

Amygdalin: A Review on Its Characteristics, Antioxidant Potential, Gastrointestinal Microbiota Intervention, Anticancer Therapeutic and Mechanisms, Toxicity, and Encapsulation

 ,
,  , , , ,
, , , , 
Abstract
:1. Introduction
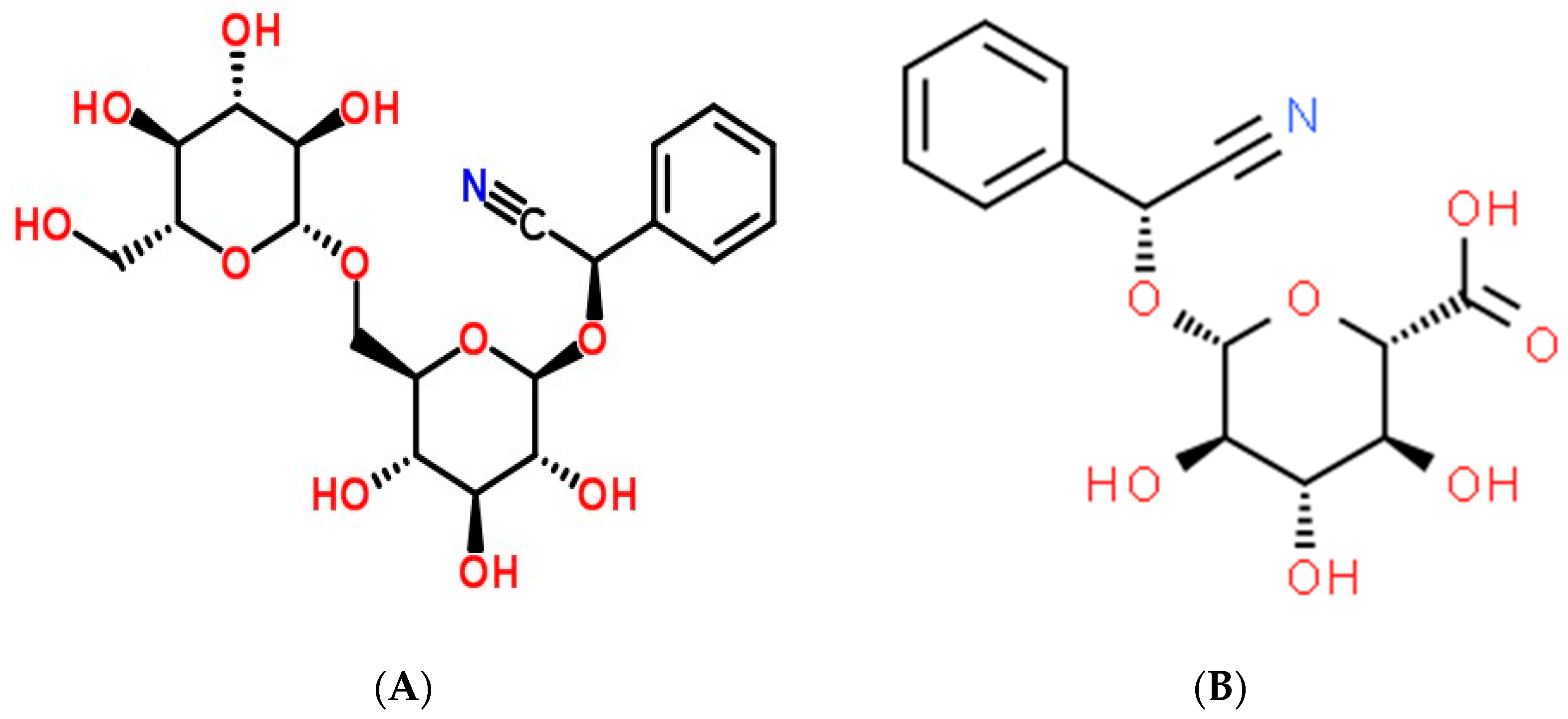
2. History and Structural Characteristics of Amygdalin as an Anticancer Therapy
3. In Vivo and In Vitro Antioxidant Potential of Amygdalin
4. The Microbiome and Oral Amygdalin Administration: Intervention with Gut Microbiota
5. Anticancer Therapeutic Effectiveness of Amygdalin
5.1. Anticancer Effects
5.1.1. Amygdalin as an Anti-Tumor Drug
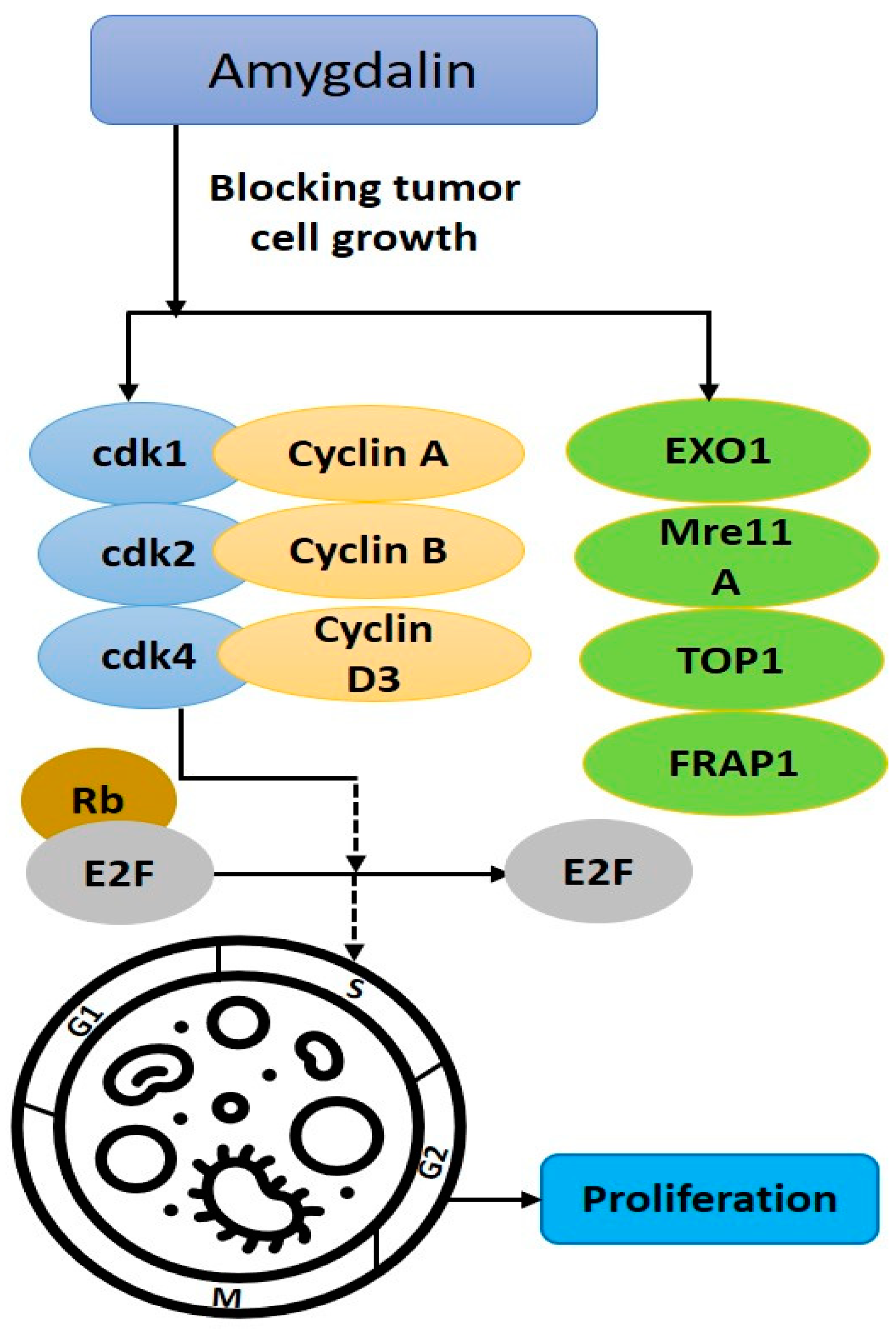
5.1.2. Inhibiting Tumor Cell Growth
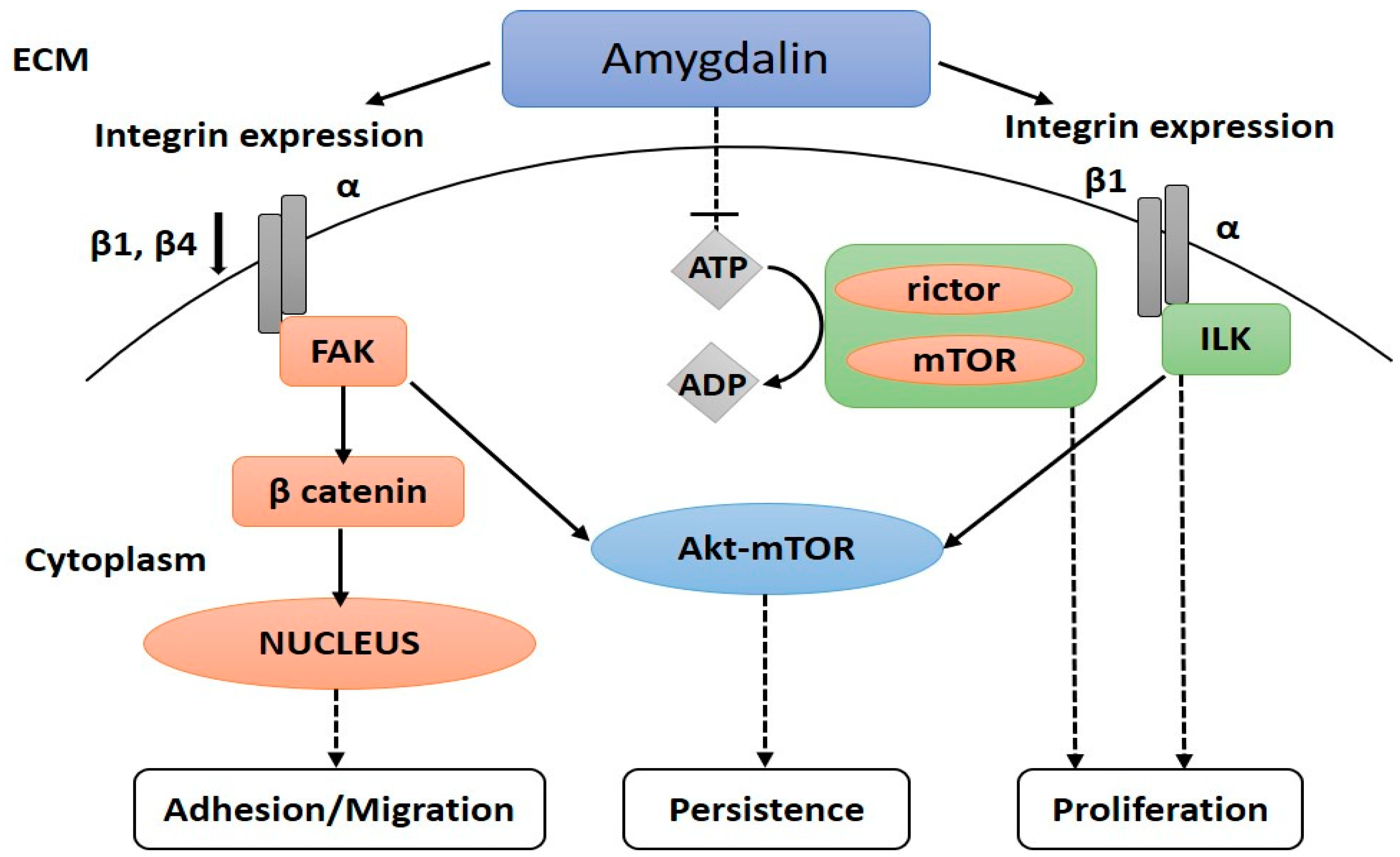
5.1.3. Reducing the Spread of Cancer Cells
5.1.4. Inducing Apoptosis
5.2. Anti-Fibrosis Effect
5.2.1. Modulating the Immune System
5.2.2. Efficacy in Preventing Atherosclerosis
5.2.3. Contribution to the Reproductive Process
5.2.4. Neurodegeneration Improvement
5.2.5. Anti-Renal Interstitial Fibrosis
5.2.6. Anti-Liver Fibrosis
5.2.7. Anti-Pulmonary Fibrosis
5.2.8. Anti-Inflammatory Effect
5.2.9. Pain-Relieving Effect
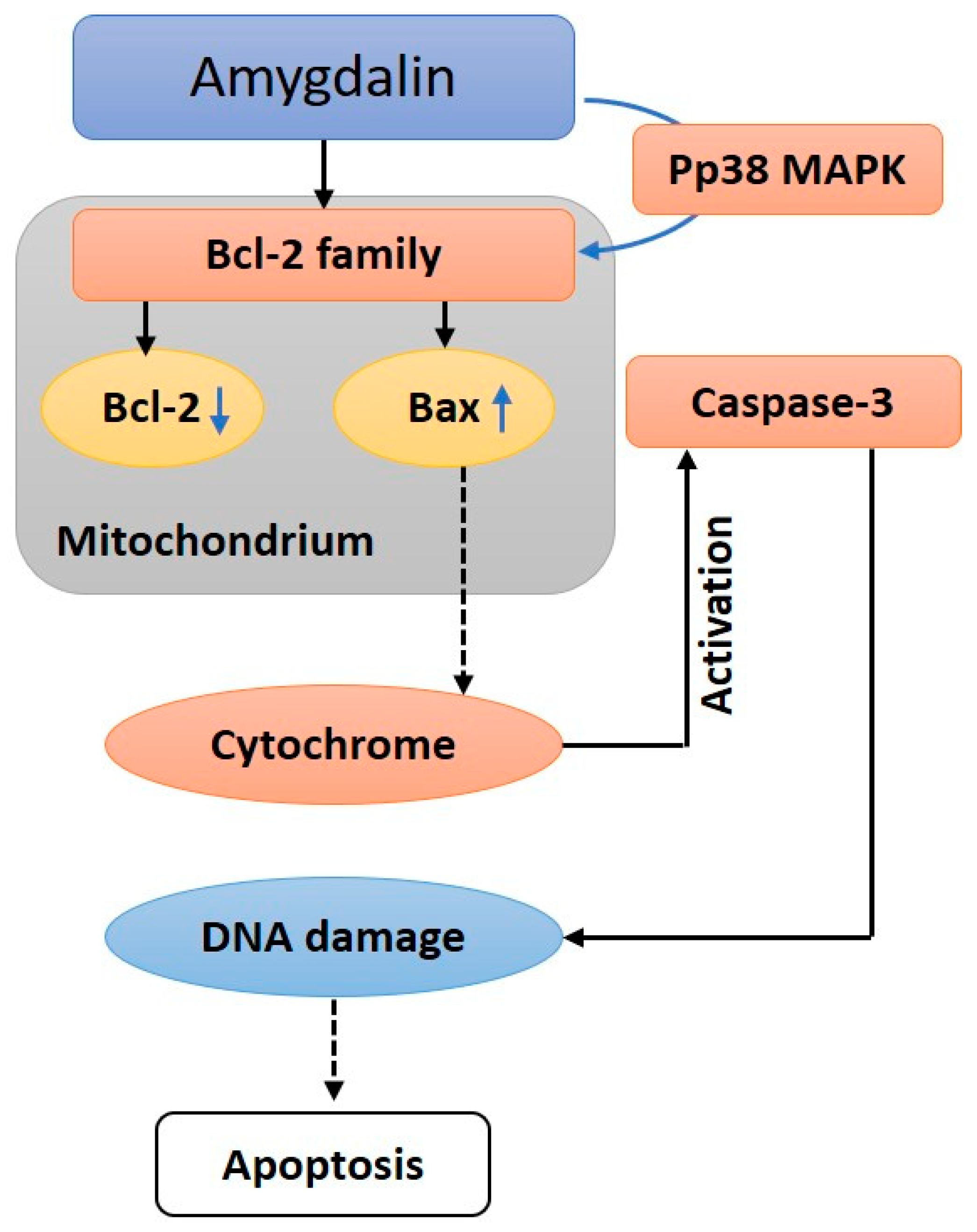
6. Anticancer Mechanisms of Amygdalin: A Molecular Approach
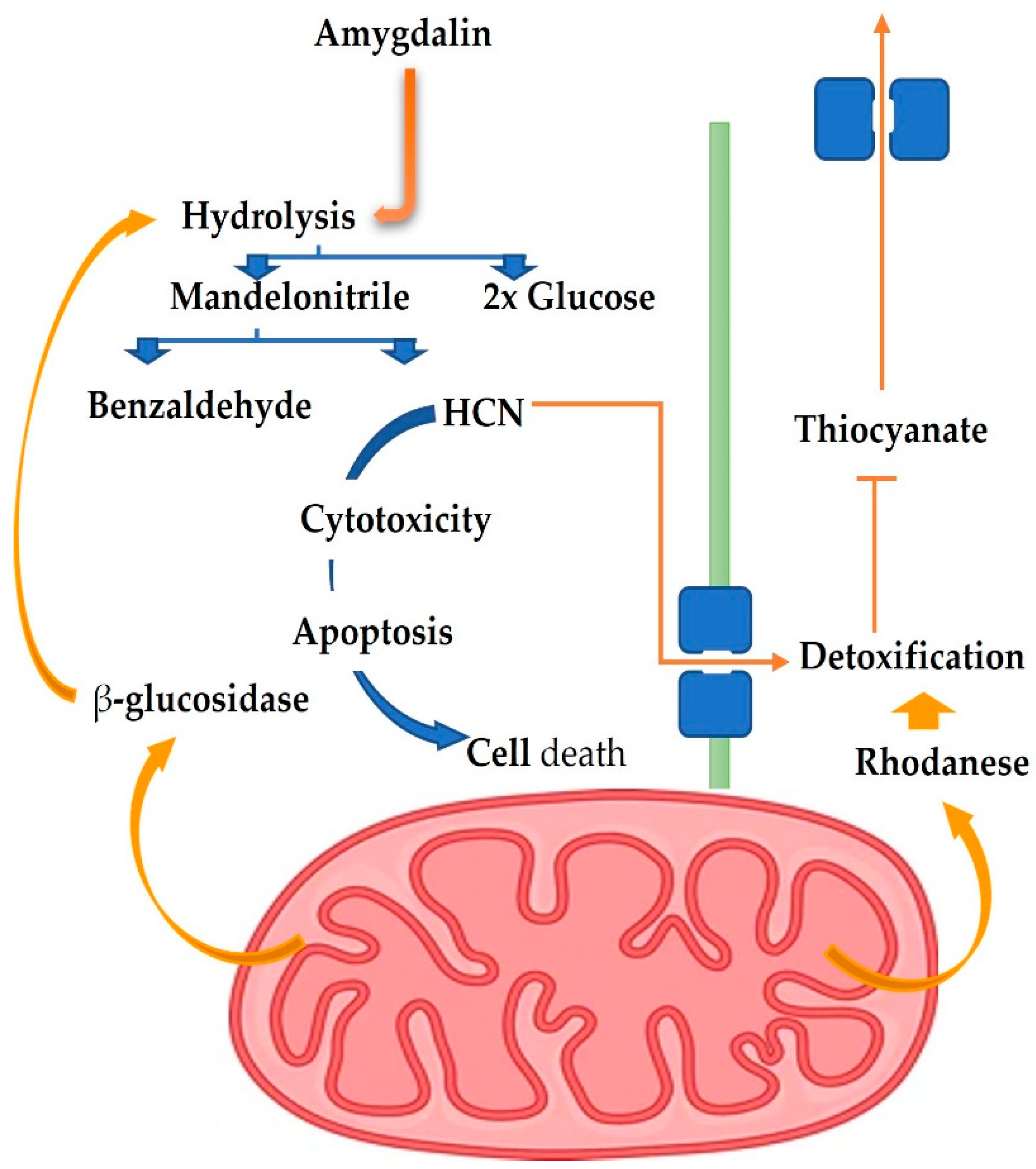
7. Toxicological Effects of Amygdalin
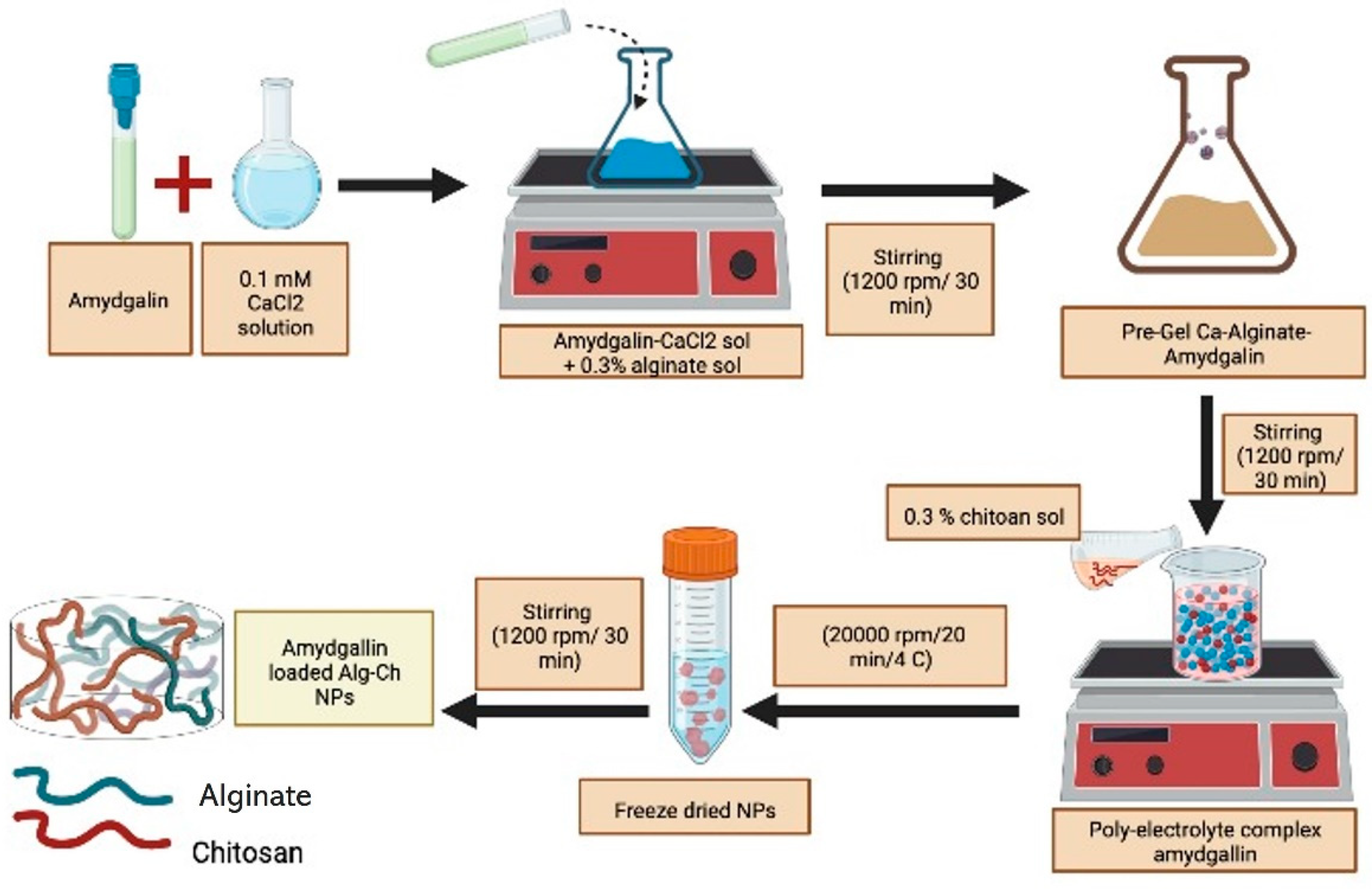
8. Microencapsulation and Bioavailability
9. Future Perspectives and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Jaswal, V.; Palanivelu, J.; Ramalingam, C. Effects of The Gut Microbiota on Amygdalin and Its Use as an Anti-Cancer Therapy: Substantial Review on the Key Components Involved in Altering Dose Efficacy and Toxicity. Biochem. Biophys. Rep. 2018, 14, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Toomey, V.M.; Nickum, E.A.; Flurer, C.L. Cyanide and Amygdalin as Indicators of the Presence of Bitter Almonds in Imported Raw Almonds. J. Forensic Sci. 2012, 57, 1313–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlikova, B.; Legrand, N.; Panning, J.; Dicato, M.; Diederich, M. Anti-Inflammatory and Anticancer Drugs from Nature. In Advances in Nutrition and Cancer; Springer: Berlin/Heidelberg, Germany, 2014; pp. 123–143. [Google Scholar]
- Qadir, M.; Fatima, K. Review on Pharmacological Activity of Amygdalin. Arch. Cancer Res. 2017, 5, 160. [Google Scholar] [CrossRef] [Green Version]
- Barakat, H. Amygdalin as A Plant-Based Bioactive Constituent: An Updated Substantial Review on Intervention with Gut Microbiota, Anticancer Therapy, Bioavailability, and Microencapsulation. In Proceedings of the 1st International Electronic Conference on Nutrients—Nutritional and Microbiota Effects on Chronic Disease session Potential nutraceutical effects of nutrients, phytochemicals, and microbiota in chronic metabolic disorders, online, 2–15 November 2020. [Google Scholar]
- Li, X.-B.; Liu, C.-H.; Zhang, R.; Huang, X.-T.; Li, Y.-Y.; Han, L.; Xu, M.-L.; Mi, S.-Q.; Wang, N.-S. Determination and Pharmacokinetics of Amygdalin in Rats By LC–MS-MS. J. Chromatogr. Sci. 2014, 52, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Xu, X. Advanced Research on Anti-tumor Effects of Amygdalin. J. Cancer Res. 2014, 10, 3. [Google Scholar]
- Chang, H.-K.; Shin, M.-S.; Yang, H.-Y.; Lee, J.-W.; Kim, Y.-S.; Lee, M.-H.; Kim, J.; Kim, K.-H.; Kim, C.-J. Amygdalin Induces Apoptosis Through Regulation of Bax and Bcl-2 Expressions in Human Du145 and Lncap Prostate Cancer Cells. Biol. Pharm. Bull. 2006, 29, 1597–1602. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Qian, L.; Ma, H.; Yu, X.; Zhang, Y.; Qu, W.; Zhang, X.; Xia, W. Enhancement of Amygdalin Activated with Β-D-Glucosidase on HEPG2 Cells Proliferation and Apoptosis. Carbohydr. Polym. 2012, 90, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-J.; Yoon, S.-H.; Han, L.-S.; Zheng, L.-T.; Jung, K.-H.; Uhm, Y.-K.; Lee, J.-H.; Jeong, J.-S.; Joo, W.-S.; Yim, S.-V. Amygdalin Inhibits Genes Related to Cell Cycle in Snu-C4 Human Colon Cancer Cells. World J. Gastroenterol. 2005, 11, 5156. [Google Scholar]
- Chen, Y.; Ma, J.; Wang, F.; Hu, J.; Cui, A.; Wei, C.; Yang, Q.; Li, F. Amygdalin Induces Apoptosis in Human Cervical Cancer Cell Line Hela Cells. Immunopharmacol. Immunotoxicol. 2013, 35, 43–51. [Google Scholar] [CrossRef]
- Dorr, R.T.; Paxinos, J. The Current Status of Laetrile. Ann. Intern. Med. 1978, 89, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Davignon, J.P.; Trissel, L.A.; Kleinman, L.M. Pharmaceutical Assessment of Amygdalin (Laetrile) Products. Cancer Treat. Rep. 1978, 62, 99–104. [Google Scholar] [PubMed]
- Moertel, C.G.; Fleming, T.R.; Rubin, J.; Kvols, L.K.; Sarna, G.; Koch, R.; Currie, V.E.; Young, C.W.; Jones, S.E.; Davignon, J.P. A Clinical Trial of Amygdalin (Laetrile) in The Treatment of Human Cancer. N. Engl. J. Med. 1982, 306, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Society, A.C. Actualización de American Cancer Society 1991.Questionable Cancer Practices in Tijuana and Other Mexican Border Clinics. Cancer J. Clin. 1991, 41, 310–319. [Google Scholar]
- Milazzo, S.; Lejeune, S.; Ernst, E. Laetrile for Cancer: A Systematic Review of the Clinical Evidence. Support. Care Cancer 2007, 15, 583–595. [Google Scholar] [CrossRef]
- Bolarinwa, I.F.; Orfila, C.; Morgan, M.R. Amygdalin Content of Seeds, Kernels and Food Products Commercially-Available in The Uk. Food Chem. 2014, 152, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Sireesha, D.; Reddy, B.S.; Reginald, B.A.; Samatha, M.; Kamal, F. Effect of Amygdalin on Oral Cancer Cell Line: An in Vitro Study. J. Oral Maxillofac. Pathol. 2019, 23, 104. [Google Scholar]
- Shi, J.; Chen, Q.; Xu, M.; Xia, Q.; Zheng, T.; Teng, J.; Li, M.; Fan, L. Recent Updates and Future Perspectives about Amygdalin as A Potential Anticancer Agent: A Review. Cancer Med. 2019, 8, 3004–3011. [Google Scholar] [CrossRef]
- Mirhosseini, H.; Amid, B.T. Effect of Different Drying Techniques on Flowability Characteristics and Chemical Properties of Natural Carbohydrate-Protein Gum from Durian Fruit Seed. Chem. Cent. J. 2013, 7, 44. [Google Scholar] [CrossRef] [Green Version]
- Binita, K.; Kumar, S.; Sharma, V.K.; Sharma, V.; Yadav, S. Proteomic Identification of Syzygium cumini Seed Extracts by MALDI-TOF/MS. Appl. Biochem. Biotechnol. 2014, 172, 2091–2105. [Google Scholar] [CrossRef]
- Ambigaipalan, P.; Al-Khalifa, A.S.; Shahidi, F. Antioxidant and Angiotensin I Converting Enzyme (Ace) Inhibitory Activities of Date Seed Protein Hydrolysates Prepared Using Alcalase, Flavourzyme and Thermolysin. J. Funct. Foods 2015, 18, 1125–1137. [Google Scholar] [CrossRef]
- Stosic, D.; Gorunovic, M.; Popovic, B. Preliminary Toxicological Study of The Kernel and The Oil of Some Prunus Species. Plantes Med. Et. Phytother. 1987, 21, 8–13. [Google Scholar]
- Tunçel, G.; Nout, M.J.R.; Brimer, L. Degradation of Cyanogenic Glycosides of Bitter Apricot Seeds (Prunus Armeniaca) by Endogenous and Added Enzymes as Affected by Heat Treatments and Particle Size. Food Chem. 1998, 63, 65–69. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Schrenk, D.; Bignami, M.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.-C.; et al. Evaluation of the Health Risks Related to the Presence of Cyanogenic Glycosides in Foods other than Raw Apricot Kernels. EFSA J. 2019, 17, e05662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, M.C.; Gonzalez-Garcia, E.; Vasquez-Villanueva, R.; Marina, M.L. Apricot and other Seed Stones: Amygdalin Content and the Potential to Obtain Antioxidant, Angiotensin I Converting Enzyme Inhibitor and Hypocholesterolemic Peptides. Food Funct. 2016, 7, 4693–4701. [Google Scholar] [CrossRef] [PubMed]
- Alwan, A.M.; Afshari, J.T. In Vivo Growth Inhibition of Human Caucasian Prostate Adenocarcinoma in Nude Mice Induced by Amygdalin with Metabolic Enzyme Combinations. Biomed. Res. Int. 2022, 2022, 4767621. [Google Scholar] [CrossRef]
- Albogami, S.; Hassan, A.; Ahmed, N.; Alnefaie, A.; Alattas, A.; Alquthami, L.; Alharbi, A. Evaluation of the Effective Dose of Amygdalin for the Improvement of Antioxidant Gene Expression and Suppression of Oxidative Damage in Mice. PeerJ 2020, 8, e9232. [Google Scholar] [CrossRef]
- Moslehi, A.; Komeili-movahed, T.; Moslehi, M. Antioxidant Effects of Amygdalin on Tunicamycin-Induced Endoplasmic Reticulum Stress in The Mice Liver: Cross Talk Between Endoplasmic Reticulum Stress and Oxidative Stress. J. Rep. Pharm. Sci. 2019, 8, 298. [Google Scholar] [CrossRef]
- Sushma, P.; Jacob, B.; Narendhirakannan, R.T. Evaluation of Antioxidant And Cytotoxicity Properties Of Amygdalin Extracted From Prunus Dulcis. Kongunadu Res. J. 2019, 6, 8–12. [Google Scholar] [CrossRef]
- Sokkar, N.; El-Gindi, O.; Sayed, S.; Mohamed, S.; Ali, Z.; Alfishawy, I. Antioxidant, Anticancer and Hepatoprotective Activities of Cotoneaster Horizontalis Decne Extract as well as A-Tocopherol and Amygdalin Production from in Vitro Culture. Acta Physiol. Plant. 2013, 35, 2421–2428. [Google Scholar] [CrossRef]
- Ushakova, N.; Nekrasov, R.; Pravdin, I.; Sverchkova, N.; Kolomiyets, E.; Pavlov, D. Mechanisms of the Effects of Probiotics on Symbiotic Digestion. Biol. Bull. 2015, 42, 394–400. [Google Scholar] [CrossRef]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of Dietary Compounds, Especially Polyphenols, with The Intestinal Microbiota: A Review. Eur. J. Nutr. 2015, 54, 325–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Sousa, I.P.; Bernkop-Schnürch, A. Pre-Systemic Metabolism of Orally Administered Drugs and Strategies to Overcome it. J. Control Release. 2014, 192, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Berrin, J.-G.; Czjzek, M.; Kroon, P.A.; McLauchlan, W.R.; Puigserver, A.; Williamson, G.; Juge, N.J.B.J. Substrate (Aglycone) Specificity of Human Cytosolic Beta-Glucosidase. Biochem. J. 2003, 373, 41–48. [Google Scholar] [CrossRef]
- Serban, D.E. Gastrointestinal Cancers: Influence of Gut Microbiota, Probiotics and Prebiotics. Cancer Lett. 2014, 345, 258–270. [Google Scholar] [CrossRef]
- Shapiro, S.E.; Donaldson, N.A. Evidence-Based Practice for Advanced Practice Emergency Nurses, Part Ii: Critically Appraising the Literature. Adv. Emerg. Nurs. J. 2008, 30, 139–150. [Google Scholar] [CrossRef]
- Chang, J.; Zhang, Y.J.P.B. Catalytic Degradation of Amygdalin by Extracellular Enzymes from Aspergillus niger. Process Biochem. 2012, 47, 195–200. [Google Scholar] [CrossRef]
- Blaheta, R.A.; Nelson, K.; Haferkamp, A.; Juengel, E. Amygdalin, quackery or cure? Phytomedicine 2016, 23, 367–376. [Google Scholar] [CrossRef]
- Tokpohozin, S.E.; Fischer, S.; Sacher, B.; Becker, T. Β-D-Glucosidase As “Key Enzyme” for Sorghum Cyanogenic Glucoside (Dhurrin) Removal and Beer Bioflavouring. Food Chem. Toxicol. 2016, 97, 217–223. [Google Scholar] [CrossRef]
- Chong, E.S.L. A Potential Role of Probiotics in Colorectal Cancer Prevention: Review of Possible Mechanisms of Action. World J. Microbiol. Biotechnol. 2014, 30, 351–374. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, N.; Qian, C.; Wang, Q.; Wang, Q.; Long, Y.; Huang, Y.; Zhou, Z.; Yan, X. Phylogenetic and Functional Analysis of Gut Microbiota of A Fungus-Growing Higher Termite: Bacteroidetes from Higher Termites are A Rich Source of Β-Glucosidase Genes. Microb. Ecol. 2014, 68, 416–425. [Google Scholar] [CrossRef]
- Sorbo, B. On the Properties of Rhodanese: Partial Purification, Inhibitors and Intracellular Distribution. Acta Chem. Scand. 1951, 5, 724–726. [Google Scholar] [CrossRef]
- Greenberg, D.M. The Vitamin Fraud in Cancer Quackery. West. J. Med. 1975, 122, 345. [Google Scholar] [PubMed]
- Baffoni, L.; Gaggìa, F.; Di Gioia, D.; Biavati, B. Role of Intestinal Microbiota in Colon Cancer Prevention. Ann. Microbiol. 2012, 62, 15–30. [Google Scholar] [CrossRef]
- Hullar, M.A.; Burnett-Hartman, A.N.; Lampe, J.W. Gut Microbes, Diet, and Cancer. In Advances in Nutrition and Cancer; Springer: Berlin/Heidelberg, Germany, 2014; pp. 377–399. [Google Scholar]
- Kleessen, B.; Sykura, B.; Zunft, H.-J.; Blaut, M. Effects of Inulin and Lactose on Fecal Microflora, Microbial Activity, and Bowel Habit in Elderly Constipated Persons. Am. J. Clin. Nutr. 1997, 65, 1397–1402. [Google Scholar] [CrossRef] [Green Version]
- Chiang, H.C. A Hybrid Two-Component System in Bacteroides Thetaiotaomicron That Regulates Nitrogen Metabolism. Ph.D. Thesis, Washington University in St. Louis, Seattle, WA, USA, 2009. [Google Scholar]
- Ravcheev, D.A.; Godzik, A.; Osterman, A.L.; Rodionov, D.A. Polysaccharides Utilization in Human Gut Bacterium Bacteroides Thetaiotaomicron: Comparative Genomics Reconstruction of Metabolic and Regulatory Networks. BMC Genom. 2013, 14, 873. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, F.H.; Ussery, D.W.; Nielsen, J.; Nookaew, I. A Closer Look at Bacteroides: Phylogenetic Relationship and Genomic Implications of A Life in the Human Gut. Microb. Ecol. 2011, 61, 473–485. [Google Scholar] [CrossRef] [Green Version]
- Steer, T.E.; Johnson, I.T.; Gee, J.M.; Gibson, G.R. Metabolism of The Soyabean Isoflavone Glycoside Genistin in Vitro by Human Gut Bacteria and the Effect of Prebiotics. Br. J. Nutr. 2003, 90, 635–642. [Google Scholar] [CrossRef] [Green Version]
- Kuugbee, E.D.; Shang, X.; Gamallat, Y.; Bamba, D.; Awadasseid, A.; Suliman, M.A.; Zang, S.; Ma, Y.; Chiwala, G.; Xin, Y. Structural Change in Microbiota by A Probiotic Cocktail Enhances the Gut Barrier and Reduces Cancer Via TLR2 Signaling in A Rat Model of Colon Cancer. Dig. Dis. Sci. 2016, 61, 2908–2920. [Google Scholar] [CrossRef]
- Bromley, J.; Hughes, B.G.; Leong, D.C.; Buckley, N.A. Life-Threatening Interaction Between Complementary Medicines: Cyanide Toxicity Following Ingestion of Amygdalin and Vitamin C. Ann. Pharm. 2005, 39, 1566–1569. [Google Scholar] [CrossRef]
- Padayatty, S.J.; Sun, A.Y.; Chen, Q.; Espey, M.G.; Drisko, J.; Levine, M. Vitamin C: Intravenous Use by Complementary and Alternative Medicine Practitioners and Adverse Effects. PLoS ONE 2010, 5, e11414. [Google Scholar] [CrossRef] [Green Version]
- Richards, E. The Politics of Therapeutic Evaluation: The Vitamin C and Cancer Controversy. Soc. Stud. Sci. 1988, 18, 653–701. [Google Scholar] [CrossRef]
- Calabrese, E.J. Conjoint Use of Laetrile and Megadoses of Ascorbic Acid in Cancer Treatment: Possible Side Effects. Med. Hypotheses 1979, 5, 995–997. [Google Scholar] [CrossRef]
- Ward, E.; Jemal, A.; Cokkinides, V.; Singh, G.K.; Cardinez, C.; Ghafoor, A.; Thun, M. Cancer Disparities by Race/Ethnicity and Socioeconomic Status. CA Cancer J. Clin. 2004, 54, 78–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holstege, C.P.; Forrester, J.D.; Borek, H.A.; Lawrence, D.T. A Case of Cyanide Poisoning and the Use of Arterial Blood Gas Analysis to Direct Therapy. Hosp. Pract. 2010, 38, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Borron, S.W.; Baud, F.J.; Mégarbane, B.; Bismuth, C. Hydroxocobalamin for Severe Acute Cyanide Poisoning by Ingestion or Inhalation. Am. J. Emerg. Med. 2007, 25, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Oyewole, O.; Olayinka, E. Hydroxocobalamin (vit b12a) Effectively Reduced Extent of Cyanide Poisoning Arising from Oral Amygdalin Ingestion in Rats. J. Toxicol. Environ. Health 2009, 1, 008–011. [Google Scholar]
- Chan, T.Y. A Probable Case of Amygdalin-Induced Peripheral Neuropathy in A Vegetarian with Vitamin B12 Deficiency. Ther. Drug Monit. 2006, 28, 140–141. [Google Scholar] [CrossRef]
- Attia, A.A.; Salama, A.F.; Eldiasty, J.G.; Mosallam, S.A.E.; El-Naggar, S.A.; El-Magd, M.A.; Nasser, H.M.; Elmetwalli, A. Amygdalin Potentiates the Anti-Cancer Effect of Sorafenib on Ehrlich ascites Carcinoma and Ameliorates the Associated Liver Damage. Sci. Rep. 2022, 12, 6494. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Wen, J.; Xu, X.; Shi, J.; Zhang, W.; Zheng, T.; Hou, Y.; Zhang, Y.; Li, Z.; Wang, K.; et al. Amygdalin Induced Mitochondria-Mediated Apoptosis of Lung Cancer Cells via Regulating NF [Formula: See text]B-1/NF[Formula: See text]B Signaling Cascade in Vitro and in Vivo. Am. J. Chin. Med. 2022, 50, 1361–1386. [Google Scholar] [CrossRef]
- Tsaur, I.; Thomas, A.; Monecke, M.; Zugelder, M.; Rutz, J.; Grein, T.; Maxeiner, S.; Xie, H.; Chun, F.K.; Rothweiler, F.; et al. Amygdalin Exerts Antitumor Activity in Taxane-Resistant Prostate Cancer Cells. Cancers 2022, 14, 3111. [Google Scholar] [CrossRef] [PubMed]
- Albogami, S.; Alnefaie, A. Role of Amygdalin in Blocking DNA Replication in Breast Cancer In Vitro. Curr. Pharm. Biotechnol. 2021, 22, 1612–1627. [Google Scholar] [CrossRef]
- Al-Khafaji, K.; Taskin Tok, T. Amygdalin as Multi-Target Anticancer Drug Against Targets of Cell Division Cycle: Double Docking and Molecular Dynamics Simulation. J. Biomol. Struct. Dyn. 2021, 39, 1965–1974. [Google Scholar] [CrossRef]
- Aydin, D.; Ozkan, K.; Aydin, A. The Combination of Amygdalin with Some Anticancer, Antiparasitic, and Antigout Drugs Against MG63, Saos2, SW1353, and FL Cells In Vitro. J. Med. Food 2021, 24, 1230–1234. [Google Scholar] [CrossRef] [PubMed]
- Jaszczak-Wilke, E.; Polkowska, Z.; Koprowski, M.; Owsianik, K.; Mitchell, A.E.; Balczewski, P. Amygdalin: Toxicity, Anticancer Activity and Analytical Procedures for Its Determination in Plant Seeds. Molecules 2021, 26, 2253. [Google Scholar] [CrossRef]
- Mosayyebi, B.; Mohammadi, L.; Kalantary-Charvadeh, A.; Rahmati, M. Amygdalin Decreases Adhesion and Migration of MDA-MB-231 and MCF-7 Breast Cancer Cell Lines. Curr. Mol. Pharmacol. 2021, 14, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Hosny, S.; Sahyon, H.; Youssef, M.; Negm, A. Prunus Armeniaca L. Seed Extract and Its Amygdalin Containing Fraction Induced Mitochondrial-Mediated Apoptosis and Autophagy in Liver Carcinogenesis. Anticancer Agents Med. Chem. 2021, 21, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Si, Z.; Zhang, B. Amygdalin Attenuates Airway Epithelium Apoptosis, Inflammation, and Epithelial-Mesenchymal Transition through Restraining the TLR4/NF-kappaB Signaling Pathway on LPS-Treated BEAS-2B Bronchial Epithelial Cells. Int. Arch. Allergy Immunol. 2021, 182, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, D.; Tang, D.; Sun, K.; Peng, J.; Zhu, W.; Yin, S.; Wu, Y. Amygdalin Inhibits TGFbeta1-Induced Activation of Hepatic Stellate Cells (HSCs) in vitro and CCl4-induced Hepatic Fibrosis in Rats in vivo. Int. Immunopharmacol. 2021, 90, 107151. [Google Scholar] [CrossRef]
- Xiao, Z.; Ji, Q.; Fu, Y.D.; Gao, S.Q.; Hu, Y.H.; Liu, W.; Chen, G.F.; Mu, Y.P.; Chen, J.M.; Liu, P. Amygdalin Ameliorates Liver Fibrosis through Inhibiting Activation of TGF-beta/Smad Signaling. Chin. J. Integr. Med. 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Zhang, C.; Lin, J.; Zhen, C.; Wang, F.; Sun, X.; Kong, X.; Gao, Y. Amygdalin Protects Against Acetaminophen-Induced Acute Liver Failure by Reducing Inflammatory Response and Inhibiting Hepatocyte Death. Biochem. Biophys. Res. Commun. 2022, 602, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Al-Khafaji, K.; Taskin Tok, T. Molecular Dynamics Simulation, Free Energy Landscape and Binding Free Energy Computations in Exploration the Anti-Invasive Activity of Amygdalin Against Metastasis. Comput. Methods Programs Biomed. 2020, 195, 105660. [Google Scholar] [CrossRef] [PubMed]
- Syrigos, K.N.; Rowlinson-Busza, G.; Epenetos, A.A. In Vitro Cytotoxicity Following Specific Activation of Amygdalin by Β-Glucosidase Conjugated to A Bladder Cancer-Associated Monoclonal Antibody. Int. J. Cancer 1998, 78, 712–719. [Google Scholar] [CrossRef]
- Li, Y.-l.; Li, Q.-x.; Liu, R.-j.; Shen, X.-q.J.C.j.o.i.m. Chinese Medicine Amygdalin and Β-Glucosidase Combined with Antibody Enzymatic Prodrug System as A Feasible Antitumor Therapy. Chin. J. Integr. Med. 2018, 24, 237–240. [Google Scholar] [CrossRef]
- Makarević, J.; Tsaur, I.; Juengel, E.; Borgmann, H.; Nelson, K.; Thomas, C.; Bartsch, G.; Haferkamp, A.; Blaheta, R.A. Amygdalin Delays Cell Cycle Progression and Blocks Growth of Prostate Cancer Cells In Vitro. Life Sci. 2016, 147, 137–142. [Google Scholar] [CrossRef]
- Park, J.-H.; Seo, B.-l.; Cho, S.-Y.; Park, K.-R.; Choi, S.-H.; Han, C.-K.; Song, C.-H.; Park, S.-J.; Ku, S.-K. Single Oral Dose Toxicity Study of Prebrewed Armeniacae Semen in Rats. Toxicol. Res. 2013, 29, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mani, J.; Neuschafer, J.; Resch, C.; Rutz, J.; Maxeiner, S.; Roos, F.; Chun, F.K.; Juengel, E.; Blaheta, R.A. Amygdalin Modulates Prostate Cancer Cell Adhesion and Migration In Vitro. Nutr. Cancer 2020, 72, 528–537. [Google Scholar] [CrossRef]
- Makarević, J.; Rutz, J.; Juengel, E.; Kaulfuss, S.; Reiter, M.; Tsaur, I.; Bartsch, G.; Haferkamp, A.; Blaheta, R.A. Amygdalin Blocks Bladder Cancer Cell Growth in Vitro by Diminishing Cyclin A And Cdk2. PLoS ONE 2014, 9, e105590. [Google Scholar] [CrossRef] [PubMed]
- Makarević, J.; Rutz, J.; Juengel, E.; Kaulfuss, S.; Tsaur, I.; Nelson, K.; Pfitzenmaier, J.; Haferkamp, A.; Blaheta, R.A. Amygdalin Influences Bladder Cancer Cell Adhesion and Invasion in Vitro. PLoS ONE 2014, 9, e110244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abboud, M.M.; Al Awaida, W.; Alkhateeb, H.H.; Abu-Ayyad, A.N. Antitumor Action of Amygdalin on Human Breast Cancer Cells by Selective Sensitization to Oxidative Stress. Nutr. Cancer 2019, 71, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.-Y.; Kim, S.-W.; Yun, G.-M.; Lee, H.-S.; Kim, Y.-D.; Jeong, G.-J.; Ullah, I.; Rho, G.-J.; Jeon, B.-G. Inhibition of Cell Growth and Down-Regulation of Telomerase Activity by Amygdalin in Human Cancer Cell Lines. Anim. Cells Syst. 2015, 19, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Yanjun, L.; Tianwen, X.; Yongbin, Z.; Maolin, K.; Zhi, L.; Tao, H.; Daoda, C. Therapeutic Effect of Anti-Cea Mcab-Beta-Glucosidase Conjugate/Amygdalin System on Colorectal Cancer Xenografts in Nude Mice. Chin. Med. J. 2005, 29, 49–50. [Google Scholar]
- Genua, M.; Xu, S.-Q.; Buraschi, S.; Peiper, S.C.; Gomella, L.G.; Belfiore, A.; Iozzo, R.V.; Morrione, A. Proline-Rich Tyrosine Kinase 2 (PYK2) Regulates IGF-I-Induced Cell Motility and Invasion of Urothelial Carcinoma Cells. PLoS ONE 2012, 7, e40148. [Google Scholar] [CrossRef] [PubMed]
- Ay, C.; Yeh, C.-C.; Hsu, M.-C.; Hurng, H.-Y.; Kwok, P.C.L.; Chang, H.-I. Evaluation of the Correlation Between Focal Adhesion Kinase Phosphorylation and Cell Adhesion Force Using “Dep” Technology. Sensors 2012, 12, 5951–5965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, K.-C.; Hsu, S.-C.; Yang, J.-S.; Kuo, C.-L.; Ip, S.-W.; Lai, T.-Y.; Lin, S.-Y.; Huang, C.-C.; Kuo, S.-C.; Gibson Wood, W. 2-(3-Methoxyphenyl)-6, 7-Methylenedioxoquinolin-4-One, A Novel Synthetic Compound, Inhibited Migration and Invasion in Tsgh8301 Human Bladder Cancer Cells. Hum. Exp. Toxicol. 2011, 30, 1045–1052. [Google Scholar] [CrossRef]
- Green, T.P.; Fennell, M.; Whittaker, R.; Curwen, J.; Jacobs, V.; Allen, J.; Logie, A.; Hargreaves, J.; Hickinson, D.M.; Wilkinson, R.W. Preclinical Anticancer Activity of the Potent, Oral SRC Inhibitor AZD0530. Mol. Oncol. 2009, 3, 248–261. [Google Scholar] [CrossRef] [Green Version]
- Qian, L.; Xie, B.; Wang, Y.; Qian, J. Amygdalin-Mediated Inhibition of Non-Small Cell Lung Cancer Cell Invasion in Vitro. Int. J. Clin. Exp. Pathol. 2015, 8, 5363. [Google Scholar]
- Ayaz, Z.; Zainab, B.; Khan, S.; Abbasi, A.M.; Elshikh, M.S.; Munir, A.; Al-Ghamdi, A.A.; Alajmi, A.H.; Alsubaie, Q.D.; Mustafa, A. In Silico Authentication of Amygdalin as A Potent Anticancer Compound in the Bitter Kernels of Family Rosaceae. Saudi J. Biol. Sci. 2020, 27, 2444–2451. [Google Scholar] [CrossRef]
- Stock, C.C.; Martin, D.S.; Sugiura, K.; Fugmann, R.A.; Mountain, I.M.; Stockert, E.; Schmid, F.A.; Tarnowski, G.S. Antitumor Tests of Amygdalin in Spontaneous Animal Tumor Systems. J. Surg. Oncol. 1978, 10, 89–123. [Google Scholar] [CrossRef]
- Lee, H.M.; Moon, A. Amygdalin Regulates Apoptosis and Adhesion in HS578T Triple-Negative Breast Cancer Cells. Biomol. Ther. 2016, 24, 62. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Liu, J.; Zhang, M.; Wei, S.; Li, R.; Gao, Y.; Peng, W.; Wu, C. Apoptosis Induction of Fibroblast-Like Synoviocytes Is an Important Molecular-Mechanism for Herbal Medicine along with its Active Components in Treating Rheumatoid Arthritis. Biomolecules 2019, 9, 795. [Google Scholar] [CrossRef] [Green Version]
- Moradipoodeh, B.; Jamalan, M.; Zeinali, M.; Fereidoonnezhad, M.; Mohammadzadeh, G. In Vitro and In Silico Anticancer Activity of Amygdalin on The Sk-Br-3 Human Breast Cancer Cell Line. Mol. Biol. Rep. 2019, 46, 6361–6370. [Google Scholar] [CrossRef]
- Jumaa, A.H.; Hussein, S.M. Study The in Vitro Effect of Alcoholic Extract of Prunus Aremasia Kernels, Methotreaxte, Amygdalin and The Combination Between Them on Hela Cancer Cell Line. Iraqi J. Cancer Med. Genet. 2015, 8, 101–108. [Google Scholar]
- Shi, H.; Zheng, G.; Chen, N. Amygdalin Improves the Renal Function Via Anti-Oxidation and Anti-Fibrosis in Rats with Chronic Renal Failure. Panminerva. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Fang, Q.; Chen, G.; Wang, Z. Effects of Tetrandrine, Tetramethylpyrazine and Amygdalin on Human Kidney Fibroblast. Chin. J. Nephrol. 2000, 16, 186–189. [Google Scholar]
- Hong, C.F.; Lou, J.P.; Zhou, H.S.; He, L.Y.; Zhu, L.Q. The Study of Effects of Semen Persicae Extract on Silicotic Fibrosis in Rats. J. Lab. Med. 2000, 4, 218–219. [Google Scholar]
- Guo, J.; Sheng, M.; Wang, L.; Tan, J.; Wu, W.; Yang, S. Inhibitory Effect of Amygdalin on Human Renal Fibroblast Proliferation. J. Tissue Eng. Regen. Med. 2008, 12, 3575–3578. [Google Scholar]
- Du, H.K.; Song, F.C.; Zhou, X.; Li, H.; Zhang, J.P. Effect of Amygdlin on Type I and Iii Collagen Expression in Rats with Pulmonary Fibrosis Induced by Bleomycin. J. Chin. PLA Postgrad. Med. Sch. 2009, 30, 712–717. [Google Scholar]
- Guo, J.; Wu, W.; Sheng, M.; Yang, S.; Tan, J. Amygdalin Inhibits Renal Fibrosis in Chronic Kidney Disease. Mol. Med. Rep. 2013, 7, 1453–1457. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Li, L.; Tang, J.; Zhang, F.; Zhao, F.; Sun, D.; Zheng, F.; Wang, X. Amygdalin Inhibits HSC-T6 Cell Proliferation and Fibrosis Through the Regulation of TGF-β/CTGF. Mol. Cell. Toxicol. 2016, 12, 265–271. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, J.; Zhuo, Y.; Cui, L.; Li, C.; Cui, N.; Zhang, S. Amygdalin Improves Microcirculatory Disturbance and Attenuates Pancreatic Fibrosis by Regulating the Expression of Endothelin-1 and Calcitonin Gene-Related Peptide in Rats. J. Chin. Med. Assoc. 2018, 81, 437–443. [Google Scholar] [CrossRef]
- Zheng, Q.; Guo, Y.; Sun, J.; Xie, G.; Li, X.; Liao, G. An Experiment of Amygdalin Induced Premature Chromosome Condensation in Human Blood T Lymphocytes. J. Occup. Environ. Med. 2009, 26, 572–574. [Google Scholar]
- Dogru, H.Y.; Isguder, C.K.; Arici, A.; Ozsoy, A.Z.; Delibas, I.B.; Cakmak, B.J.P.b. Effect of Amygdalin on the Treatment and Recurrence of Endometriosis in an Experimental Rat Study. Period. Biol. 2017, 119, 173–180. [Google Scholar] [CrossRef]
- Jiagang, D.; Wang, H.; Liu, Y.; Li, C.; Hao, E.; Du, Z.; Bao, C.; Lv, J.; Wang, Y. Anti-atherosclerotic Effects Mediated by the Combination of Probucol and Amygdalin in Apolipoprotein E-Knockout Mice Fed with A High Fat Diet. J. Anim. Vet. Adv. 2012, 11, 20–25. [Google Scholar] [CrossRef]
- Moslehi, A.; Farahabadi, M.; Chavoshzadeh, S.A.; Barati, A.; Ababzadeh, S.; Mohammadbeigi, A. The Effect of Amygdalin on Endoplasmic Reticulum (Er) Stress Induced Hepatic Steatosis in Mice. Malays. J. Med. Sci. 2018, 25, 16. [Google Scholar] [CrossRef]
- Lv, J.; Xiong, W.; Lei, T.; Wang, H.; Sun, M.; Hao, E.; Wang, Z.; Huang, X.; Deng, S.; Deng, J. Amygdalin Ameliorates the Progression of Atherosclerosis in LDL Receptor-Deficient Mice. Mol. Med. Rep. 2017, 16, 8171–8179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halenar, M.; Medvedova, M.; Maruniakova, N.; Kolesarova, A. Assessment of A Potential Preventive Ability of Amygdalin in Mycotoxin-Induced Ovarian Toxicity. J. Environ. Sci. Health B 2015, 50, 411–416. [Google Scholar] [CrossRef]
- Tanyildizi, S.; Bozkurt, T. In Vitro Effects of Linamarin, Amygdalin and Gossypol Acetic Acid on Hyaluronidase Activity, Sperm Motility and Morphological Abnormality in Bull Sperm. Turk. J. Vet. Anim. Sci. 2004, 28, 819–824. [Google Scholar]
- Kamiński, P.; Baszyński, J.; Jerzak, I.; Kavanagh, B.P.; Nowacka-Chiari, E.; Polanin, M.; Szymański, M.; Woźniak, A.; Kozera, W. External and Genetic Conditions Determining Male Infertility. Int. J. Mol. Sci. 2020, 21, 5274. [Google Scholar] [CrossRef]
- Yasui, T.; Matsuzaki, T.; Ushigoe, K.; Kuwahara, A.; Maegawa, M.; Furumoto, H.; Aono, T.; Irahara, M. Stimulatory Effect of the Herbal Medicine Keishi-Bukuryo-Gan on A Cytokine-Induced Neutrophil Chemoattractant, in Rat Ovarian Cell Culture. Am. J. Reprod. Immunol. 2003, 50, 90–97. [Google Scholar] [CrossRef]
- Cheng, Y.; Yang, C.; Zhao, J.; Tse, H.F.; Rong, J. Proteomic Identification of Calcium-Binding Chaperone Calreticulin as A Potential Mediator for The Neuroprotective and Neuritogenic Activities of Fruit-Derived Glycoside Amygdalin. J. Nutr. Biochem. 2015, 26, 146–154. [Google Scholar] [CrossRef]
- Yang, C.; Zhao, J.; Cheng, Y.; Li, X.; Rong, J. Bioactivity-Guided Fractionation Identifies Amygdalin as A Potent Neurotrophic Agent from Herbal Medicine Semen Persicae Extract. Biomed. Res. Int. 2014, 2014, 306857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Sun, Y.; Wei, G.; Li, S.; Zhao, Z. Ergosterol Ameliorates Diabetic Nephropathy by Attenuating Mesangial Cell Proliferation and Extracellular Matrix Deposition Via The TGF-Β1/SMAD2 Signaling Pathway. Nutrients 2019, 11, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Desouky, M.A.; Fahmi, A.A.; Abdelkader, I.Y.; Nasraldin, K.M. Anticancer Effect of Amygdalin (Vitamin B-17) on Hepatocellular Carcinoma Cell Line (HepG2) in the Presence and Absence of Zinc. Anticancer Agents Med. Chem 2020, 20, 486–494. [Google Scholar] [CrossRef]
- He, X.Y.; Wu, L.J.; Wang, W.X.; Xie, P.J.; Chen, Y.H.; Wang, F. Amygdalin—A Pharmacological and Toxicological Review. J. Ethnopharmacol. 2020, 254, 112717. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Yan, L.; Shi, Z.; Wang, L.; Shan, L.; Efferth, T. Hepatoprotective and Anti-inflammatory Effects of Total Flavonoids of Qu Zhi Ke (Peel of Citrus Changshan-huyou) on Non-alcoholic Fatty Liver Disease in Rats via Modulation of NF-κB and MAPKs. Phytomedicine 2019, 64, 153082. [Google Scholar] [CrossRef]
- Boshtam, M.; Asgary, S.; Kouhpayeh, S.; Shariati, L.; Khanahmad, H. Aptamers Against Pro-and Anti-Inflammatory Cytokines: A Review. Inflammation 2017, 40, 340–349. [Google Scholar] [CrossRef]
- Yang, H.-Y.; Chang, H.-K.; Lee, J.-W.; Kim, Y.-S.; Kim, H.; Lee, M.-H.; Shin, M.-S.; Ham, D.-H.; Park, H.-K.; Lee, H.; et al. Amygdalin Suppresses Lipopolysaccharide-Induced Expressions of Cyclooxygenase-2 and Inducible Nitric Oxide Synthase in Mouse BV2 Microglial Cells. Neurol. Res. 2007, 29, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.-M.; Park, Y.-M.; Kim, I.-T.; Hong, S.-P.; Hong, J.-P.; Lee, K.-T. In vitro Anti-inflammatory Activity of Amygdalin in Murine Macrophage Raw 264.7 Cells. Korean J. Pharmacogn. 2003, 34, 223–227. [Google Scholar]
- Hwang, H.-J.; Lee, H.-J.; Kim, C.-J.; Shim, I.-S.; Hahm, D.-H. Inhibitory Effect of Amygdalin on Lipopolysaccharide-Inducible TNF-Alpha and Il-1beta mRNA Expression and Carrageenan-Induced Rat Arthritis. J. Microbiol. Biotechnol. 2008, 18, 1641–1647. [Google Scholar]
- Zhu, Y.; Su, Z.; Li, C. Analgesic Effect and No Physical Dependence of Amygdalin. Chin. J. Chin. Mat. Med. 1994, 19, 105–107, 128. [Google Scholar]
- Luo, D.; Shan, Z.; Jinlian, G.; Liu, Q.; Luo, L. Effects of Amygdalin on Tnf-A and Sicam-1 of Rats with Type II Collagen-Induced Arthritis. Chin. J. Inf. Tradit. Chin. Med. 2015, 12, 75–77. [Google Scholar]
- Zhang, A.; Pan, W.; Lv, J.; Wu, H. Protective Effect of Amygdalin on LPS-induced Acute lung injury by Inhibiting NF-κB and NLRP3 Signaling Pathways. Inflammation 2017, 40, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.Q.; Li, L.; Lu, C.J.; Lu, Y.; Wei, J.A.; Han, L. Anti-Inflammation Effect of Amygdalin on Macrophage 264.7 Cells Stimulated by Lipopolysaccharide. Trad. Chin. Drug Res. Clin. Pharmacol. 2018, 29, 257–263. [Google Scholar]
- Liczbiński, P.; Bukowska, B. Molecular Mechanism of Amygdalin Action In Vitro: Review of the Latest Research. Immunopharmacol. Immunotoxicol. 2018, 40, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Ying, J.; Ge, Q.; Hu, S.; Luo, C.; Lu, F.; Yu, Y.; Xu, T.; Lv, S.; Zhang, L.; Shen, J.; et al. Amygdalin Promotes Fracture Healing through TGF-beta/Smad Signaling in Mesenchymal Stem Cells. Stem Cells Int. 2020, 2020, 8811963. [Google Scholar] [CrossRef] [PubMed]
- Go, M.-R.; Kim, H.-J.; Yu, J.; Choi, S.-J. Toxicity and Toxicokinetics of Amygdalin in Maesil (Prunus Mume) Syrup: Protective Effect of Maesil Against Amygdalin Toxicity. J. Agric. Food Chem. 2018, 66, 11432–11440. [Google Scholar] [CrossRef]
- Hodgson, E. Toxicology and Human Environments; Academic Press: Cambridge, MA, USA, 2012; Volume 112. [Google Scholar]
- Akyildiz, B.N.; Kurtoğlu, S.; Kondolot, M.; Tunç, A. Cyanide Poisoning Caused by Ingestion of Apricot Seeds. Ann. Trop. Paediatr. 2010, 30, 39–43. [Google Scholar] [CrossRef]
- Parker-Cote, J.L.; Rizer, J.; Vakkalanka, J.P.; Rege, S.V.; Holstege, C.P. Challenges in the Diagnosis of Acute Cyanide Poisoning. Clin. Toxicol. 2018, 56, 609–617. [Google Scholar] [CrossRef]
- Rauws, A.G.; Olling, M.; Timmerman, A. The Pharmacokinetics of Prunasin, a Metabolite of Amygdalin. J. Toxicol. Clin. Toxicol. 1982, 19, 851–856. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.H.; McLafferty, M.A.; Goldman, P. Role of The Gastrointestinal Microflora in Amygdalin (Laetrile)-Induced Cyanide Toxicity. Biochem. Pharmacol. 1980, 29, 301–304. [Google Scholar] [CrossRef]
- Li, X.; Shi, F.; Gu, P.; Liu, L.; He, H.; Ding, L. A Sensitive LC–MS/MS Method for Simultaneous Determination of Amygdalin and Paeoniflorin in Human Plasma and its Application. J. Pharm. Biomed. Anal. 2014, 92, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Mani, J.; Rutz, J.; Maxeiner, S.; Juengel, E.; Bon, D.; Roos, F.; Chun, F.K.H.; Blaheta, R.A. Cyanide and Lactate Levels in Patients During Chronic Oral Amygdalin Intake Followed by Intravenous Amygdalin Administration. Complement. Ther. Med. 2019, 43, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Erikel, E.; Yuzbasioglu, D.; Unal, F. Genotoxic and Antigenotoxic Potential of Amygdalin on Isolated Human Lymphocytes by the Comet Assay. J. Food Biochem. 2020, 44, e13436. [Google Scholar] [CrossRef]
- Sarmento, B.; Ribeiro, A.; Veiga, F.; Ferreira, D.; Neufeld, R. Insulin-Loaded Nanoparticles are Prepared by Alginate Ionotropic Pre-Gelation Followed by Chitosan Polyelectrolyte Complexation. J. Nanosci. Nanotechnol. 2007, 7, 2833–2841. [Google Scholar] [CrossRef]
- De Jong, W.H.; Borm, P.J. Drug Delivery and Nanoparticles: Applications and Hazards. Int. J. Nanomed. 2008, 3, 133. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Jiao, Y.; Wang, Y.; Zhou, C.; Zhang, Z. Polysaccharides-Based Nanoparticles as Drug Delivery Systems. Adv. Drug Deliv. Rev. 2008, 60, 1650–1662. [Google Scholar] [CrossRef]
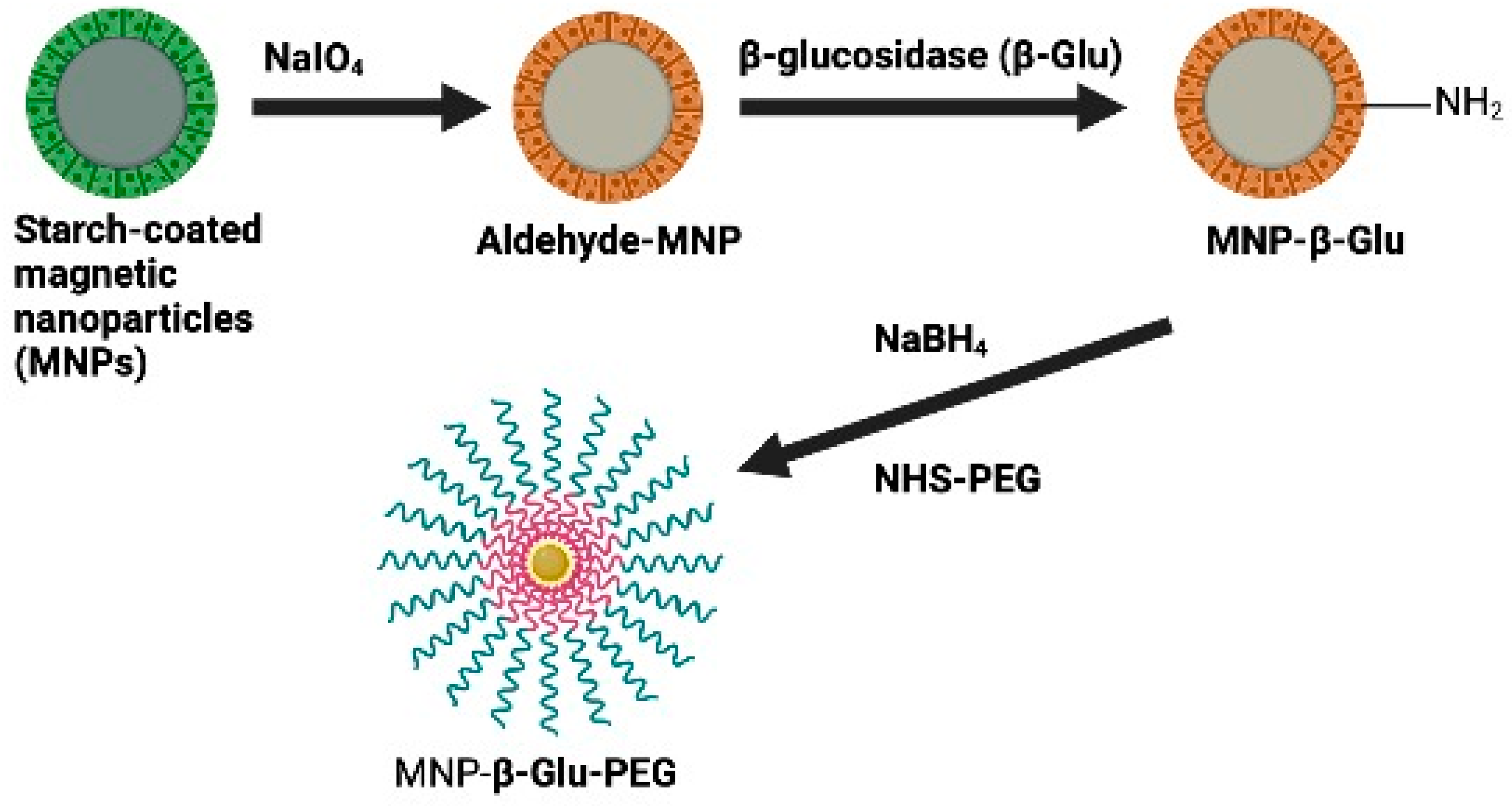
- Zhou, J.; Hou, J.; Rao, J.; Zhou, C.; Liu, Y.; Gao, W. Magnetically Directed Enzyme/Prodrug Prostate Cancer Therapy Based on beta-Glucosidase/Amygdalin. Int. J. Nanomed. 2020, 15, 4639–4657. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Dose/Method/Period | Activity | Refs. |
|---|---|---|---|
| KFB | 100 μg mL−1; culture; 48–72 h 80 μg mL−1; culture; 48–72 h 80 μg mL−1; culture; 24–48 h | Collagenase Activity Types ↓ Type-collagen ↓ Apoptosis of KFB ↑ | [98] |
| Tracheal injection of 50 mg/mL SiO2 suspension 1ml, SD rats | 48mg mL−1; i.p.; 1 month | Creating collagen in the lungs ↓ Blood ceruloplasmin ↓ | [99] |
| KFB | 100 mg L−1; culture; 48 h | Significantly reduces KFB cell proliferation. | [100] |
| Tracheal injection of 5 mg kg−1 bleomycin, Wistar rats | 15 mg kg−1 body weight; ip; 28 days | collagen types I and III ↓ | [101] |
| KFB | 25, 50, 100, 200, 400 and 800 μg mL−1; culture; 48 h | TGF-β1 ↓ KFB proliferation is dose-dependently inhibited. | [102] |
| Wistar rats undergo unilateral ureteral obstruction | 3 mg/kg, 5 mg kg−1; i.p.; 7,14 and 21 days | Renal interstitial lesion severity markedly decreased by day 21. | [102] |
| HTC-T6 cells | 200 μg mL−1; culture; 48 h and 72 h | TGF-β ↓ CTGF ↓ | [103] |
| HTC-T6 cells | 10−5, 10−4 and 10−3 mol L−1; culture; 48 h | PDGF ↓ IGF ↓ | [103] |
| CP model rats (injecting DBTC into the right caudal vein) | An intravenous injection of 10 milligrams per kilogram of body weight once daily for the first three days, then every other day for 28 days. | Pancreatic fibrosis, Acinar destruction, α-SMA, PDGF-BB, TGF β-1, and ET-1 ↓ CGRP ↓ | [104] |
| Model | Dose/Method/Period | Activity | Refs. |
|---|---|---|---|
| Glial cell type BV2 | 1, 10, 100, 1000 μg mL−1; culture; 24 h | COX-2 mRNA, iNOS mRNA the synthesis of prostaglandin E2 the production of nitric oxide | [121] |
| Cell line RAW 264.7 | 1, 10, 100 mmol L−1; culture; 6 h | At a concentration of 1 mM, TNF-α and IL-1β mRNA Amygdalin does not inhibit TNF-α and IL-1β mRNA expression in a dose-dependent manner. | [123] |
| SD male rats with Carrageenan-induced arthritis pain model | 0.005, 0.05 and 0.1 mg kg−1; im; 8 h | At a concentration of 0.005 mg/kg, Fos, TNF-α and IL-1β; However, no analgesic effect of amygdalin was observed at doses greater than 0.005 mg/kg | [123] |
| Modeling pain via plantar injection of formalin in SD male rats | 0.1, 0.5, 1.0 and 10.0 mg kg−1; Plantar injection | C-Fos, TNF-α, IL-1β Laetrile reduces pain in a dose-dependent manner in a dose range of less than 1 mg/kg. | [123] |
| Type II collagen-induced CIA model, Wistar rats | 120 mg kg−1; gavage; 28 days | TNF-α and sICAM-1 | [125] |
| The BALB/c Mice | 0.5, 1 and 2 mg kg−1; ip; 7 h | NF-κB Reduced pulmonary edema in a dose-dependent manner. | [126] |
| RAW264.7 cells | 6.25, 12.5, 25, 50, 100, 200, 400 µmol L−1; culture; 24 h | IL-17A, IL-23, CCL2 and CCL5 mRNA p-p38 the viability of RAW264.7 cell | [127] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barakat, H.; Aljutaily, T.; Almujaydil, M.S.; Algheshairy, R.M.; Alhomaid, R.M.; Almutairi, A.S.; Alshimali, S.I.; Abdellatif, A.A.H. Amygdalin: A Review on Its Characteristics, Antioxidant Potential, Gastrointestinal Microbiota Intervention, Anticancer Therapeutic and Mechanisms, Toxicity, and Encapsulation. Biomolecules 2022, 12, 1514. https://doi.org/10.3390/biom12101514
Barakat H, Aljutaily T, Almujaydil MS, Algheshairy RM, Alhomaid RM, Almutairi AS, Alshimali SI, Abdellatif AAH. Amygdalin: A Review on Its Characteristics, Antioxidant Potential, Gastrointestinal Microbiota Intervention, Anticancer Therapeutic and Mechanisms, Toxicity, and Encapsulation. Biomolecules. 2022; 12(10):1514. https://doi.org/10.3390/biom12101514
Chicago/Turabian StyleBarakat, Hassan, Thamer Aljutaily, Mona S. Almujaydil, Reham M. Algheshairy, Raghad M. Alhomaid, Abdulkarim S. Almutairi, Saleh I. Alshimali, and Ahmed A. H. Abdellatif. 2022. "Amygdalin: A Review on Its Characteristics, Antioxidant Potential, Gastrointestinal Microbiota Intervention, Anticancer Therapeutic and Mechanisms, Toxicity, and Encapsulation" Biomolecules 12, no. 10: 1514. https://doi.org/10.3390/biom12101514
APA StyleBarakat, H., Aljutaily, T., Almujaydil, M. S., Algheshairy, R. M., Alhomaid, R. M., Almutairi, A. S., Alshimali, S. I., & Abdellatif, A. A. H. (2022). Amygdalin: A Review on Its Characteristics, Antioxidant Potential, Gastrointestinal Microbiota Intervention, Anticancer Therapeutic and Mechanisms, Toxicity, and Encapsulation. Biomolecules, 12(10), 1514. https://doi.org/10.3390/biom12101514