
The Role of Polymorphisms in Collagen-Encoding Genes in Intervertebral Disc Degeneration

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. COL1A1 Gene
3.2. COL1A2 Gene
3.3. COL2A1 Gene
3.4. COL9A1 Gene
3.5. COL9A2 Gene
3.6. COL9A3 Gene
3.7. COL11A1 Gene
3.8. COL11A2 Gene
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Munir, S.; Rade, M.; Määttä, J.H.; Freidin, M.B.; Williams, F.M.K. Intervertebral Disc Biology: Genetic Basis of Disc Degeneration. Curr. Mol. Biol. Rep. 2018, 4, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motina, A.N.; Astaschenko, Y.A.; Masaleva, I.O.; Tretyakova, E.E. The social hygienic characteristic of patients with osteochondrosis of spine. Probl. Sotsial’noi Gig. Zdr. Istor. Meditsiny 2020, 28, 396–399. (In Russian) [Google Scholar] [CrossRef] [PubMed]
- Dagenais, S.; Caro, J.; Haldeman, S.A. Systematic review of low back pain cost of illness studies in the United States and internationally. Spine J. 2008, 8, 8–20. [Google Scholar] [CrossRef]
- Byvaltsev, V.A.; Kalinin, A.A.; Okoneshnikova, A.K.; Irintseev, A.A. Differentiated surgical tactics in degenerative diseases of lumbar spine department with the use of functional methods. Sib. Med. Rev. 2018, 5, 54–65. [Google Scholar] [CrossRef]
- Melnikova, E.V.; Popov, A.P. Venlafaxine in the treatment of chronic pain syndromes. V.M. Bekhterev Rev. Psychiatry Med. Psychol. 2010, 4, 55–58. (In Russian) [Google Scholar]
- Schmidt, I.R. Solved and unsolved problems of vertebral neurology at the present stage of development of science. Med. Kuzbasse 2004, 3, 13–17. (In Russian) [Google Scholar]
- Feng, Y.; Egan, B.; Wang, J. Genetic Factors in Intervertebral Disc Degeneration. Genes Dis. 2016, 3, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Hanaei, S.; Abdollahzade, S.; Khoshnevisan, A.; Kepler, C.K.; Rezaei, N. Genetic aspects of intervertebral disc degeneration. Rev. Neurosci. 2015, 26, 581–606. [Google Scholar] [CrossRef]
- Kitis, S.; Coskun, Z.M.; Tasdemir, P.; Tuncez, E.; Zamani, A.G.; Acar, A. Analysis of genetic polymorphisms associated with intervertebral disc degeneration. Cell. Mol. Biol. 2018, 64, 61–65. [Google Scholar] [CrossRef]
- Vieira, L.A.; Dos Santos, A.A.; Peluso, C.; Barbosa, C.P.; Bianco, B.; Rodrigues, L.M.R. Influence of lifestyle characteristics and VDR polymorphisms as risk factors for intervertebral disc degeneration: A case-control study. Eur. J. Med. Res. 2018, 23, 11. [Google Scholar] [CrossRef] [Green Version]
- Seo, B.R.; Chen, X.; Ling, L.; Song, Y.H.; Shimpi, A.A.; Choi, S.; Gonzalez, J.; Sapudom, J.; Wang, K.; Andresen Eguiluz, R.C.; et al. Collagen microarchitecture mechanically controls myofibroblast differentiation. Proc. Natl. Acad. Sci. USA 2020, 117, 11387–11398. [Google Scholar] [CrossRef] [PubMed]
- Mostaço-Guidolin, L.; Rosin, N.L.; Hackett, T.L. Imaging Collagen in Scar Tissue: Developments in Second Harmonic Generation Microscopy for Biomedical Applications. Int. J. Mol. Sci. 2017, 18, 1772. [Google Scholar] [CrossRef] [PubMed]
- de Wild, M.; Pomp, W.; Koenderink, G.H. Thermal memory in self-assembled collagen fibril networks. Biophys. J. 2013, 105, 200–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzi, A.; Storelli, E.; Bettini, S.; Sibillano, T.; Altamura, D.; Salvatore, L.; Madaghiele, M.; Romano, A.; Siliqi, D.; Ladisa, M.; et al. Effects of processing on structural, mechanical and biological properties of collagen-based substrates for regenerative medicine. Sci. Rep. 2018, 8, 1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, M.C.; Yumnam, S.; Kim, S.Y. Oral Intake of Collagen Peptide Attenuates Ultraviolet B Irradiation-Induced Skin Dehydration In Vivo by Regulating Hyaluronic Acid Synthesis. Int. J. Mol. Sci. 2018, 19, 3551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeldin, L.; Mosley, G.E.; Laudier, D.; Gallate, Z.S.; Gansau, J.; Hoy, R.C.; Poeran, J.; Iatridis, J.C. Spatial mapping of collagen content and structure in human intervertebral disk degeneration. JOR Spine 2020, 3, 1129. [Google Scholar] [CrossRef]
- Yakovleva, N.V.; Shavkuta, G.V.; Shnyukova, T.V. The effectiveness of chondroprotective therapy in patients with lumbosacral dorsopathies. Lechashchij Vrach 2020, 12, 61–65. (In Russian) [Google Scholar] [CrossRef]
- Sarrigiannidis, S.O.; Rey, J.M.; Dobre, O.; González-García, C.; Dalby, M.J.; Salmeron-Sanchez, M. A tough act to follow: Collagen hydrogel modifications to improve mechanical and growth factor loading capabilities. Mater. Today Bio 2021, 10, 100098. [Google Scholar] [CrossRef]
- Manka, S.W.; Bihan, D.; Farndale, R.W. Structural studies of the MMP-3 interaction with triple-helical collagen introduce new roles for the enzyme in tissue remodeling. Sci. Rep. 2019, 9, 18785. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.Z.; Rui, Y.F.; Lu, J.; Wang, C. Cell and molecular biology of intervertebral disc degeneration: Current understanding and implications for potential therapeutic strategies. Cell Prolif. 2014, 47, 381–390. [Google Scholar] [CrossRef]
- NCBI. Genes & Expression. Gene. Available online: https://www.ncbi.nlm.nih.gov/gene/ (accessed on 23 April 2021).
- Eyre, D.R.; Muir, H. Quantitative analysis of types I and II collagens in human intervertebral discs at various ages. Biochim. Biophys. Acta 1977, 492, 29–42. [Google Scholar] [CrossRef]
- Henriksen, K.; Karsdal, M.A. Type I collagen. In Biochemistry of Collagens, Laminins and Elastin; Academic Press: Cambridge, MA, USA, 2016; pp. 1–11. [Google Scholar] [CrossRef]
- Rong, Y.; Sugumaran, G.; Silbert, J.E.; Spector, M. Proteoglycans synthesized by canine intervertebral disc cells grown in a type I collagen-glycosaminoglycan matrix. Tissue Eng. 2002, 8, 1037–1047. [Google Scholar] [CrossRef] [PubMed]
- Sarver, J.J.; Elliott, D.M. Altered disc mechanics in mice genetically engineered for reduced type I collagen. Spine 2004, 29, 1094–1098. [Google Scholar] [CrossRef] [PubMed]
- Anjankar, S.D.; Poornima, S.; Raju, S.; Jaleel, M.A.; Bhiladvala, D.; Hasan, Q. Degenerated intervertebral disc prolapse and its association of collagen I alpha 1 Spl gene polymorphism: A preliminary case control study of Indian population. Indian J. Orthop. 2015, 49, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Genetic Home Reference, Your Guide to Undersatanding Genetic Conditions COL1A1. 2012. Available online: http://www.ghr.nlm.nih.gov/gene/COL1A1 (accessed on 14 April 2021).
- Pluijm, S.M.; van Essen, H.W.; Bravenboer, N.; Uitterlinden, A.G.; Smit, J.H.; Pols, H.A.; Lips, P. Collagen type I alpha1 Sp1 polymorphism, osteoporosis, and intervertebral disc degeneration in older men and women. Ann. Rheum Dis. 2004, 63, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Tilkeridis, C.; Bei, T.; Garantziotis, S.; Stratakis, C.A. Association of a COL1A1 polymorphism with lumbar disc disease in young military recruits. J. Med. Genet. 2005, 42, 44. [Google Scholar] [CrossRef] [Green Version]
- Videman, T.; Saarela, J.; Kaprio, J.; Näkki, A.; Levälahti, E.; Gill, K.; Peltonen, L.; Battié, M.C. Associations of 25 structural, degradative, and inflammatory candidate genes with lumbar disc desiccation, bulging, and height narrowing. Arthritis Rheum. 2009, 60, 470–481. [Google Scholar] [CrossRef]
- Zhong, B.; Huang, D.; Ma, K.; Deng, X.; Shi, D.; Wu, F.; Shao, Z. Association of COL1A1 rs1800012 polymorphism with musculoskeletal degenerative diseases: A meta-analysis. Oncotarget 2017, 8, 75488–75499. [Google Scholar] [CrossRef] [Green Version]
- Hanaei, S.; Abdollahzade, S.; Sadr, M.; Fattahi, E.; Mirbolouk, M.H.; Khoshnevisan, A.; Rezaei, N. Lack of association between COL1A1 and COL9A2 single nucleotide polymorphisms and intervertebral disc degeneration. Br. J. Neurosurg. 2021, 35, 77–79. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, Z.; Liu, J.; Guo, X. Advances in susceptibility genetics of intervertebral degenerative disc disease. Int. J. Biol. Sci. 2008, 4, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Antoniou, J.; Steffen, T.; Nelson, F.; Winterbottom, N.; Hollander, A.P.; Poole, R.A.; Aebi, M.; Alini, M. The human lumbar intervertebral disc. Evidence for changes in the biosynthesis and denaturation of the extracellular matrix with growth, maturation, ageing, and degeneration. J. Clin. Investig. 1996, 98, 996–1003. [Google Scholar] [CrossRef]
- Bedore, J.; Quesnel, K.; Quinonez, D.; Séguin, C.A.; Leask, A. Targeting the annulus fibrosus of the intervertebral disc: Col1a2-Cre(ER)T mice show specific activity of Cre recombinase in the outer annulus fibrosus. J. Cell Commun. Signal. 2016, 10, 137–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Wu, D.; Zhu, X.; Ni, H.; Wei, X.; Mao, N.; Xie, Y.; Niu, Y.; Li, M. Gene expression profile analysis of human intervertebral disc degeneration. Genet. Mol. Biol. 2013, 36, 448–454. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Fu, Y.; Yan, B.; Shen, Z.; Lan, T. Analysis of key genes and pathways associated with the pathogenesis of intervertebral disc degeneration. J. Orthop. Surg. Res. 2020, 15, 371. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, G.; Jiao, W.; Wang, D.; Cao, Y.; Zhang, Q.; Wang, J. Identification of critical genes in nucleus pulposus cells isolated from degenerated intervertebral discs using bioinformatics analysis. Mol. Med. Rep. 2017, 16, 553–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponticos, M.; Abraham, D.; Alexakis, C.; Lu, Q.L.; Black, C.; Partridge, T.; Bou-Gharios, G. Col1a2 enhancer regulates collagen activity during development and in adult tissue repair. Matrix Biol. 2004, 22, 619–628. [Google Scholar] [CrossRef]
- Mayer, J.E.; Iatridis, J.C.; Chan, D.; Qureshi, S.A.; Gottesman, O.; Hecht, A.C. Genetic polymorphisms associated with intervertebral disc degeneration. Spine J. 2013, 13, 299–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gudmann, N.S.; Karsdal, M.A. Type II collagen. In Biochemistry of Collagens, Laminins and Elastin; Academic Press: Cambridge, MA, USA, 2016; pp. 13–20. [Google Scholar] [CrossRef]
- Li, Y.Z.; Li, J.; Zhang, J.; Lin, Q. Association of COL2A and Aggrecan polymorphisms with the susceptibility of intervertebral disc degeneration. Int. J. Clin. Exp. Med. 2016, 9, 3885–3892. [Google Scholar]
- Deng, Y.; Tan, X.T.; Wu, Q.; Wang, X. Correlations Between COL2A and Aggrecan Genetic Polymorphisms and the Risk and Clinicopathological Features of Intervertebral Disc Degeneration in a Chinese Han Population: A Case-Control Study. Genet. Test. Mol. Biomark. 2017, 21, 108–115. [Google Scholar] [CrossRef]
- Huang, D.; Deng, X.; Ma, K.; Wu, F.; Shi, D.; Liang, H.; Chen, S.; Shao, Z. Association of COL9A3 trp3 polymorphism with intervertebral disk degeneration: A meta-analysis. BMC Musculoskelet. Disord. 2018, 19, 381. [Google Scholar] [CrossRef] [PubMed]
- Bächinger, H.P.; Mizuno, K.; Vranka, J.A.; Boudko, S.P. Collagen Formation and Structure. In Comprehensive Natural Products II: Chemistry and Biology; Elsevier Ltd.: Amsterdam, The Netherlands, 2010; pp. 469–530. ISBN 978-0-08-045382-8. [Google Scholar] [CrossRef]
- He, Y.; Karsdal, M.A. Type IX collagen. In Biochemistry of Collagens, Laminins and Elastin; Academic Press: Cambridge, MA, USA, 2016; pp. 67–71, 77–80. [Google Scholar]
- Nakata, K.; Ono, K.; Miyazaki, J.; Olsen, B.R.; Muragaki, Y.; Adachi, E.; Yamamura, K.; Kimura, T. Osteoarthritis associated with mild chondrodysplasia in transgenic mice expressing alpha 1(IX) collagen chains with a central deletion. Proc. Natl. Acad. Sci. USA 1993, 90, 2870–2874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, T.; Nakata, K.; Tsumaki, N.; Miyamoto, S.; Matsui, Y.; Ebara, S.; Ochi, T. Progressive degeneration of articular cartilage and intervertebral discs. An experimental study in transgenic mice bearing a type IX collagen mutation. Int. Orthop. 1996, 20, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Zielinska, N.; Podgórski, M.; Haładaj, R.; Polguj, M.; Olewnik, L. Risk Factors of Intervertebral Disc Pathology—A Point of View Formerly and Today—A Review. J. Clin. Med. 2021, 10, 409. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.J.; Eyre, D.R. Intervertebral disc collagen. Usage of the short form of the alpha1(IX) chain in bovine nucleus pulposus. J. Biol. Chem. 2003, 278, 24521–245355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eyre, D.R.; Matsui, Y.; Wu, J.J. Collagen polymorphisms of the intervertebral disc. Biochem. Soc. Trans. 2002, 30, 844–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathod, T.N.; Chandanwale, A.S.; Gujrathi, S.; Patil, V.; Chavan, S.A.; Shah, M.N. Association between single nucleotide polymorphism in collagen IX and intervertebral disc disease in the Indian population. Indian J. Orthop. 2012, 46, 420–426. [Google Scholar] [CrossRef]
- Annunen, S.; Paassilta, P.; Lohiniva, J.; Perälä, M.; Pihlajamaa, T.; Karppinen, J.; Tervonen, O.; Kröger, H.; Lähde, S.; Vanharanta, H.; et al. An allele of COL9A2 associated with intervertebral disc disease. Science 1999, 285, 409–412. [Google Scholar] [CrossRef]
- Karppinen, J.; Paakko, E.; Raina, S.; Tervonen, O.; Kurunlahti, M.; Nieminen, P.; Ala-Kokko, L.; Malmivaara, A.; Vanharanta, H. Magnetic resonance imaging findings in relation to the COL9A2 tryptophan allele among patients with sciatica. Spine 2002, 27, 78–83. [Google Scholar] [CrossRef]
- Wrocklage, C.; Wassmann, H.; Paulus, W. COL9A2 allelotypes in intervertebral disc disease. Biochem. Biophys. Res. Commun. 2000, 279, 398–400. [Google Scholar] [CrossRef]
- Seki, S.; Kawaguchi, Y.; Mori, M.; Mio, F.; Chiba, K.; Mikami, Y.; Ikegawa, S. Association study of COL9A2 with lumbar disc disease in the Japanese population. J. Hum. Genet. 2006, 51, 1063–1067. [Google Scholar] [CrossRef]
- Feng, H.; Danfelter, M.; Stromqvist, B.; Heinegard, D. Extracellular matrix in disc degeneration. J. Bone Jt. Surg. Am. 2006, 88, 25–29. [Google Scholar]
- Jim, J.J.; Noponen-Hietala, N.; Cheung, K.M.; Ott, J.; Karppinen, J.; Sahraravand, A.; Luk, K.D.; Yip, S.P.; Sham, P.C.; Song, Y.Q.; et al. The TRP2 allele of COL9A2 is an age-dependent risk factor for the development and severity of intervertebral disc degeneration. Spine 2005, 30, 2735–2742. [Google Scholar] [CrossRef]
- Wu, H.; Wang, S.; Chen, W.; Zhan, X.; Xiao, Z.; Jiang, H.; Wei, Q.; Wu, H.; Wang, S.; Chen, W.; et al. Collagen IX gene polymorphisms and lumbar disc degeneration: A systematic review and meta-analysis. J. Orthop. Surg. Res. 2018, 5, 47. [Google Scholar] [CrossRef] [PubMed]
- Janeczko, Ł.; Janeczko, M.; Chrzanowski, R.; Zieliński, G. The role of polymorphisms of genes encoding collagen IX and XI in lumbar disc disease. Neurol. Neurochir. Polska 2014, 48, 60–62. [Google Scholar] [CrossRef] [PubMed]
- Mio, F.; Chiba, K.; Hirose, Y.; Kawaguchi, Y.; Mikami, Y.; Oya, T.; Mori, M.; Kamata, M.; Matsumoto, M.; Ozaki, K.; et al. A functional polymorphism in COL11A1, which encodes the alpha 1 chain of type XI collagen, is associated with susceptibility to lumbar disc herniation. Am. J. Hum. Genet. 2007, 81, 1271–1277. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Sun, G.; Guo, L.; Wang, L.; Fan, W.; Lang, M.; Chen, D.; Yi, X. A genetic variant in COL11A1 is functionally associated with lumbar disc herniation in Chinese population. J. Genet. 2017, 96, 867–872. [Google Scholar] [CrossRef]
- Jiang, H.; Yang, Q.; Jiang, J.; Zhan, X.; Xiao, Z. Association between COL11A1 (rs1337185) and ADAMTS5 (rs162509) gene polymorphisms and lumbar spine pathologies in Chinese Han population: An observational study. BMJ Open 2017, 7, e015644. [Google Scholar] [CrossRef] [Green Version]
- Rajasekaran, S.; Kanna, R.M.; Senthil, N.; Raveendran, M.; Ranjani, V.; Cheung, K.M.; Chan, D.; Kao, P.Y.; Yee, A.; Shetty, A.P. Genetic susceptibility of lumbar degenerative disc disease in young Indian adults. Eur Spine J. 2015, 24, 1969–1975. [Google Scholar] [CrossRef]
- Melkoniemi, M.; Brunner, H.G.; Manouvrier, S.; Hennekam, R.; Superti-Furga, A.; Kääriäinen, H.; Pauli, R.M.; van Essen, T.; Warman, M.L.; Bonaventure, J.; et al. Autosomal recessive disorder otospondylomegaepiphyseal dysplasia is associated with loss-of-function mutations in the COL11A2 gene. Am. J. Hum. Genet. 2000, 66, 368–377. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, E.A.; Kague, E.; Aggleton, J.A.; Harniman, R.L.; Roddy, K.A.; Hammond, C.L. The mechanical impact of col11a2 loss on joints; col11a2 mutant zebrafish show changes to joint development and function, which leads to early-onset osteoarthritis. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.; Jiang, X.; Lu, J.; Wang, K.; Sun, Y.; Zhang, Y. Genetic variant of COL11A2 gene is functionally associated with developmental dysplasia of the hip in Chinese Han population. Aging 2020, 12, 7694–7703. [Google Scholar] [CrossRef]
- Caldeira, J.; Santa, C.; Osório, H.; Molinos, M.; Manadas, B.; Gonçalves, R.; Barbosa, M. Matrisome Profiling During Intervertebral Disc Development and Ageing. Sci. Rep. 2017, 7, 11629. [Google Scholar] [CrossRef] [PubMed]
- Noponenhietala, N.; Kyllönen, E.; Männikkö, M.; Ilkko, E.; Karppinen, J.; Ott, J.; Ala-Kokko, L. Sequence variations in the collagen IX and XI genes are associated with degenerative lumbar spinal stenosis. Ann. Rheum. Dis. 2003, 62, 1208. [Google Scholar] [CrossRef]
- Virtanen, I.M.; Karppinen, J.; Taimela, S.; Ott, J.; Barral, S.; Kaikkonen, K.; Ala-Kokko, L. Occupational and genetic risk factors associated with intervertebral disc disease. Spine 2007, 32, 1129. [Google Scholar] [CrossRef] [PubMed]
- Solovieva, S.; Lohiniva, J.; Leino-Arjas, P.; Raininko, R.; Luoma, K.; Ala-Kokko, L.; Riihimäki, H. Intervertebral disc degeneration in relation to the COL9A3 and the IL-1ss gene polymorphisms. Eur. Spine J. 2006, 15, 613–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Jia, H.; Xing, W.; Li, F.; Li, M.; Sun, K.; Zhu, Y. Genetic variants in COL11A2 of lumbar disk degeneration among Chinese Han population. Mol. Genet. Genom. Med. 2019, 7, 00524. [Google Scholar] [CrossRef]
- Buckwalter, J.A. Aging and degeneration of the human intervertebral disc. Spine 1995, 20, 1307–1314. [Google Scholar] [CrossRef]
- Eskola, P.J.; Kjaer, P.; Daavittila, I.M.; Solovieva, S.; Okuloff, A.; Sorensen, J.S.; Karppinen, J.I. Genetic risk factors of disc degeneration among 12-14-year-old Danish children: A population study. Int. J. Mol. Epidemiol. Genet. 2010, 1, 158–165. [Google Scholar]
- Zheng, C.J.; Chen, J. Disc degeneration implies low back pain. Theor. Biol. Med. Model. 2015, 12, 24. [Google Scholar] [CrossRef] [Green Version]
- Brinjikji, W.; Diehn, F.E.; Jarvik, J.G.; Carr, C.M.; Kallmes, D.F.; Murad, M.H.; Luetmer, P.H. MRI Findings of Disc Degeneration are More Prevalent in Adults with Low Back Pain than in Asymptomatic Controls: A Systematic Review and Meta-Analysis. AJNR Am. J. Neuroradiol. 2015, 36, 2394–2399. [Google Scholar] [CrossRef] [Green Version]
- Kadow, T.; Sowa, G.; Vo, N.; Kang, J.D. Molecular basis of intervertebral disc degeneration and herniations: What are the important translational questions? Clin. Orthop. Relat. Res. 2015, 473, 1903–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalb, S.; Martirosyan, N.L.; Kalani, M.Y.; Broc, G.G.; Theodore, N. Genetics of the degenerated intervertebral disc. World Neurosurg. 2012, 77, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Hemanta, D.; Jiang, X.X.; Feng, Z.Z.; Chen, Z.X.; Cao, Y.W. Etiology for Degenerative Disc Disease. Chin. Med. Sci. J. 2016, 31, 185–191. [Google Scholar] [CrossRef]
- Trefilova, V.V.; Shnayder, N.A.; Popova, T.E.; Balberova, O.V.; Nasyrova, R.F. The role of NO system in low back pain chronicity. Pers. Psychiatry Neurol. 2021, 1, 37–45. [Google Scholar] [CrossRef]
- Cornetta, K.; Brown, C.G. Balancing personalized medicine and personalized care. Acad. Med. 2013, 88, 309–313. [Google Scholar] [CrossRef] [Green Version]
- Neznanov, N.G. A paradigm shift to treat psychoneurological disorders. Pers. Psychiatry Neurol. 2021, 1, 1–2. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
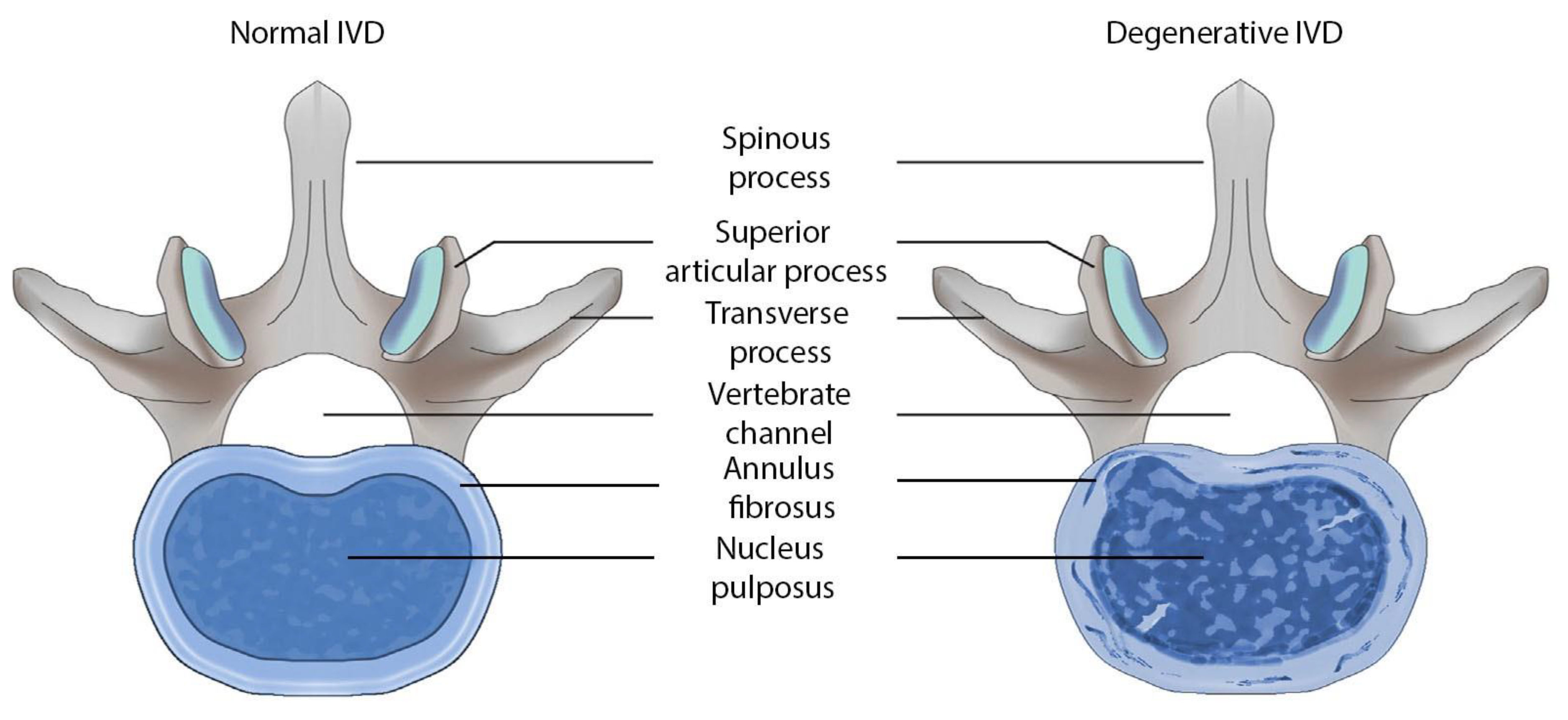
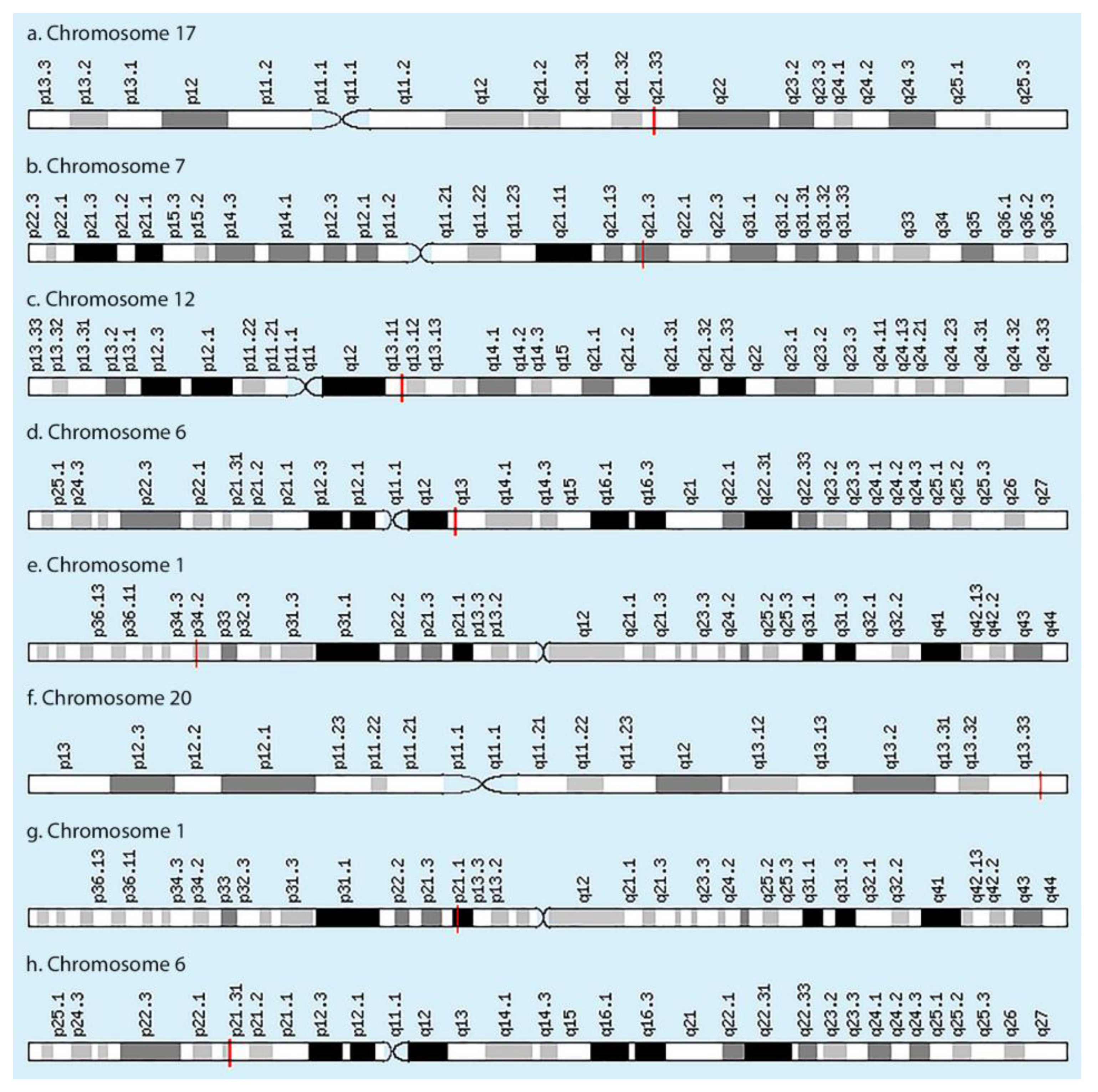
| Gene and Protein/Enzyme Encoded by This Gene | Chromosome Localization | Clinical Manifestations of Mutation/Polymorphism |
|---|---|---|
| COL1A1, encoding the α1 chain of collagen type I | 17q21.33, 51 exons | Osteogenesis imperfecta Classic type of Ehlers-Danlos syndrome Caffey disease Idiopathic osteoporosis |
| COL1A2, encoding α2 chain of collagen type I chain | 17q21.3, 52 exons | Osteogenesis imperfecta Ehlers-Danlos syndrome type VII B Idiopathic osteoporosis Atypical Marfan syndrome |
| COL2A1, encoding the α1 chain of collagen type II | 12q13.11, 57 exons | Achondrogenesis Chondrodysplasia Early onset familial osteoarthritis SED congenita Langer-Saldino achondrogenesis Kniest dysplasia Stickler syndrome type I Spondyloepimetaphyseal dysplasia Strudwick type |
| COL9A1, encoding the α1 chain of collagen type IX | 6q13, 43 exons | Osteoarthritis with early onset with multiple epiphyseal dysplasia Chondrodysplasia type Vl Stickler syndrome |
| COL9A2, encoding the α2 chain of collagen type IX | 1p34.2, 35 exons | Multiple epiphyseal dysplasia |
| COL9A3, encoding the α3 chain of collagen type IX | 20q13.33, 33 exons | Multiple epiphyseal dysplasia type III |
| COL11A1, encoding the α1 chain of collagen type XI | 1p21.1, 71 exons | Stickler syndrome type II Marshall syndrome Susceptibility to lumbar disc herniation |
| COL11A2, encoding the α2 chain of collagen type XI | 6p21.32, 70 exons | Stickler syndrome type III Otospondylomegaepiphyseal dysplasia (OSMED syndrome) Weissenbacher–Zweymuller syndrome Autosomal dominant non-syndromic sensorineural type 13 deafness (DFNA13) Autosomal recessive non-syndromic sensorineural type 53 deafness (DFNB53) |
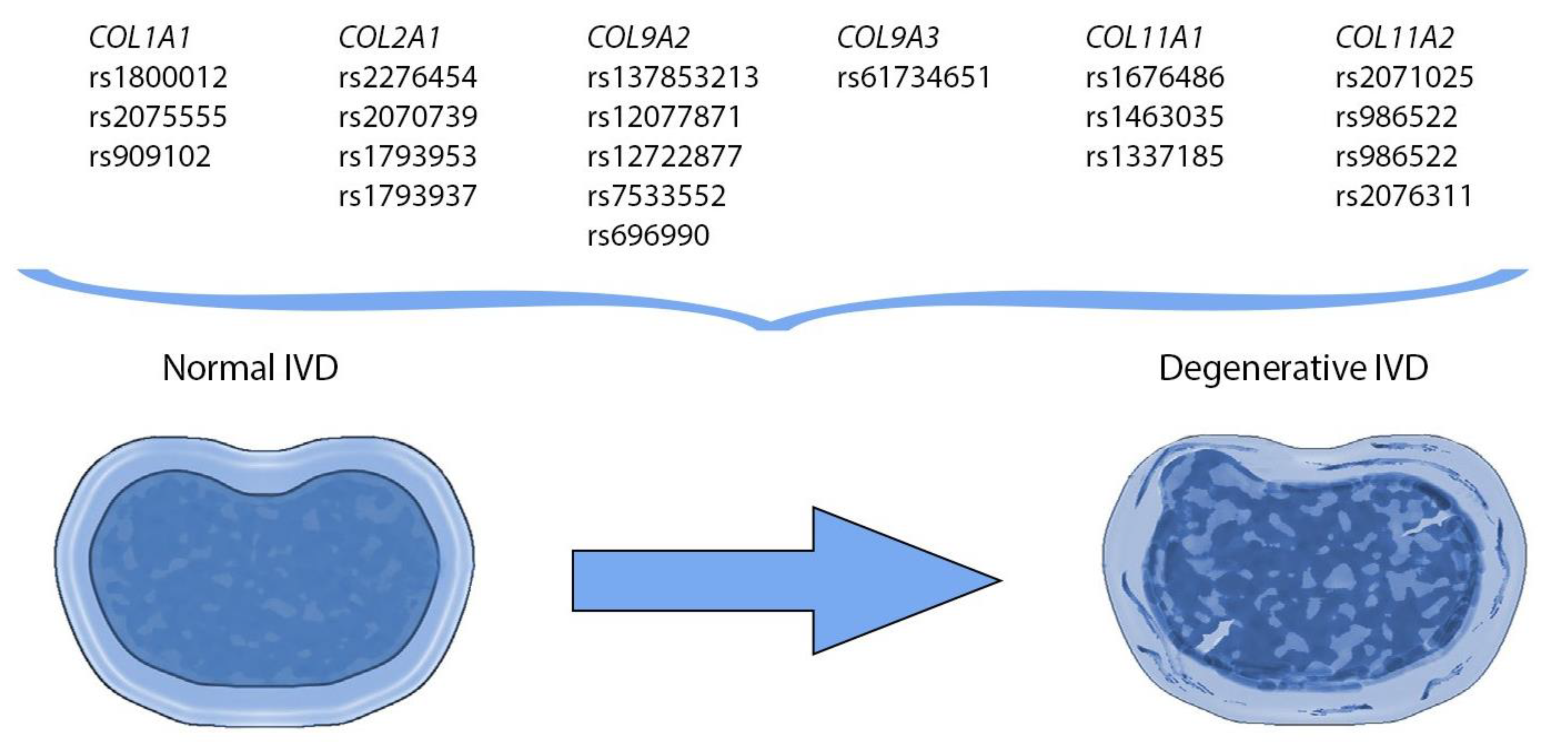
| Gene, Chromosomal Locus | Single Nucleotide Variants | Protein | Source |
|---|---|---|---|
| COL1A1 17q21.33 | rs1800012 rs2075555 rs909102 | Alpha 1 chain of collagen type I | [7,26,28,29,30,31,32] |
| COL1A2 7q21.3 | n/a * | Alpha 2 chain of collagen type I | [33,36] |
| COL2A1 12q13.11 | rs2276454 rs2070739 rs1793953 rs1793937 | Alpha 1 chain of collagen type II | [1,42,43]. |
| COL9A1 6q13 | n/a * | Alpha 1 chain of collagen type IX | [7,49] |
| COL9A2 1p34.2 | rs137853213 rs12077871 rs12722877 rs7533552 rs696990 | Alpha 2 chain of collagen type IX | [7,44,49,53] |
| COL9A3 20q13.33 | rs61734651 | Alpha 3 chain of collagen type IX | [44,49,59] |
| COL11A1 1p21.1 | rs1676486 rs1463035 rs1337185 | Alpha 1 chain of collagen type XI | [7,30,49,61,62] |
| COL11A2 6p21.32 | rs2071025 rs986522 rs986522 rs2076311 | Alpha 2 chain of collagen type XI | [7,30,71,72] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trefilova, V.V.; Shnayder, N.A.; Petrova, M.M.; Kaskaeva, D.S.; Tutynina, O.V.; Petrov, K.V.; Popova, T.E.; Balberova, O.V.; Medvedev, G.V.; Nasyrova, R.F. The Role of Polymorphisms in Collagen-Encoding Genes in Intervertebral Disc Degeneration. Biomolecules 2021, 11, 1279. https://doi.org/10.3390/biom11091279
Trefilova VV, Shnayder NA, Petrova MM, Kaskaeva DS, Tutynina OV, Petrov KV, Popova TE, Balberova OV, Medvedev GV, Nasyrova RF. The Role of Polymorphisms in Collagen-Encoding Genes in Intervertebral Disc Degeneration. Biomolecules. 2021; 11(9):1279. https://doi.org/10.3390/biom11091279
Chicago/Turabian StyleTrefilova, Vera V., Natalia A. Shnayder, Marina M. Petrova, Daria S. Kaskaeva, Olga V. Tutynina, Kirill V. Petrov, Tatiana E. Popova, Olga V. Balberova, German V. Medvedev, and Regina F. Nasyrova. 2021. "The Role of Polymorphisms in Collagen-Encoding Genes in Intervertebral Disc Degeneration" Biomolecules 11, no. 9: 1279. https://doi.org/10.3390/biom11091279
APA StyleTrefilova, V. V., Shnayder, N. A., Petrova, M. M., Kaskaeva, D. S., Tutynina, O. V., Petrov, K. V., Popova, T. E., Balberova, O. V., Medvedev, G. V., & Nasyrova, R. F. (2021). The Role of Polymorphisms in Collagen-Encoding Genes in Intervertebral Disc Degeneration. Biomolecules, 11(9), 1279. https://doi.org/10.3390/biom11091279