
Elevated Expression of JMJD5 Protein Due to Decreased miR-3656 Levels Contributes to Cancer Stem Cell-Like Phenotypes under Overexpression of Cancer Upregulated Gene 2

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Antibodies and Transfection
2.3. Immunoblotting
2.4. Immunofluorescence Microscopy
2.5. 3′ Untranslated Region (UTR) Reporter Assay
2.6. Wound Healing Assay
2.7. Transwell Invasion Assay
2.8. Sphere Forming Assay
2.9. Measurement of Reactive Oxygen Species
2.10. Statistical Analysis
3. Results
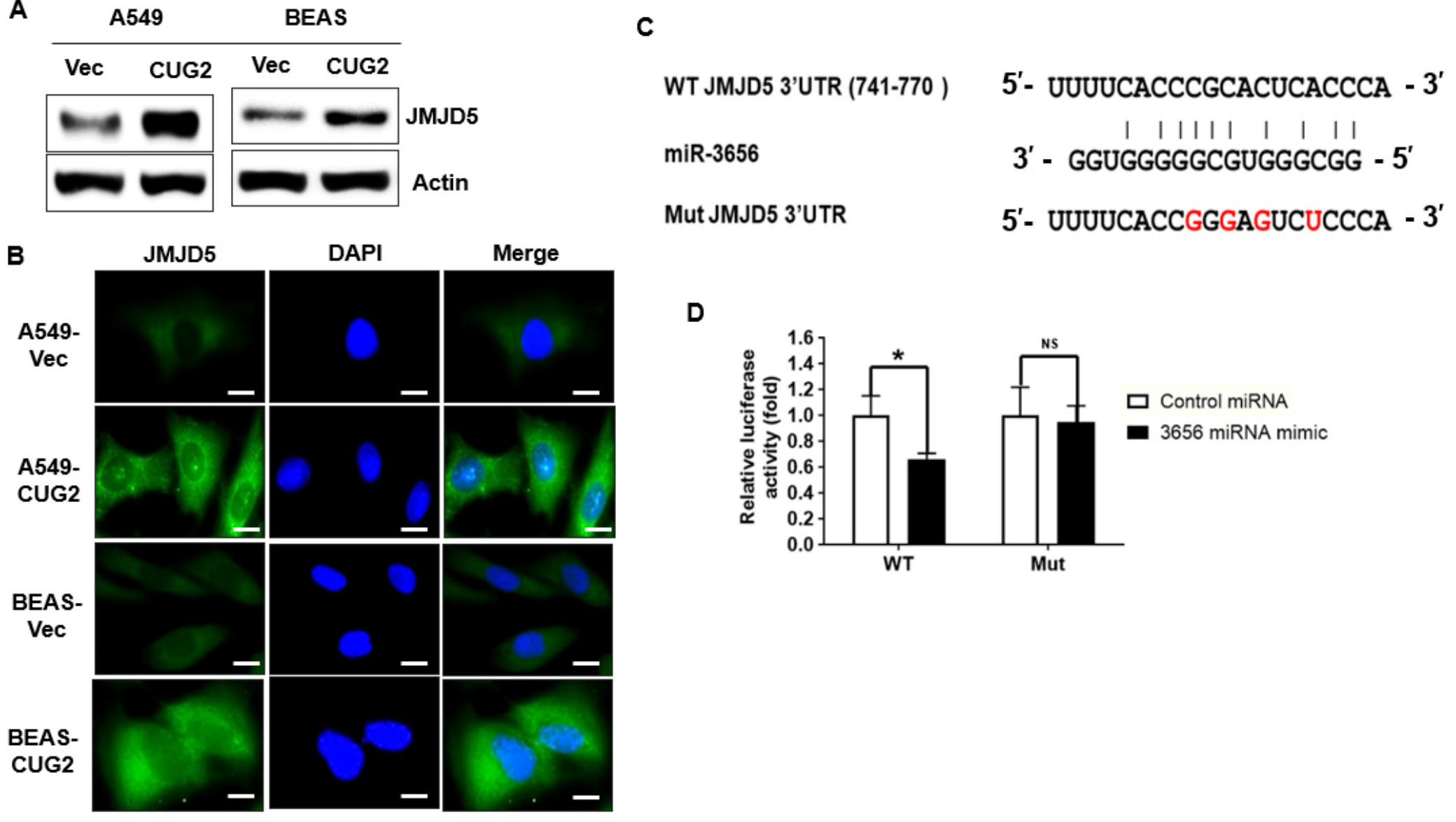
3.1. JMJD5 Is a Functional Target of miR-3656 under CUG2 Overexpression
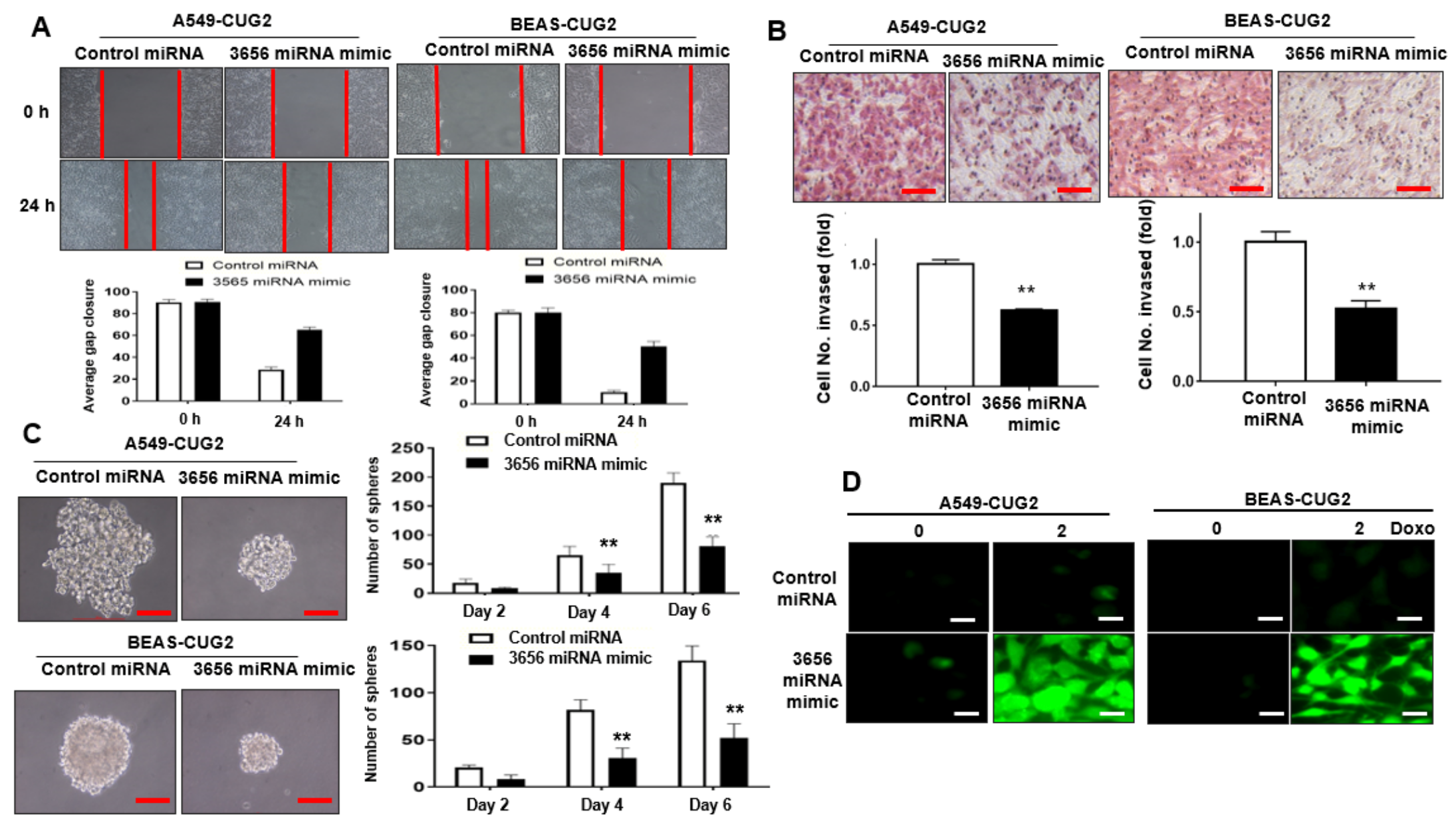
3.2. Administration of miR-3656 Mimic Hinders CUG2-Induced CSC-Like Phenotypes
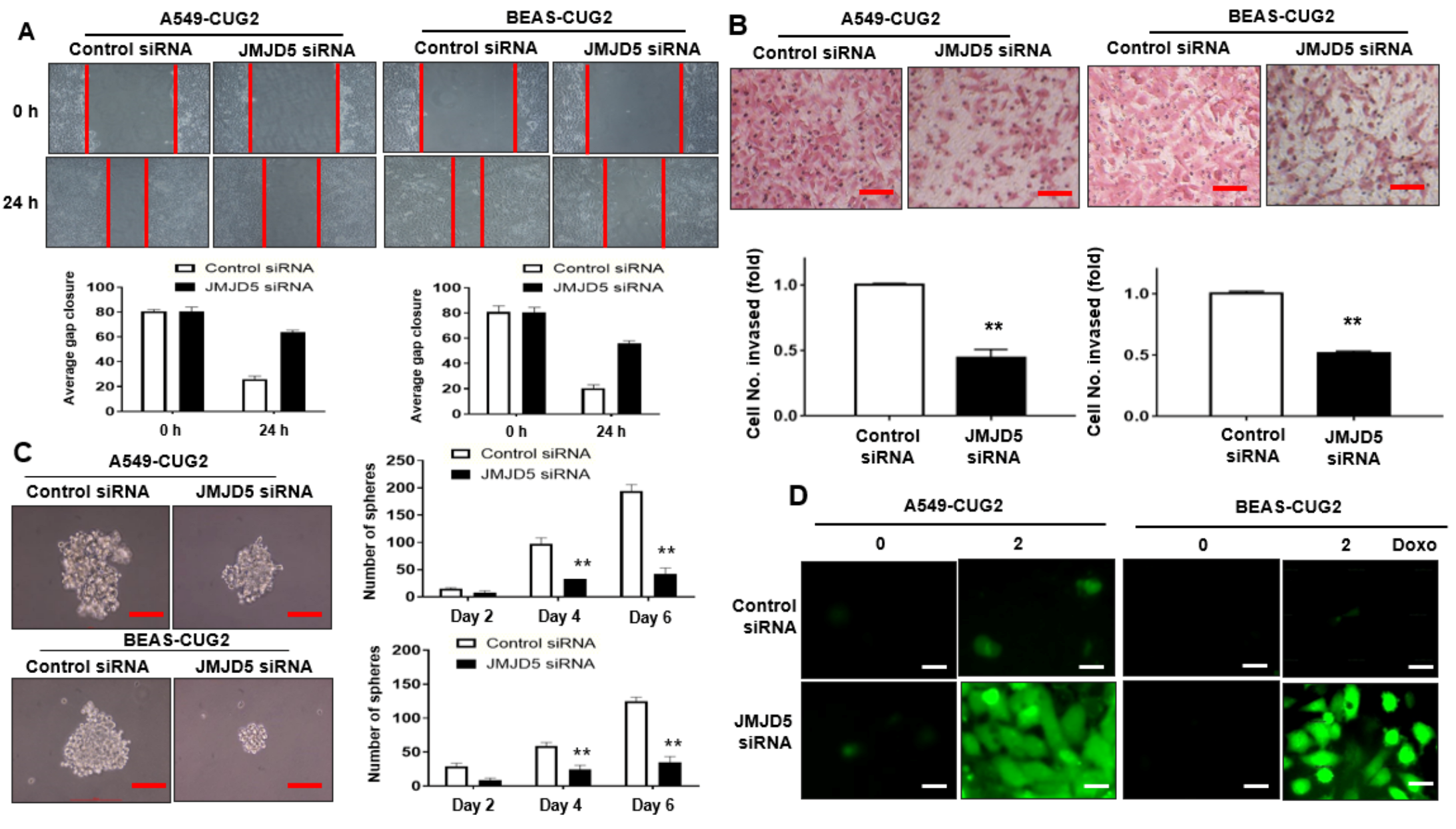
3.3. Suppression of JMJD5, a Target of miR-3656, Inhibits CUG2-Induced CSC-Like Phenotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engels, B.M.; Hutvagner, G. Principles and effects of microRNA-mediated post-transcriptional gene regulation. Oncogene 2006, 25, 6163–6169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacFarlane, L.-A.; R Murphy, P. MicroRNA: Biogenesis, function and role in cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef]
- Nelson, K.M.; Weiss, G.J. MicroRNAs and cancer: Past, present, and potential future. Mol. Cancer Ther. 2008, 7, 3655–3660. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Gang, J.; Jeon, S.B.; Choo, S.H.; Lee, B.; Kim, Y.-G.; Lee, Y.S.; Jung, J.; Song, S.Y.; Koh, S.S. Molecular cloning and functional analysis of a novel oncogene, cancer-upregulated gene 2 (CUG2). Biochem. Biophys. Res. Commun. 2007, 360, 633–639. [Google Scholar] [CrossRef]
- Kaowinn, S.; Kim, J.; Lee, J.; Shin, D.H.; Kang, C.-D.; Kim, D.-K.; Lee, S.; Kang, M.K.; Koh, S.S.; Kim, S.-J. Cancer upregulated gene 2 induces epithelial-mesenchymal transition of human lung cancer cells via TGF-β signaling. Oncotarget 2017, 8, 5092. [Google Scholar] [CrossRef] [Green Version]
- Kaowinn, S.; Seo, E.J.; Heo, W.; Bae, J.-H.; Park, E.-J.; Lee, S.; Kim, Y.J.; Koh, S.S.; Jang, I.H.; Shin, D.H. Cancer upregulated gene 2 (CUG2), a novel oncogene, promotes stemness-like properties via the NPM1-TGF-β signaling axis. Biochem. Biophys. Res. Commun. 2019, 514, 1278–1284. [Google Scholar] [CrossRef]
- Kaowinn, S.; Kaewpiboon, C.; Koh, S.S.; Krämer, O.H.; Chung, Y.H. STAT1-HDAC4 signaling induces epithelial-mesenchymal transition and sphere formation of cancer cells overexpressing the oncogene, CUG2. Oncol. Rep. 2018, 40, 2619–2627. [Google Scholar] [CrossRef]
- Kaowinn, S.; Yawut, N.; Koh, S.S.; Chung, Y.-H. Cancer upregulated gene (CUG) 2 elevates YAP1 expression, leading to enhancement of epithelial-mesenchymal transition in human lung cancer cells. Biochem. Biophys. Res. Commun. 2019, 511, 122–128. [Google Scholar] [CrossRef]
- Yawut, N.; Kaewpiboon, C.; Budluang, P.; Cho, I.-R.; Kaowinn, S.; Koh, S.S.; Chung, Y.-H. Overexpression of Cancer Upregulated Gene 2 (CUG2) Decreases Spry2 Through c-Cbl, Leading to Activation of EGFR and β-Catenin Signaling. Cancer Manag. Res. 2020, 12, 10243. [Google Scholar] [CrossRef]
- Hori, T.; Amano, M.; Suzuki, A.; Backer, C.B.; Welburn, J.P.; Dong, Y.; McEwen, B.F.; Shang, W.-H.; Suzuki, E.; Okawa, K. CCAN makes multiple contacts with centromeric DNA to provide distinct pathways to the outer kinetochore. Cell 2008, 135, 1039–1052. [Google Scholar] [CrossRef] [Green Version]
- Chun, Y.; Kim, R.; Lee, S. Centromere protein (CENP)-W interacts with heterogeneous nuclear ribonucleoprotein (hnRNP) U and may contribute to kinetochore-microtubule attachment in mitotic cells. PLoS ONE 2016, 11, e0149127. [Google Scholar] [CrossRef] [Green Version]
- Prendergast, L.; Müller, S.; Liu, Y.; Huang, H.; Dingli, F.; Loew, D.; Vassias, I.; Patel, D.J.; Sullivan, K.F.; Almouzni, G. The CENP-T/-W complex is a binding partner of the histone chaperone FACT. Genes Dev. 2016, 30, 1313–1326. [Google Scholar] [CrossRef] [Green Version]
- Koh, W.; Park, B.; Lee, S. A new kinetochore component CENP-W interacts with the polycomb-group protein EZH2 to promote gene silencing. Biochem. Biophys. Res. Commun. 2015, 464, 256–262. [Google Scholar] [CrossRef]
- Klose, R.J.; Kallin, E.M.; Zhang, Y. JmjC-domain-containing proteins and histone demethylation. Nat. Rev. Genet. 2006, 7, 715–727. [Google Scholar] [CrossRef]
- Hsia, D.A.; Tepper, C.G.; Pochampalli, M.R.; Hsia, E.Y.; Izumiya, C.; Huerta, S.B.; Wright, M.E.; Chen, H.-W.; Kung, H.-J.; Izumiya, Y. KDM8, a H3K36me2 histone demethylase that acts in the cyclin A1 coding region to regulate cancer cell proliferation. Proc. Natl. Acad. Sci. USA 2010, 107, 9671–9676. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Wu, J.; Su, X.; Zhang, Y.; Pan, L.; Wei, H.; Fang, Q.; Li, H.; Wang, D.-L.; Sun, F.-L. JMJD5 (Jumonji Domain-containing 5) associates with spindle microtubules and is required for proper mitosis. J. Biol. Chem. 2016, 291, 4684–4697. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Zhang, S.; Qi, H.; Wang, Z.; Chen, H.-W.; Shao, J.; Shen, J. JMJD5 interacts with p53 and negatively regulates p53 function in control of cell cycle and proliferation. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2015, 1853, 2286–2295. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Hu, H.; Chang, R.; Zhong, J.; Knabel, M.; O’Meally, R.; Cole, R.N.; Pandey, A.; Semenza, G.L. Pyruvate kinase M2 is a PHD3-stimulated coactivator for hypoxia-inducible factor 1. Cell 2011, 145, 732–744. [Google Scholar] [CrossRef] [Green Version]
- Kaowinn, S.; Jun, S.W.; Kim, C.S.; Shin, D.-M.; Hwang, Y.-H.; Kim, K.; Shin, B.; Kaewpiboon, C.; Jeong, H.H.; Koh, S.S. Increased EGFR expression induced by a novel oncogene, CUG2, confers resistance to doxorubicin through Stat1-HDAC4 signaling. Cell. Oncol. 2017, 40, 549–561. [Google Scholar] [CrossRef]
- Varier, R.A.; Timmers, H.M. Histone lysine methylation and demethylation pathways in cancer. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2011, 1815, 75–89. [Google Scholar] [CrossRef]
- Zhao, Z.; Sun, C.; Li, F.; Han, J.; Li, X.; Song, Z. Overexpression of histone demethylase JMJD5 promotes metastasis and indicates a poor prognosis in breast cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 10325. [Google Scholar]
- Yao, Y.; Zhou, W.-Y.; He, R.-X. Down-regulation of JMJD5 suppresses metastasis and induces apoptosis in oral squamous cell carcinoma by regulating p53/NF-κB pathway. Biomed. Pharmacother. 2019, 109, 1994–2004. [Google Scholar] [CrossRef]
- Zhang, R.; Huang, Q.; Li, Y.; Song, Y.; Li, Y. JMJD5 is a potential oncogene for colon carcinogenesis. Int. J. Clin. Exp. Pathol. 2015, 8, 6482. [Google Scholar]
- Matamala, N.; Vargas, M.T.; Gonzalez-Campora, R.; Minambres, R.; Arias, J.I.; Menendez, P.; Andres-Leon, E.; Gomez-Lopez, G.; Yanowsky, K.; Calvete-Candenas, J. Tumor microRNA expression profiling identifies circulating microRNAs for early breast cancer detection. Clin. Chem. 2015, 61, 1098–1106. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Zhu, Z.; Wang, H.; Yu, Y.; Chen, W.; Waqas, A.; Wang, Y.; Chen, L. Exosomes derived from human neural stem cells stimulated by interferon gamma improve therapeutic ability in ischemic stroke model. J. Adv. Res. 2020, 24, 435–445. [Google Scholar] [CrossRef]
- Yang, R.-M.; Zhan, M.; Xu, S.-W.; Long, M.-M.; Yang, L.-H.; Chen, W.; Huang, S.; Liu, Q.; Zhou, J.; Zhu, J. miR-3656 expression enhances the chemosensitivity of pancreatic cancer to gemcitabine through modulation of the RHOF/EMT axis. Cell Death Dis. 2017, 8, e3129. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Gao, F.; Lv, L.; Chen, X. The interaction of hypertension and homocysteine increases the risk of mortality among middle-aged and older population in the United States. J. Hypertens. 2021, 40, 254–263. [Google Scholar] [CrossRef]
- Ali Syeda, Z.; Langden, S.S.S.; Munkhzul, C.; Lee, M.; Song, S.J. Regulatory mechanism of microRNA expression in cancer. Int. J. Mol. Sci. 2020, 21, 1723. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.; Y Cheng, Y.; Blenkiron, C.; Reid, G. Exploring mechanisms of microRNA downregulation in cancer. MicroRNA 2017, 6, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Ding, L.; Bennewith, K.L.; Tong, R.T.; Welford, S.M.; Ang, K.K.; Story, M.; Le, Q.-T.; Giaccia, A.J. Hypoxia-inducible mir-210 regulates normoxic gene expression involved in tumor initiation. Mol. Cell 2009, 35, 856–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, T.-C.; Yu, D.; Lee, Y.-S.; Wentzel, E.A.; Arking, D.E.; West, K.M.; Dang, C.V.; Thomas-Tikhonenko, A.; Mendell, J.T. Widespread microRNA repression by Myc contributes to tumorigenesis. Nat. Genet. 2008, 40, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kent, O.; Fox-Talbot, K.; Halushka, M. RREB1 repressed miR-143/145 modulates KRAS signaling through downregulation of multiple targets. Oncogene 2013, 32, 2576–2585. [Google Scholar] [CrossRef] [Green Version]
- He, L.; He, X.; Lim, L.P.; De Stanchina, E.; Xuan, Z.; Liang, Y.; Xue, W.; Zender, L.; Magnus, J.; Ridzon, D. A microRNA component of the p53 tumour suppressor network. Nature 2007, 447, 1130–1134. [Google Scholar] [CrossRef] [Green Version]
- Shindo, T.; Niinuma, T.; Nishiyama, N.; Shinkai, N.; Kitajima, H.; Kai, M.; Maruyama, R.; Tokino, T.; Masumori, N.; Suzuki, H. Epigenetic silencing of miR-200b is associated with cisplatin resistance in bladder cancer. Oncotarget 2018, 9, 24457. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.-J.; Wang, Q.-Y.; Zhou, C.-X.; Yin, Q.-Q.; He, M.; Yu, X.-T.; Cao, D.-X.; Chen, G.-Q.; He, J.-R.; Zhao, Q. MiR-124 targets Slug to regulate epithelial–mesenchymal transition and metastasis of breast cancer. Carcinogenesis 2013, 34, 713–722. [Google Scholar] [CrossRef]
- Saito, Y.; Liang, G.; Egger, G.; Friedman, J.M.; Chuang, J.C.; Coetzee, G.A.; Jones, P.A. Specific activation of microRNA-127 with downregulation of the proto-oncogene BCL6 by chromatin-modifying drugs in human cancer cells. Cancer Cell 2006, 9, 435–443. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yawut, N.; Cho, I.-R.; Budluang, P.; Kaowinn, S.; Kaewpiboon, C.; Jeon, B.; Kim, S.-W.; Kang, H.Y.; Kang, M.-K.; Koh, S.S.; et al. Elevated Expression of JMJD5 Protein Due to Decreased miR-3656 Levels Contributes to Cancer Stem Cell-Like Phenotypes under Overexpression of Cancer Upregulated Gene 2. Biomolecules 2022, 12, 122. https://doi.org/10.3390/biom12010122
Yawut N, Cho I-R, Budluang P, Kaowinn S, Kaewpiboon C, Jeon B, Kim S-W, Kang HY, Kang M-K, Koh SS, et al. Elevated Expression of JMJD5 Protein Due to Decreased miR-3656 Levels Contributes to Cancer Stem Cell-Like Phenotypes under Overexpression of Cancer Upregulated Gene 2. Biomolecules. 2022; 12(1):122. https://doi.org/10.3390/biom12010122
Chicago/Turabian StyleYawut, Natpaphan, Il-Rae Cho, Phatcharaporn Budluang, Sirichat Kaowinn, Chutima Kaewpiboon, Byeoleun Jeon, Sang-Woo Kim, Ho Young Kang, Min-Kyung Kang, Sang Seok Koh, and et al. 2022. "Elevated Expression of JMJD5 Protein Due to Decreased miR-3656 Levels Contributes to Cancer Stem Cell-Like Phenotypes under Overexpression of Cancer Upregulated Gene 2" Biomolecules 12, no. 1: 122. https://doi.org/10.3390/biom12010122
APA StyleYawut, N., Cho, I. -R., Budluang, P., Kaowinn, S., Kaewpiboon, C., Jeon, B., Kim, S. -W., Kang, H. Y., Kang, M. -K., Koh, S. S., & Chung, Y. -H. (2022). Elevated Expression of JMJD5 Protein Due to Decreased miR-3656 Levels Contributes to Cancer Stem Cell-Like Phenotypes under Overexpression of Cancer Upregulated Gene 2. Biomolecules, 12(1), 122. https://doi.org/10.3390/biom12010122