
Knockdown of Two Trehalase Genes by RNA Interference Is Lethal to the White-Backed Planthopper Sogatella furcifera (Horváth) (Hemiptera: Delphacidae)
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Primer Design
2.3. Total RNA Extraction and cDNA Synthesis
2.4. SfTre1 and SfTre2 Gene Cloning
2.5. Sequence Analysis
2.6. Quantitative Real-Time PCR (qRT-PCR)
2.7. SfTre1 and SfTre2 Gene Silencing and Phenotypes
2.8. Statistical Analysis
3. Results
3.1. SfTre1 and SfTre2 Sequence Analysis
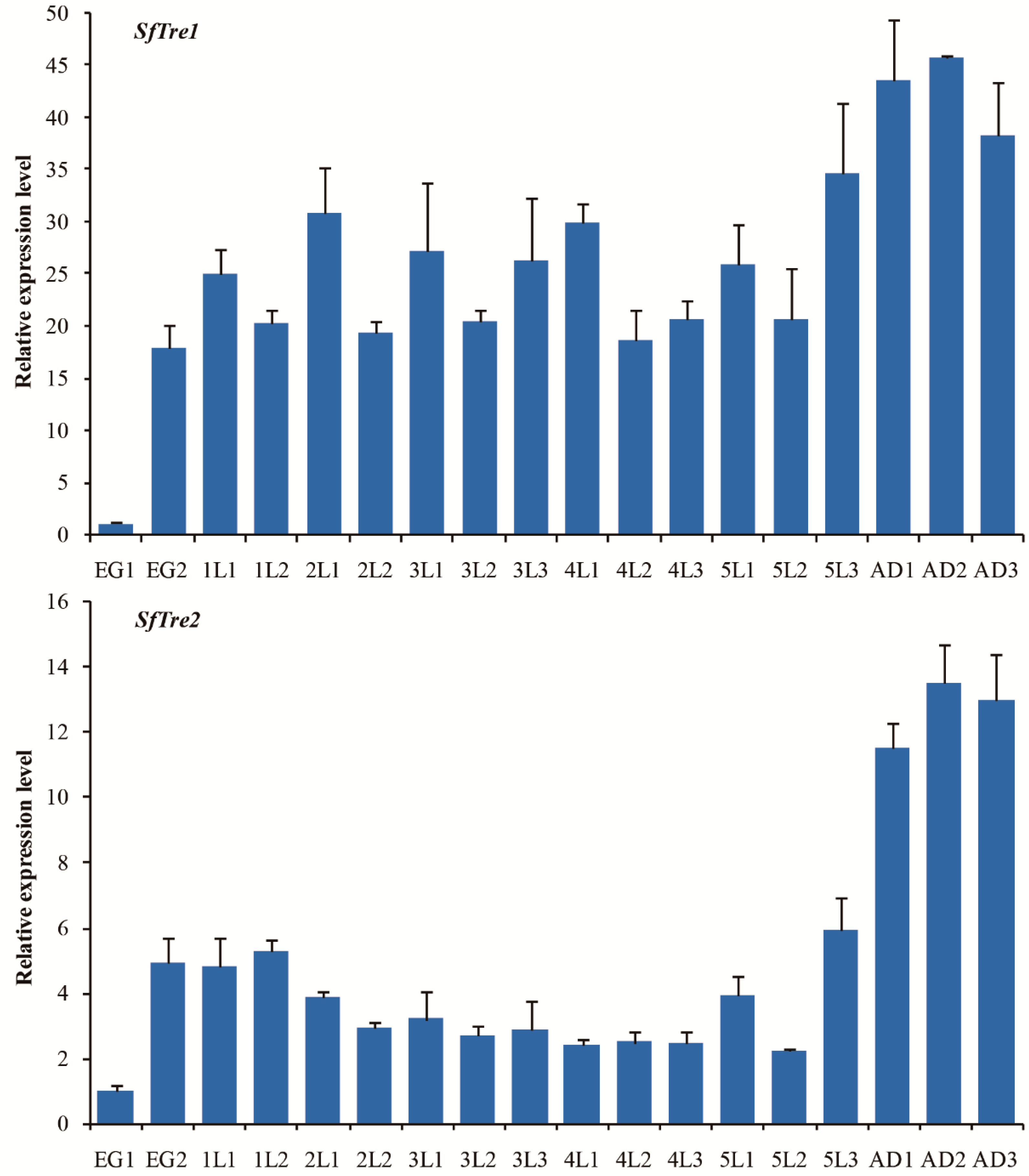
3.2. Spatiotemporal Expression of SfTre1 and SfTre2
3.3. Effects of RNAi
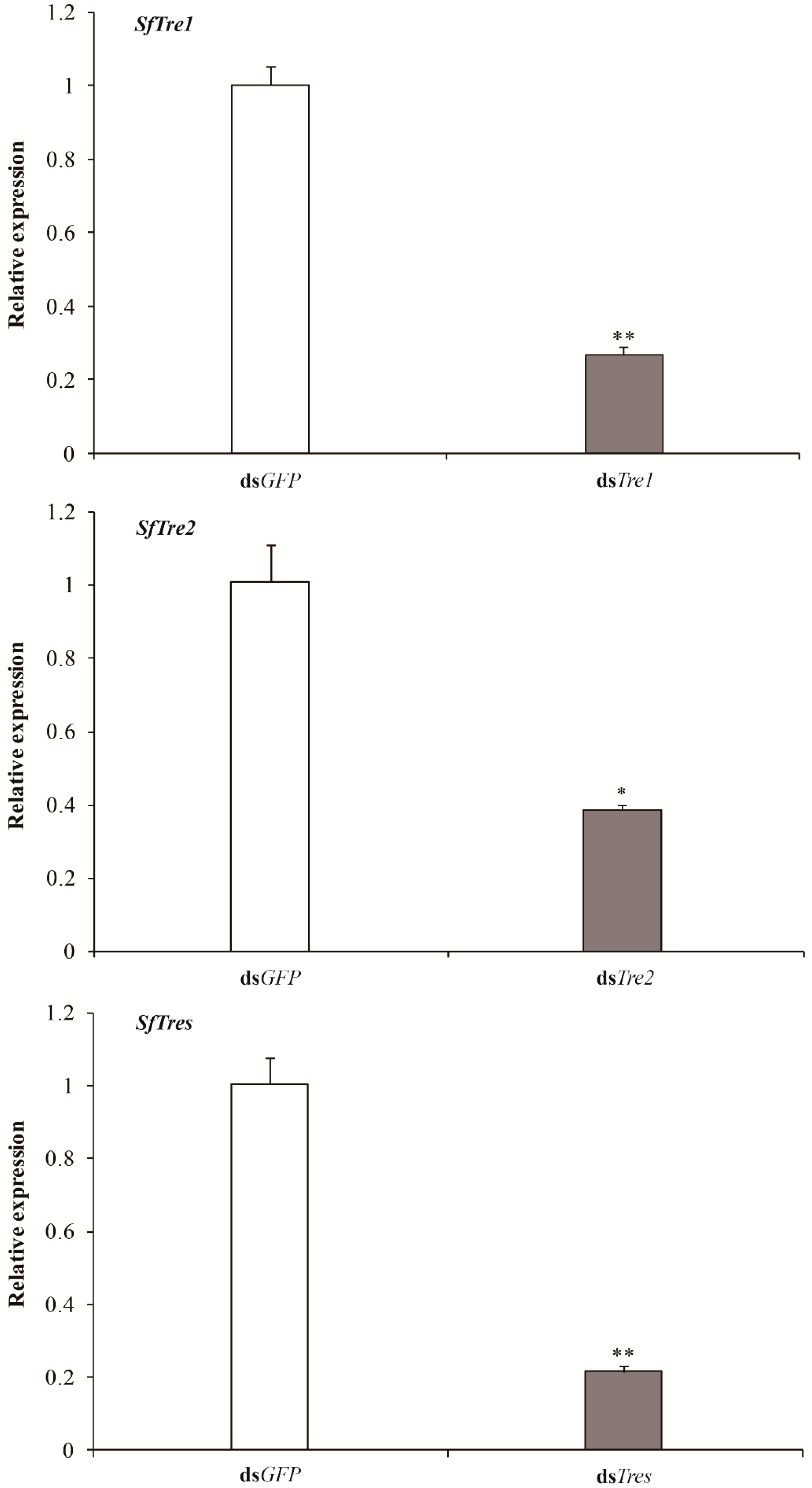
3.3.1. Effects of RNAi on the expression levels of SfTre1 and SfTre2
3.3.2. Effects of RNAi on the Survival Rates
3.3.3. Effects of RNAi on Phenotypes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhai, B.P. Rice planthoppers: A China problem under the international perspectives. Chin. J. Appl. Entomol. 2011, 48, 1184–1193. [Google Scholar]
- Zhu, Z.R.; Cheng, J.A. Sucking rates of the white-backed planthopper Sogatella furcifera (Horv.) (Homoptera: Delphacidae) and yield loss of rice. J. Pest Sci. 2002, 75, 113–117. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, C.; Long, G.Y.; Yang, H.; Jin, D.C. Sublethal effects of buprofezin on development, reproduction, and chitin synthase 1 gene (SfCHS1) expression in the white-backed planthopper, Sogatella furcifera (Hemiptera: Delphacidae). J. Asia-Pac. Entomol. 2018, 21, 585–591. [Google Scholar] [CrossRef]
- Huang, S.H.; Cheng, C.H.; Chen, C.N.; Wu, W.J.; Otuka, A. Estimating the immigration source of rice planthoppers, Nilaparvata lugens (Stål) and Sogatella furcifera (Horváth) (Homoptera: Delphacidae), in Taiwan. Appl. Entomol. Zool. 2010, 45, 521–531. [Google Scholar] [CrossRef] [Green Version]
- Zheng, D.B.; Hu, G.; Yang, F.; Du, X.D.; Yang, H.B.; Zhang, G.; Qi, G.J.; Liang, Z.L.; Zhang, X.X.; Cheng, X.N.; et al. Ovarian development status and population characteristics of Sogatella furcifera (Horváth) and Nilaparvata lugens (Stål): Implications for pest forecasting. J. Appl. Entomol. 2014, 138, 67–77. [Google Scholar] [CrossRef]
- Ali, E.; Liao, X.; Yang, P.; Mao, K.K.; Zhang, X.L.; Shakeel, M.; Salim, A.M.A.; Wan, H.; Li, J.H. Sublethal effects of buprofezin on development and reproduction in the white-backed planthopper, Sogatella furcifera (Hemiptera: Delphacidae). Sci. Rep. 2017, 7, 16913. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.H.; Wen, J.J.; Cai, D.J.; Li, P.; Xu, D.L.; Zhang, S.G. Southern rice black-streaked dwarf virus: A new proposed Fijivirus species in the family Reoviridae. Chin. Sci. Bull. 2008, 53, 3677–3685. [Google Scholar] [CrossRef]
- Matsukura, K.; Towata, T.; Yoshida, K.; Sakai, J.; Okuda, M.; Onuki, M.; Matsumura, M. Quantitative analysis of southern rice black-streaked dwarf virus in Sogatella furcifera and virus threshold for transmission. Phytopathology 2015, 105, 550–554. [Google Scholar] [CrossRef] [Green Version]
- Mao, K.K.; Ren, Z.J.; Li, W.B.; Liu, C.Y.; Xu, P.F.; He, S.; Li, J.H.; Wan, H. An insecticide resistance diagnostic kit for whitebacked planthopper Sogatella furcifera (Horváth). J. Pest Sci. 2021, 94, 531–540. [Google Scholar] [CrossRef]
- Abudulai, M.; Shepard, B.M.; Mitchell, P.L. Parasitism and predation on eggs of Leptoglossus phyllopus (Hemiptera: Coreidae) in cowpea: Impact of endosulfan sprays. Agric. Urban. Entomol. 2001, 18, 105–115. [Google Scholar]
- Jin, J.X.; Jin, D.C.; Li, W.H.; Cheng, Y.; Li, F.L.; Ye, Z.C. Monitoring trends in insecticide resistance of field populations of Sogatella furcifera (Hemiptera: Delphacidae) in Guizhou Province, China, 2012–2015. J. Econ. Entomol. 2017, 110, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E. Chitin synthesis and inhibition: A revisit. Pest Manag. Sci. 2001, 57, 946–950. [Google Scholar] [CrossRef] [PubMed]
- Elbein, A.D.; Pan, Y.T.; Pastuszak, I.; Carroll, D. New insights on trehalose: A multifunctional molecule. Glycobiology 2003, 13, 17R–27R. [Google Scholar] [CrossRef] [PubMed]
- Shukla, E.; Thorat, L.J.; Nath, B.B.; Gaikwad, S.M. Insect trehalase: Physiological significance and potential applications. Glycobiology 2015, 25, 357–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, B.Y.; Xu, K.K.; Luo, Y.J.; Wang, Y.Y.; Zhou, M.; Li, C.; Tang, B. The sequence characteristics and functions on regulating trehalose metabolism of two PTP genes in Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae). J. Stored Prod. Res. 2020, 89, 101692. [Google Scholar] [CrossRef]
- Huang, Q.T.; Zhang, G.J.; Nan, J.L.; Cheng, W.N.; Zhu-Salzman, K. Characterization of trehalose metabolic genes and corresponding enzymatic activities during diapause of Sitodiplosis mosellana. J. Insect Physiol. 2021, 135, 104324. [Google Scholar] [CrossRef]
- Wegener, G.; Tschiedel, V.; Schlöder, P.; Ando, O. The toxic and lethal effects of the trehalase inhibitor trehazolin in locusts are caused by hypoglycaemia. J. Exp. Biol. 2003, 206, 1233–1240. [Google Scholar] [CrossRef] [Green Version]
- Tatun, N.; Singtripop, T.; Sakurai, S. Dual control of midgut trehalase activity by 20-hydroxyecdysone and an inhibitory factor in the bamboo borer Omphisa fuscidentalis Hampson. J. Insect Physiol. 2008, 54, 351–357. [Google Scholar] [CrossRef]
- Ujita, M.; Yamanaka, M.; Maeno, Y.; Yoshida, K.; Ohshio, W.; Ueno, Y.; Banno, Y.; Fujii, H.; Okumura, H. Expression of active and inactive recombinant soluble trehalase using baculovirus-silkworm expression system and their glycan structures. J. Biosci. Bioeng. 2011, 111, 22–25. [Google Scholar] [CrossRef]
- Takiguchi, M.; Niimi, T.; Su, Z.H.; Yaginuma, T. Trehalase from male accessory gland of an insect, Tenebrio molitor. cDNA sequencing and developmental profile of the gene expression. Biochem. J. 1992, 288, 19–22. [Google Scholar] [CrossRef] [Green Version]
- Parkinson, N.M.; Conyers, C.M.; Keen, J.N.; Macnicoll, A.D.; Smith, I.; Weaver, R.J. cDNAs encoding large venom proteins from the parasitoid wasp Pimpla hypochondriaca identified by random sequence analysis. Comp. Biochem. Phys. C 2003, 134, 513–520. [Google Scholar] [CrossRef]
- Lee, J.H.; Saito, S.; Mori, H.; Nishimoto, M.; Okuyama, M.; Kim, D.; Wongchawalit, J.; Kimura, A.; Chiba, S. Molecular cloning of cDNA for trehalase from the European honeybee, Apis mellifera L. and its heterologous expression in Pichia pastoris. Biosci. Biotechnol. Biochem. 2007, 71, 2256–2265. [Google Scholar] [CrossRef] [PubMed]
- Mitsumasu, K.; Azuma, M.; Niimi, T.; Yamashita, O.; Yaginuma, T. Membrane-penetrating trehalase from silkworm Bombyx mori. Molecular cloning and localization in larval midgut. Insect Mol. Biol. 2005, 14, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.H.; Shao, Y.; Zhang, C.W.; Liu, Z.W.; Zhang, Y.J. Characterization of putative soluble and membrane-bound trehalases in a hemipteran insect, Nilaparvata lugens. J. Insect Physiol. 2009, 55, 997–1002. [Google Scholar] [CrossRef]
- Tang, B.; Qin, Z.; Shi, Z.K.; Wang, S.; Guo, X.J.; Wang, S.G.; Zhang, F. Trehalase in Harmonia axyridis (Coleoptera: Coccinellidae): Effects on beetle locomotory activity and the correlation with trehalose metabolism under starvation conditions. Appl. Entomol. Zool. 2014, 49, 255–264. [Google Scholar] [CrossRef]
- Tan, Y.A.; Xiao, L.B.; Zhao, J.; Xiao, Y.F.; Sun, Y.; Bai, L.X. Ecdysone receptor isoform-B mediates soluble trehalase expression to regulate growth and development in the mirid bug, Apolygus lucorum (Meyer-Dür). Insect Mol. Biol. 2015, 24, 611–623. [Google Scholar] [CrossRef]
- Shukla, E.; Thorat, L.J.; Bendre, A.D.; Jadhav, S.; Pal, J.K.; Nath, B.B.; Gaikwad, S.M. Cloning and characterization of trehalase: A conserved glycosidase from oriental midge, Chironomus ramosus. 3 Biotech 2018, 8, 352. [Google Scholar] [CrossRef]
- Tevatiya, S.; Kumari, S.; Sharma, P.; Rani, J.; Chauhan, C.; De, T.D.; Pandey, K.C.; Pande, V.; Dixit, R. Molecular and functional characterization of trehalase in the mosquito Anopheles stephensi. Front. Physiol. 2020, 11, 575718. [Google Scholar] [CrossRef]
- Santos, R.; Alves-Bezerra, M.; Rosas-Oliveira, R.; Majerowicz, D.; Meyer-Fernandes, J.R.; Gondim, K.C. Gene identification and enzymatic properties of a membrane-bound trehalase from the ovary of Rhodnius prolixus. Arch. Insect Biochem. 2012, 81, 199–213. [Google Scholar] [CrossRef]
- Zhang, Q.; Lu, D.H.; Pu, J.; Wu, M.; Han, Z.J. Cloning and RNA interference effects of trehalase genes in Laodelphax striatellus (Homoptera: Delphacidae). Acta Entomol. Sin. 2012, 55, 911–920. [Google Scholar]
- Li, Y.; Chen, X.; Wang, S.S.; Pan, B.Y.; Wang, S.G.; Wang, S.; Tang, B. Evaluation of the expression and function of the TRE2-like and TRE2 genes in ecdysis of Harmonia axyridis. Front. Physiol. 2019, 10, 1371. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.M.; Ding, J.H.; Jiang, D.L.; Liu, Z.X.; Li, Y.J.C.; Wang, J.; Wang, J.; Sheng, S.; Wu, F.A. Characterization and functional analysis of trehalase related to chitin metabolism in Glyphodes pyloalis Walker (Lepidoptera: Pyralidae). Insects 2021, 12, 370. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.J.; Sun, Y.W.; Cui, M.; Ma, E.B.; Zhang, J.Z. Molecular characteristics and functional analysis of trehalase genes in Locusta migratoria. Sci. Agric. Sin. 2016, 49, 4375–4386. [Google Scholar]
- Wang, Z.; Yang, H.; Zhou, C.; Yang, W.J.; Jin, D.C.; Long, G.Y. Molecular cloning, expression, and functional analysis of the chitin synthase 1 gene and its two alternative splicing variants in the white-backed planthopper, Sogatella furcifera (Hemiptera: Delphacidae). Sci. Rep. 2019, 9, 1087. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Long, G.Y.; Zhou, C.; Jin, D.C.; Yang, H.; Yang, W.J. Molecular characterization of UDP-N-acetylglucosamine pyrophosphorylase and its role in the growth and development of the white-backed planthopper Sogatella furcifera (Hemiptera: Delphacidae). Genes 2022, 13, 1340. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Yu, H.Z.; Wen, D.F.; Wang, W.L.; Geng, L.; Zhang, Y.; Xu, J.P. Identification of genes putatively involved in chitin metabolism and insecticide detoxification in the rice leaf folder (Cnaphalocrocis medinalis) larvae through transcriptomic analysis. Int. J. Mol. Sci. 2015, 16, 21873–21896. [Google Scholar] [CrossRef]
- Matsumura, M.; Sanada-Morimura, S.; Otuka, A.; Sonoda, S.; Van Thanh, D.; Van Chien, H.; Van Tuong, P.; Loc, P.M.; Liu, Z.W.; Zhu, Z.R.; et al. Insecticide susceptibilities of the two rice planthoppers Nilaparvata lugens and Sogatella furcifera in East Asia, the Red River Delta, and the Mekong Delta. Pest Manag. Sci. 2018, 74, 456–464. [Google Scholar] [CrossRef]
- Merzendorfer, H.; Zimoch, L. Chitin metabolism in insects: Structure, function and regulation of chitin synthases and chitinases. J. Exp. Biol. 2003, 206, 4393–4412. [Google Scholar] [CrossRef] [Green Version]
- Ullah, F.; Gul, H.; Wang, X.; Ding, Q.; Said, F.; Gao, X.W.; Desneux, N.; Song, D.L. RNAi-mediated knockdown of chitin synthase 1 (CHS1) gene causes mortality and decreased longevity and fecundity in Aphis gossypii. Insects 2020, 11, 22. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.D.; Liu, Y.K.; Qiu, L.Y.; Wang, S.S.; Pan, B.Y.; Li, Y.; Wang, S.G.; Tang, B. GFAT and PFK genes show contrasting regulation of chitin metabolism in Nilaparvata lugens. Sci. Rep. 2021, 11, 5246. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.H.; Mu, L.L.; Jin, L.; Anjum, A.A.; Li, G.Q. Silencing uridine diphosphate N-acetylglucosamine pyrophosphorylase gene impairs larval development in Henosepilachna vigintioctopunctata. Pest Manag. Sci. 2022, 78, 3894–3902. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zou, Z.W.; Xin, T.R.; Cai, S.Y.; Wang, X.; Zhang, H.J.; Zhong, L.; Xia, B. Knockdown of hexokinase in Diaphorina citri Kuwayama (Hemiptera: Liviidae) by RNAi inhibits chitin synthesis and leads to abnormal phenobypes. Pest Manag. Sci. 2022, 78, 4303–4313. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tang, B.; Chen, H.X.; Yao, Q.; Huang, X.F.; Chen, J.; Zhang, D.W.; Zhang, W.Q. Different functions of the insect soluble and membrane-bound trehalase genes in chitin biosynthesis revealed by RNA interference. PLoS ONE 2010, 5, e10133. [Google Scholar] [CrossRef]
- Tang, B.; Chen, X.F.; Liu, Y.; Tian, H.G.; Liu, J.; Hu, J.; Xu, W.H.; Zhang, W.Q. Characterization and expression patterns of a membrane-bound trehalase from Spodoptera exigua. BMC Mol. Biol. 2008, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- Tatun, N.; Singtripop, T.; Tungjitwitayakul, J.; Sakurai, S. Regulation of soluble and membrane-bound trehalase activity and expression of the enzyme in the larval midgut of the bamboo borer Omphisa fuscidentalis. Insect Biochem. Mol. 2008, 38, 788–795. [Google Scholar] [CrossRef]
- Zou, Q.; Wei, P.; Xu, Q.; Zheng, H.Z.; Tang, B.; Wang, S.G. cDNA cloning and characterization of two trehalases from Spodoptera litura (Lepidoptera: Noctuidade). Genet. Mol. Res. 2013, 12, 901–915. [Google Scholar] [CrossRef]
- Tan, Y.A.; Xiao, L.B.; Sun, Y.; Zhao, J.; Bai, L.X.; Xiao, Y.F. Molecular characterization of soluble and membrane-bound trehalases in the cotton mirid bug, Apolygus lucorum. Insect Biochem. Physiol. 2014, 86, 107–121. [Google Scholar] [CrossRef]
- Huber, R.E.; Lefebvre, Y.A. The Purification and some properties of soluble trehalase and sucrase from Drosophila melanogaster. Can. J. Biochem. 1971, 49, 1155–1164. [Google Scholar] [CrossRef]
- Huang, J.; Furusawa, T.; Sadakane, K.; Sugimura, Y. Purification and properties of two types of soluble trehalases from embryonic larvae of the silkworm, Bombyx mori. J. Insect Biotechnol. Sericol. 2006, 75, 1–8. [Google Scholar]
- Silva, M.C.P.; Ribeiro, A.F.; Terra, W.R.; Ferreira, C. Sequencing of Spodoptera frugiperda midgut trehalases and demonstration of secretion of soluble trehalase by midgut columnar cells. Insect Mol. Biol. 2009, 18, 769–784. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Dai, W.; Li, X.C.; Zhang, Y.L.; Zhang, C.N. Molecular cloning and expression analysis of soluble and membrane-bound trehalase genes in the cotton bollworm, Helicoverpa armigera. J. Asia-Pac. Entomol. 2015, 18, 187–195. [Google Scholar] [CrossRef]
- Yu, H.Z.; Huang, Y.L.; Lu, Z.J.; Zhang, Q.; Su, H.N.; Du, Y.M.; Yi, L.; Zhong, B.L.; Chen, C.X. Inhibition of trehalase affects the trehalose and chitin metabolism pathways in Diaphorina citri (Hemiptera: Psyllidae). Insect Sci. 2021, 28, 718–734. [Google Scholar] [CrossRef] [PubMed]
- Posnien, N.; Schinko, J.B.; Kittelmann, S.; Bucher, G. Genetics, development and composition of the insect head-a beetle’s view. Arthropod Struct. Dev. 2010, 39, 399–410. [Google Scholar] [CrossRef]
- Dzitoyeva, S.; Dimitrijevic, N.; Manev, H. Intra-abdominal injection of double-stranded RNA into anesthetized adult Drosophila triggers RNA interference in the central nervous system. Mol. Psychiatr. 2001, 6, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Tomoyasu, Y.; Miller, S.C.; Tomita, S.; Schoppmeier, M.; Grossmann, D.; Bucher, G. Exploring systemic RNA interference in insects: A genome-wide survey for RNAi genes in Tribolium. Genome Biol. 2008, 9, R10. [Google Scholar] [CrossRef] [Green Version]
- Walshe, D.P.; Lehane, S.M.; Lehane, M.J.; Haines, L.R. Prolonged gene knockdown in the tsetse fly Glossina by feeding double stranded RNA. Insect Mol. Biol. 2009, 18, 11–19. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Chandrashekar, K.; Thakur, N.; Verma, P.C.; Borgio, J.F.; Singh, P.K.; Tuli, R. RNA interference for the control of whiteflies (Bemisia tabaci) by oral route. J. Biosci. 2011, 36, 153–161. [Google Scholar] [CrossRef]
- Bhatia, V.; Bhattacharya, R. Host-mediated RNA interference targeting a cuticular protein gene impaired fecundity in the green peach aphid Myzus persicae. Pest Manag. Sci. 2018, 74, 2059–2068. [Google Scholar] [CrossRef]
- Wang, Q.T.; Fang, K.; Qi, L.Z.; Wang, X.; Pan, Y.; Li, Y.S.; Xi, J.H.; Zhang, J.H. Purification and functional characterization of a soluble trehalase in Lissorhoptrus oryzophilus (Coleoptera: Curculionidae). Insects 2022, 13, 867. [Google Scholar] [CrossRef]
- Zhao, L.N.; Yang, M.M.; Shen, Q.D.; Liu, X.J.; Shi, Z.K.; Wang, S.G.; Tang, B. Functional characterization of three trehalase genes regulating the chitin metabolism pathway in rice brown planthopper using RNA interference. Sci. Rep. 2016, 6, 27841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, B.; Wei, P.; Zhao, L.N.; Shi, Z.K.; Shen, Q.D.; Yang, M.M.; Xie, G.Q.; Wang, S.G. Knockdown of five trehalase genes using RNA interference regulates the gene expression of the chitin biosynthesis pathway in Tribolium castaneum. BMC Biotechnol. 2016, 16, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Gou, Y.P.; Guo, S.F.; Zhou, J.J.; Liu, C.Z. RAN interference of trehalose-6-phosphate synthase and trehalase genes regulates chitin metabolism in two color morphs of Acyrthosiphon pisum Harris. Sci. Rep. 2021, 11, 948. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Application Type | Primer Name | Primer Sequence (5′-3′) |
|---|---|---|---|
| SfTre1 | 5′ RACE | Tre1-R51 | CTGCCATTGTGCTGTCTCTTG |
| Tre1-R52 | AGTCACCTTCGCAATAGATG | ||
| Validation PCR | SfTre1-v-F | TCAAAATGTCGATTGGCGG | |
| SfTre1-v-R | CTCACACTTATCACGTACC | ||
| qRT-PCR | SfTre1-q-F | GACTTCTGCTATGTGATATGC | |
| SfTre1-q-R | GCTGTCCACCATCTGAATA | ||
| dsRNA synthesis | dsSfTre1-F | TAATACGACTCACTATAGGGCGGACTTCTGCTATGTGATA | |
| dsSfTre1-R | TAATACGACTCACTATAGGGAACGAACCATCTTGAACTGA | ||
| SfTre2 | PCR1 | SfTre2-F1 | ATGATGCTGAGGACTGAGA |
| SfTre2-R1 | ACTCTTCAACAATGGTCACA | ||
| PCR2 | SfTre2-F2 | AACTACTACGGAGAACTGAAG | |
| SfTre2-R2 | TGTGAGATTGTATGCCTTGT | ||
| 5′ RACE | Tre2-R51 | TTCCACAGTAGATTGAGTTG | |
| Tre2-R52 | CATTCTCAGTCCTCAGCATC | ||
| Validation PCR | SfTre2-v-F | TCCATCAAGCATGATAGC | |
| SfTre2-v-R | TACCCAGTTGTTCTAATCAT | ||
| qRT-PCR | SfTre2-q-F | GTGGTTGGATGCTGTTACTA | |
| SfTre2-q-R | GAGATGTTTGTCGGGTAGAA | ||
| dsRNA synthesis | dsSfTre2-F | TAATACGACTCACTATAGGGTACTGTTGCTGTTGGTGTTA | |
| dsSfTre2-R | TAATACGACTCACTATAGGGCGTCTTCACATCATCCTTCA | ||
| 18S rRNA | qRT-PCR | Sf18S-q-F | CGGAAGGATTGACAGATTGAT |
| Sf18S-q-R | CACGATTGCTGATACCACATAC | ||
| GFP | dsRNA synthesis | dsGFP-F | TAATACGACTCACTATAGGGAAGGGCGAGGAGCTGTTCACCG |
| dsGFP-R | TAATACGACTCACTATAGGGCAGCAGGACCATGTGATCGCGC |
| Gene | Length of cDNA (bp) | Number of Amino Acids | Molecular Weight (kDa) | Isoelectric Point | Accession Number |
|---|---|---|---|---|---|
| SfTre1 | 3700 | 575 | 67.01 | 5.44 | MG869613 |
| SfTre2 | 2757 | 633 | 72.82 | 5.65 | MG869614 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Long, G.-Y.; Jin, D.-C.; Yang, H.; Zhou, C.; Yang, X.-B. Knockdown of Two Trehalase Genes by RNA Interference Is Lethal to the White-Backed Planthopper Sogatella furcifera (Horváth) (Hemiptera: Delphacidae). Biomolecules 2022, 12, 1699. https://doi.org/10.3390/biom12111699
Wang Z, Long G-Y, Jin D-C, Yang H, Zhou C, Yang X-B. Knockdown of Two Trehalase Genes by RNA Interference Is Lethal to the White-Backed Planthopper Sogatella furcifera (Horváth) (Hemiptera: Delphacidae). Biomolecules. 2022; 12(11):1699. https://doi.org/10.3390/biom12111699
Chicago/Turabian StyleWang, Zhao, Gui-Yun Long, Dao-Chao Jin, Hong Yang, Cao Zhou, and Xi-Bin Yang. 2022. "Knockdown of Two Trehalase Genes by RNA Interference Is Lethal to the White-Backed Planthopper Sogatella furcifera (Horváth) (Hemiptera: Delphacidae)" Biomolecules 12, no. 11: 1699. https://doi.org/10.3390/biom12111699
APA StyleWang, Z., Long, G. -Y., Jin, D. -C., Yang, H., Zhou, C., & Yang, X. -B. (2022). Knockdown of Two Trehalase Genes by RNA Interference Is Lethal to the White-Backed Planthopper Sogatella furcifera (Horváth) (Hemiptera: Delphacidae). Biomolecules, 12(11), 1699. https://doi.org/10.3390/biom12111699