cram, L.S.; Dani, M.; Deaven, L.L.; Jones, M.D.; Meyne, J.; Ratliff, R.L.; et al. The application of DNA and RNA G-quadruplexes to therapeutic medicines. Chem. Soc. Rev. 2011, 40, 5867. [Google Scholar] [CrossRef] [PubMed]
- Millevoi, S.; Moine, H.; Vagner, S. G-quadruplexes in RNA biology. Wiley Interdiscip. Rev. RNA 2012, 3, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Agarwala, P.; Pandey, S.; Maiti, S. The tale of RNA G-quadruplex. Org. Biomol. Chem. 2015, 13, 5570–5585. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Bugaut, A.; Huppert, J.L.; Balasubramanian, S. An RNA G-quadruplex in the 5′ UTR of the NRAS proto-oncogene modulates translation. Nat. Chem. Biol. 2007, 3, 218–221. [Google Scholar] [CrossRef] [Green Version]
- Fay, M.M.; Lyons, S.M.; Ivanov, P. RNA G-Quadruplexes in Biology: Principles and Molecular Mechanisms. J. Mol. Biol. 2017, 429, 2127–2147. [Google Scholar] [CrossRef]
- Tian, T.; Chen, Y.Q.; Wang, S.R.; Zhou, X. G-Quadruplex: A Regulator of Gene Expression and Its Chemical Targeting. Chem 2018, 4, 1314–1344. [Google Scholar] [CrossRef] [Green Version]
- Adrian, M.; Heddi, B.; Phan, A.T. NMR spectroscopy of G-quadruplexes. Methods 2012, 57, 11–24. [Google Scholar] [CrossRef]
- Kreig, A.; Calvert, J.; Sanoica, J.; Cullum, E.; Tipanna, R.; Myong, S.; Gqs, G. G-quadruplex formation in double strand DNA probed by NMM and CV fluorescence. Nucleic Acids Res. 2015, 43, 7961–7970. [Google Scholar] [CrossRef]
- Sabharwal, N.C.; Savikhin, V.; Turek-Herman, J.R.; Nicoludis, J.M.; Szalai, V.A.; Yatsunyk, L.A. N -methylmesoporphyrin IX fluorescence as a reporter of strand orientation in guanine quadruplexes. FEBS J. 2014, 281, 1726–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kypr, J.; Kejnovska, I.; Renciuk, D.; Vorlickova, M. Circular dichroism and conformational polymorphism of DNA. Nucleic Acids Res. 2009, 37, 1713–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granqvist, L.; Virta, P. Characterization of G-Quadruplex/Hairpin Transitions of RNAs by 19 F NMR Spectroscopy. Chem. A Eur. J. 2016, 22, 15360–15372. [Google Scholar] [CrossRef]
- Kejnovská, I.; Bednářová, K.; Renčiuk, D.; Dvořáková, Z.; Školáková, P.; Trantírek, L.; Fiala, R.; Vorlíčková, M.; Sagi, J. Clustered abasic lesions profoundly change the structure and stability of human telomeric G-quadruplexes. Nucleic Acids Res. 2017, 45, 4294–4305. [Google Scholar] [CrossRef] [PubMed]
- Kejnovská, I.; Stadlbauer, P.; Trantírek, L.; Renčiuk, D.; Gajarský, M.; Krafčík, D.; Palacký, J.; Bednářová, K.; Šponer, J.; Mergny, J.-L.; et al. G-Quadruplex formation by DNA sequences deficient in guanines: Two tetrad parallel quadruplexes do not fold intramolecularly. Chem. A Eur. J. 2021. [Google Scholar] [CrossRef]
- Uddin, M.K. Phosphorylation at 5’ end of guanosine stretches inhibits dimerization of G-quadruplexes and formation of a G-quadruplex interferes with the enzymatic activities of DNA enzymes. Nucleic Acids Res. 2004, 32, 4618–4629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asamitsu, S.; Bando, T.; Sugiyama, H. Ligand Design to Acquire Specificity to Intended G-Quadruplex Structures. Chem. A Eur. J. 2019, 25, 417–430. [Google Scholar] [CrossRef]
- Tseng, T.-Y.; Chien, C.-H.; Chu, J.-F.; Huang, W.-C.; Lin, M.-Y.; Chang, C.-C.; Chang, T.-C. Fluorescent probe for visualizing guanine-quadruplex DNA by fluorescence lifetime imaging microscopy. J. Biomed. Opt. 2013, 18, 101309. [Google Scholar] [CrossRef] [Green Version]
- Tseng, T.-Y.; Chu, I.-T.; Lin, S.-J.; Li, J.; Chang, T.-C. Binding of Small Molecules to G-quadruplex DNA in Cells Revealed by Fluorescence Lifetime Imaging Microscopy of o-BMVC Foci. Molecules 2018, 24, 35. [Google Scholar] [CrossRef] [Green Version]
- Koralewska, N.; Szczepanska, A.; Ciechanowska, K.; Wojnicka, M.; Pokornowska, M.; Milewski, M.; Gudanis, D.; Baranowski, D.; Nithin, C.; Bujnicki, J.; et al. RNA and DNA G-quadruplexes bind to human Dicer and inhibit its activity. Cell. Mol. Life Sci. 2020, 1–24. [Google Scholar] [CrossRef]
- Winnerdy, F.R.; Phan, A.T. Quadruplex structure and diversity. Annu. Rep. Med. Chem. 2020, 54, 45–73. [Google Scholar] [CrossRef]
- Pandey, S.; Agarwala, P.; Maiti, S. Effect of Loops and G-Quartets on the Stability of RNA G-Quadruplexes. J. Phys. Chem. B 2013, 117, 6896–6905. [Google Scholar] [CrossRef] [PubMed]
- Fotticchia, I.; Amato, J.; Pagano, B.; Novellino, E.; Petraccone, L.; Giancola, C. How are thermodynamically stable G-quadruplex-duplex hybrids? J. Therm. Anal. Calorim. 2015, 121, 1121–1127. [Google Scholar] [CrossRef]
- Agarwala, P.; Kumar, S.; Pandey, S.; Maiti, S. Human Telomeric RNA G-Quadruplex Response to Point Mutation in the G-Quartets. J. Phys. Chem. B 2015, 119, 4617–4627. [Google Scholar] [CrossRef] [PubMed]
- Gros, J.; Rosu, F.; Amrane, S.; De Cian, A.; Gabelica, V.; Lacroix, L.; Mergny, J.-L. Guanines are a quartet’s best friend: Impact of base substitutions on the kinetics and stability of tetramolecular quadruplexes. Nucleic Acids Res. 2007, 35, 3064–3075. [Google Scholar] [CrossRef] [PubMed]
- Wieland, M.; Hartig, J.S. RNA Quadruplex-Based Modulation of Gene Expression. Chem. Biol. 2007, 14, 757–763. [Google Scholar] [CrossRef] [Green Version]
- Halder, K.; Wieland, M.; Hartig, J.S. Predictable suppression of gene expression by 5′-UTR-based RNA quadruplexes. Nucleic Acids Res. 2009, 37, 6811–6817. [Google Scholar] [CrossRef]
- He, G.-X.; Williams, J.P.; Postich, M.J.; Swaminathan, S.; Shea, R.G.; Terhorst, T.; Law, V.S.; Mao, C.T.; Sueoka, C.; Coutré, S.; et al. In Vitro and in Vivo Activities of Oligodeoxynucleotide-Based Thrombin Inhibitors Containing Neutral Formacetal Linkages. J. Med. Chem. 1998, 41, 4224–4231. [Google Scholar] [CrossRef]
- Zhang, T.; Wan, B.; Zhao, Y.; Li, C.; Liu, H.; Lv, T.; Zhan, P.; Song, Y. Treatment of uncommon EGFR mutations in non-small cell lung cancer: New evidence and treatment. Transl. Lung Cancer Res. 2019, 8, 302–316. [Google Scholar] [CrossRef]
- Asamitsu, S.; Obata, S.; Yu, Z.; Bando, T.; Sugiyama, H. Recent Progress of Targeted G-Quadruplex-Preferred Ligands Toward Cancer Therapy. Molecules 2019, 24, 429. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.; Zheng, Y.; Zhai, Q.; Wei, D. Recent advances in the development of small molecules targeting RNA G-quadruplexes for drug discovery. Bioorg. Chem. 2021, 110, 104804. [Google Scholar] [CrossRef] [PubMed]
- Armas, P.; David, A.; Calcaterra, N.B. Transcriptional control by G-quadruplexes: In vivo roles and perspectives for specific intervention. Transcription 2017, 8, 21–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.U.; Bartel, D.P. RNA G-quadruplexes are globally unfolded in eukaryotic cells and depleted in bacteria. Science 2016, 353, aaf5371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.C.; Murat, P.; Abecassis, K.; Ferre-D’Amare, A.R.; Balasubramanian, S. Insights into the mechanism of a G-quadruplex-unwinding DEAH-box helicase. Nucleic Acids Res. 2015, 43, 2223–2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | RNA G-Rich Oligonucleotide Sequences (QD) 5′-3′ | Bimolecular Complexes |
|---|---|---|
| CCC | C-GGG-C-GGG-C-CUUCAAGUCCGGCA | CCC/GT, CCC/UT |
| CCC-OMe | C-GGG-C-GGG-C-(CUUCAAGUCCGGCA)2′OMe | CCC-OMe/GT, CCC-OMe/UT |
| paa | p-GGG-a-GGG-a-CUUCAAGUCCGGCA | paa/GT, paa/UT |
| Aaa | A-GGG-a-GGG-a-CUUCAAGUCCGGCA | Aaa/GT, Aaa/UT |
| paC | p-GGG-a-GGG-C-CUUCAAGUCCGGCA | paC/GT, paC/UT |
| AaC | A-GGG-a-GGG-C-CUUCAAGUCCGGCA | AaC/GT, AaC/UT |
| pCa | p-GGG-C-GGG-a-CUUCAAGUCCGGCA | pCa/GT, pCa/UT |
| pL2C | p-GGG-L2-GGG-a-CUUCAAGUCCGGCA | pL2C/GT, pL2C/UT |
| pL3C | p-GGG-L3-GGG-a-CUUCAAGUCCGGCA | pL3C/GT, pL3C/UT |
| pL4C | p-GGG-L4-GGG-a-CUUCAAGUCCGGCA | pL4C/GT, pL4C/UT |
| DNA G-rich Oligonucleotide Sequence | ||
| DNA-CCC | C-GGG-C-GGG-C-CTTCAAGTCCGGCA | DNA-CCC/GT, DNA-CCC/UT |
| G4 Ligand Bearing RNA G-rich Oligonucleotide | ||
| L-CCC | o-BMVC-C3-aminolinker-C-GGG-C-GGG-C-CUUCAAGUCCGGCA | L-CCC/GT, L-CCC/UT |
| Target Oligonucleotide Sequences | ||
| GT | UGCCGGACUUGAAG-UA-GGGCGGGC | |
| UT | UGCCGGACUUGAAG-UA-GGGCUGGC | |
| Control Oligonucleotide Sequences | ||
| DX | UGCCGGACUUGAAG/CUUCAAGUCCGGCA | |
| 2Q | CGGGCGGGC |
| QD/GT | 60 mM K+ | QD/UT | 60 mM K+ |
|---|---|---|---|
| Tm (°C) | Tm (°C) | ||
| CCC/GT | 69.4 | CCC/UT | 61.0 |
| CCC-OMe/GT | 71.7 | CCC-OMe/UT | 63.5 |
| paa/GT | 68.0 | paa/UT | 61.5 |
| Aaa/GT | 69.0 | Aaa/UT | 61.0 |
| paC/GT | 67.5 | paC/GT | 61.0 |
| AaC/GT | 68.5 | AaC/GT | 62.0 |
| pCa/GT | 67.4 | pCa/GT | 60.5 |
| pL2C/GT | 68.0 | pL2C/GT | 61.5 |
| pL3C/GT | 68.5 | pL3C/GT | 61.5 |
| pL4C/GT | 68.5 | pL4C/GT | 60.5 |
| DNA-CCC/GT | 61.4 | DNA-CCC/UT | 59.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gudanis, D.; Zielińska, K.; Baranowski, D.; Kierzek, R.; Kozłowski, P.; Gdaniec, Z. Impact of a Single Nucleotide Change or Non-Nucleoside Modifications in G-Rich Region on the Quadruplex–Duplex Hybrid Formation. Biomolecules 2021, 11, 1236. https://doi.org/10.3390/biom11081236
Gudanis D, Zielińska K, Baranowski D, Kierzek R, Kozłowski P, Gdaniec Z. Impact of a Single Nucleotide Change or Non-Nucleoside Modifications in G-Rich Region on the Quadruplex–Duplex Hybrid Formation. Biomolecules. 2021; 11(8):1236. https://doi.org/10.3390/biom11081236
Chicago/Turabian StyleGudanis, Dorota, Karolina Zielińska, Daniel Baranowski, Ryszard Kierzek, Piotr Kozłowski, and Zofia Gdaniec. 2021. "Impact of a Single Nucleotide Change or Non-Nucleoside Modifications in G-Rich Region on the Quadruplex–Duplex Hybrid Formation" Biomolecules 11, no. 8: 1236. https://doi.org/10.3390/biom11081236
APA StyleGudanis, D., Zielińska, K., Baranowski, D., Kierzek, R., Kozłowski, P., & Gdaniec, Z. (2021). Impact of a Single Nucleotide Change or Non-Nucleoside Modifications in G-Rich Region on the Quadruplex–Duplex Hybrid Formation. Biomolecules, 11(8), 1236. https://doi.org/10.3390/biom11081236