
Folic Acid, Folinic Acid, 5 Methyl TetraHydroFolate Supplementation for Mutations That Affect Epigenesis through the Folate and One-Carbon Cycles



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
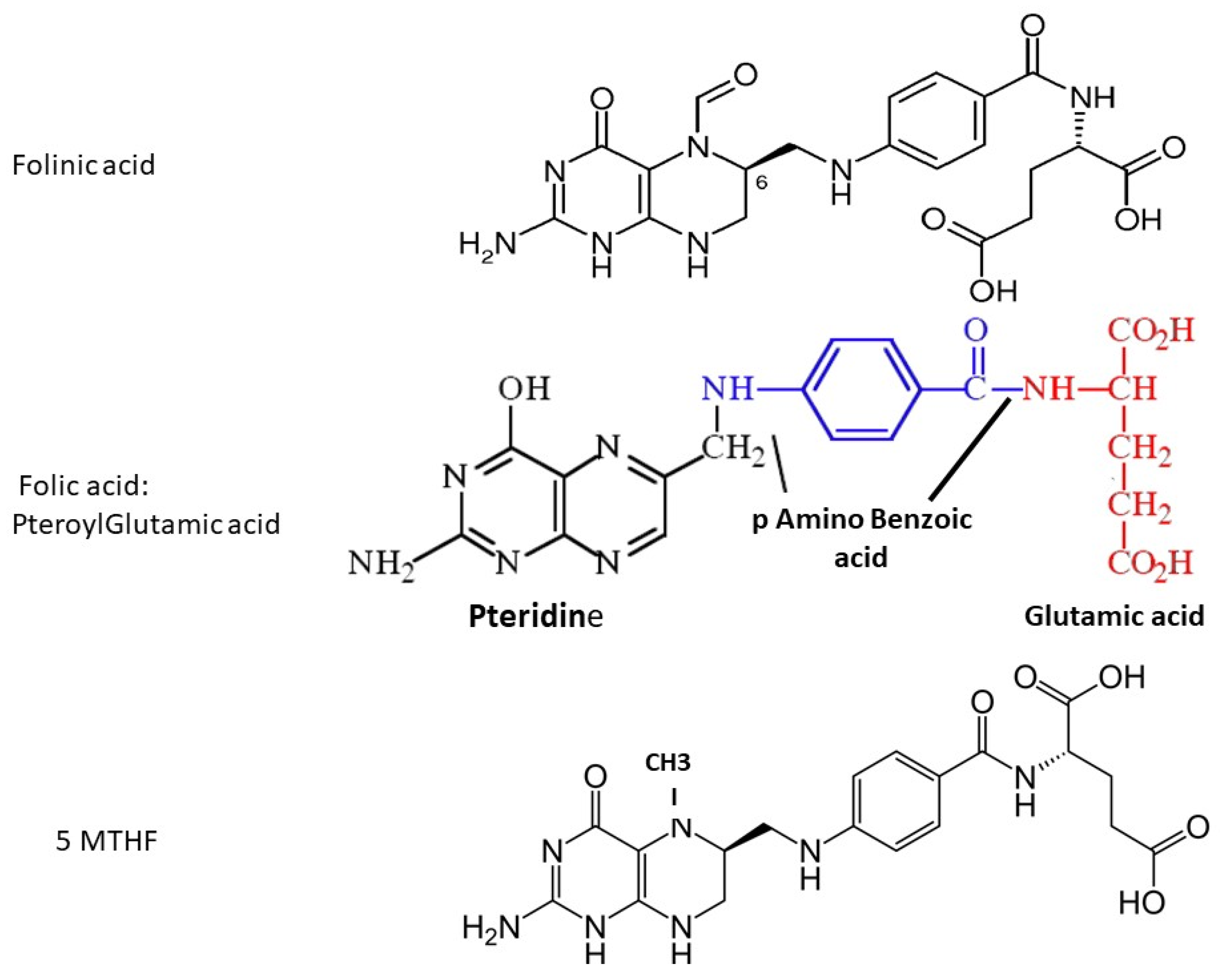
2. The Folate Family
Folate Levels and Folate Assays
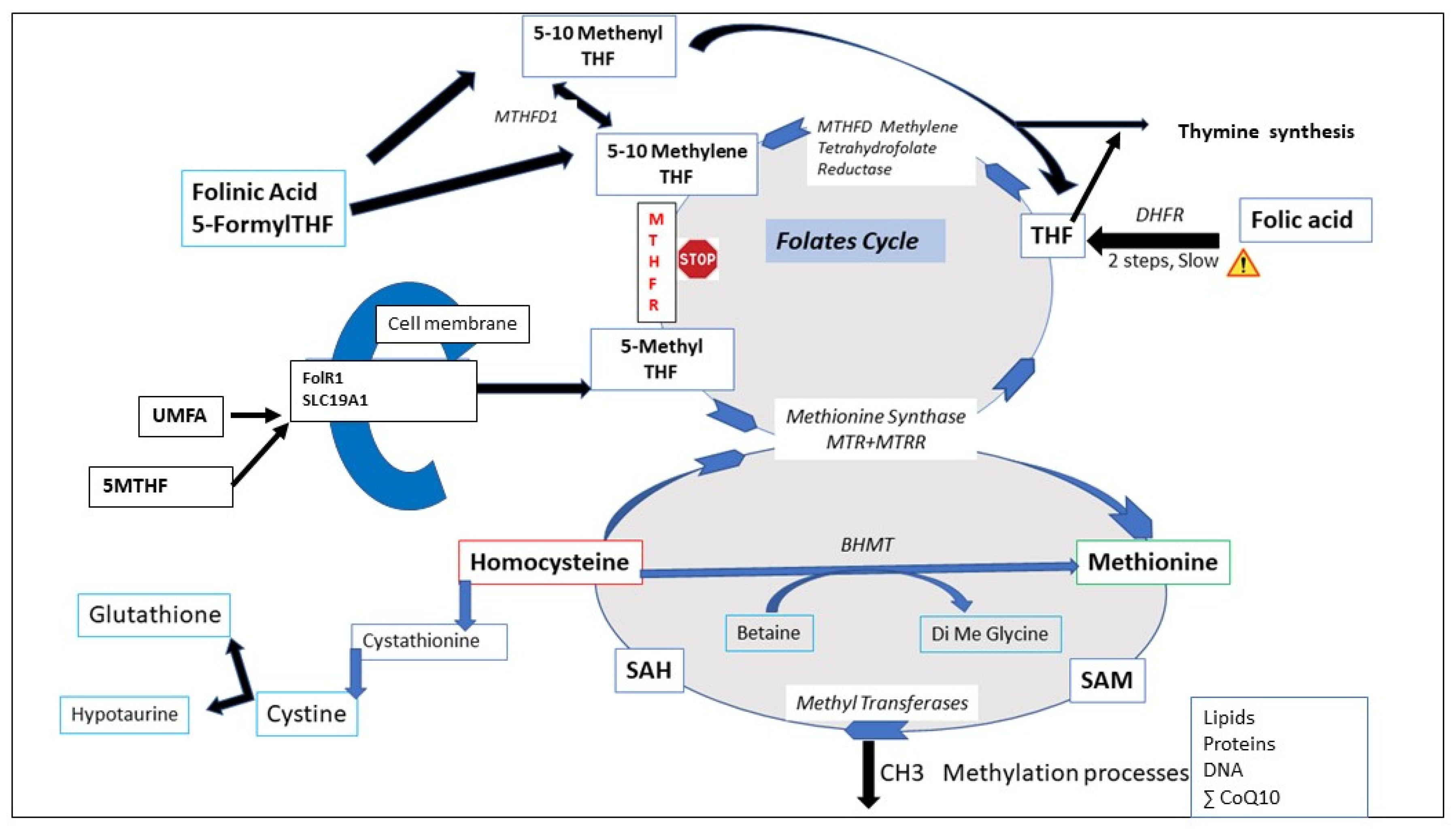
3. The One Carbon (1-CC) and the Folates Cycle (FC)
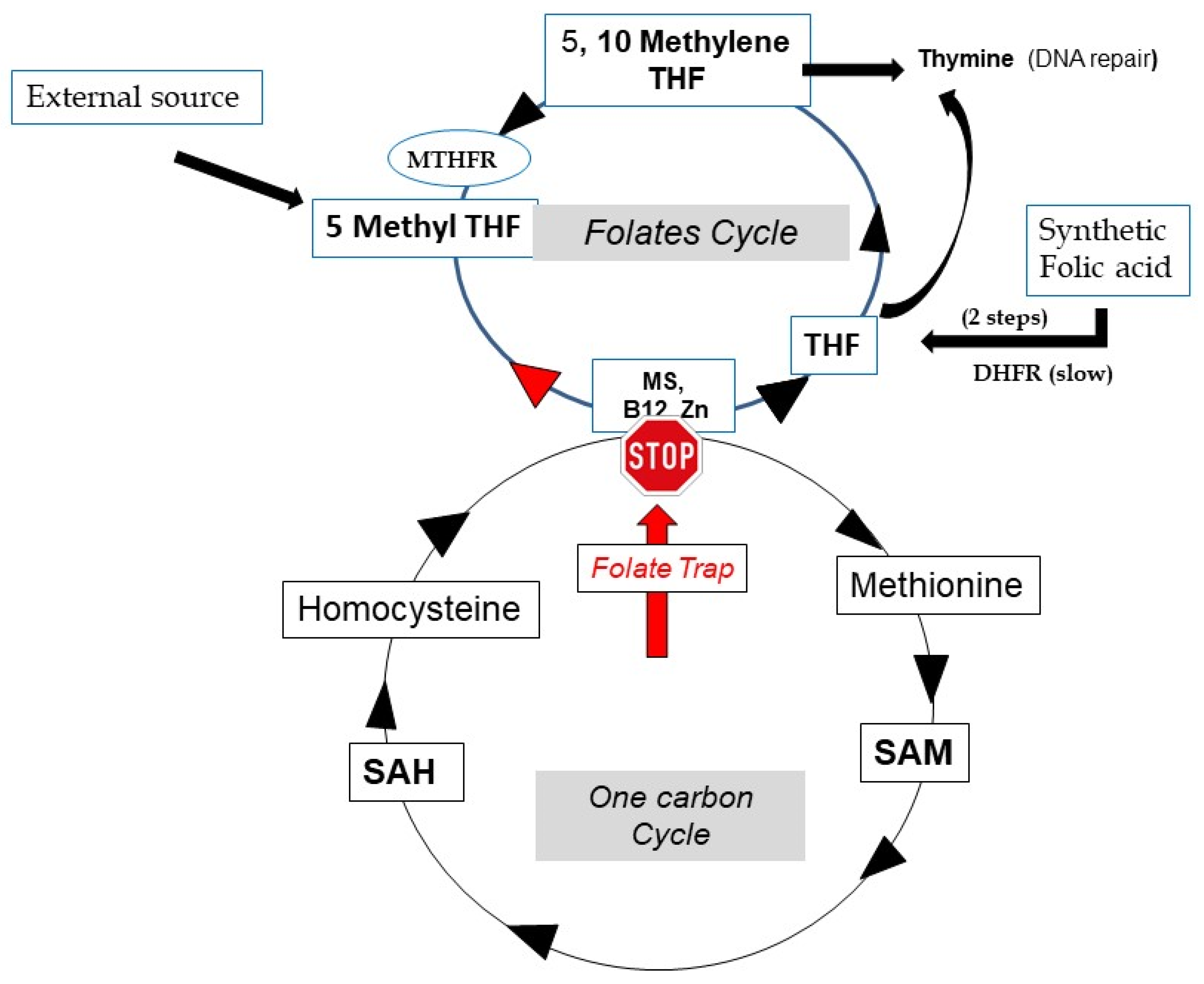
3.1. The Folate Trap
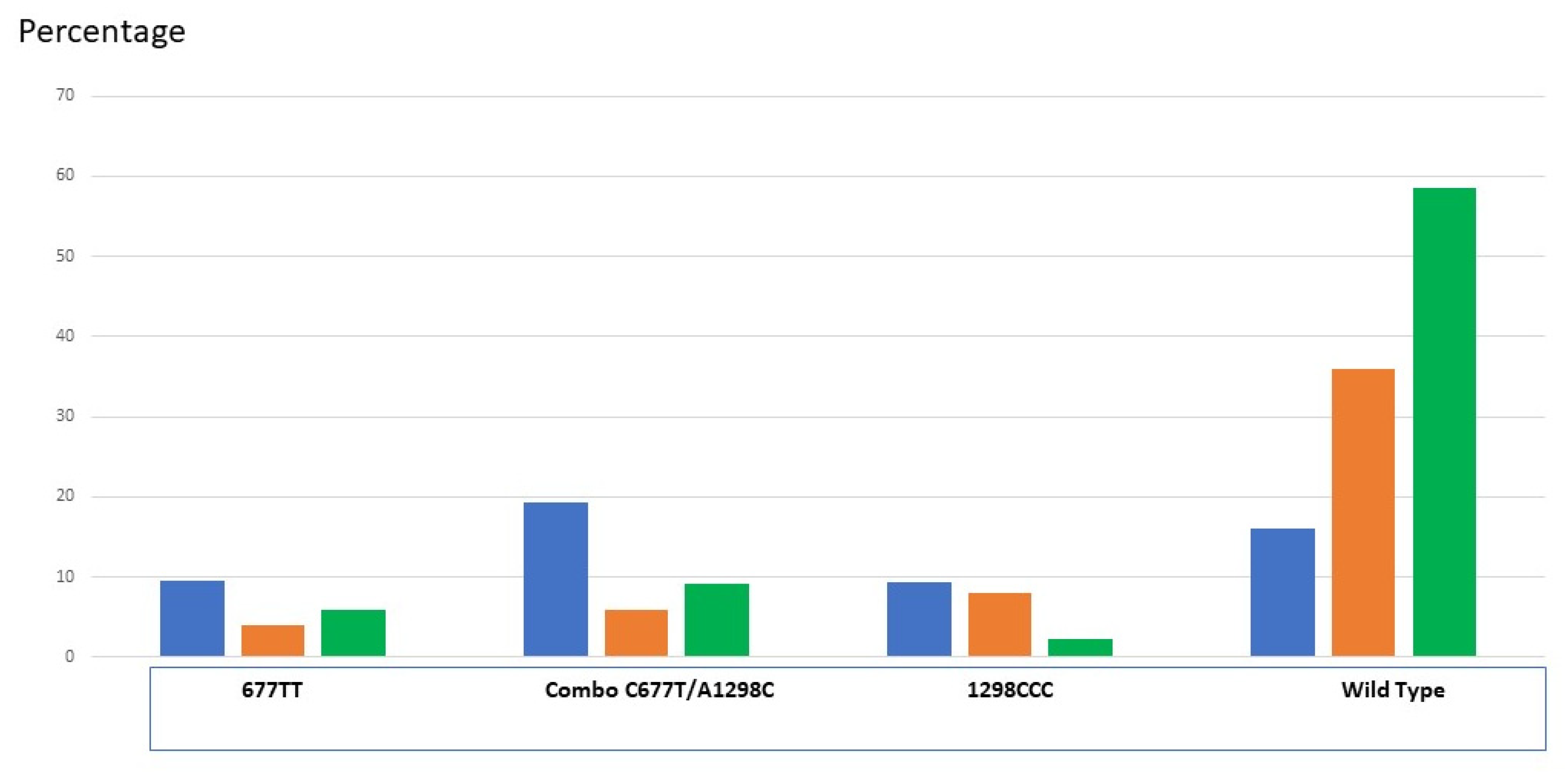
3.2. 3-MTHFR Variants
4. Metabolism of Synthetic Folic Acid (Pteroylglutamic Acid, FA) and the UMFA Syndrome
5. Folinic Acid (5 Formyl THF, FLA)
6. L MethylFolate (5-MTHF)
7. Folate Malabsorption
7.1. Pathologies Associated with Folate Metabolism
7.2. FA, FLA and Homocysteine
7.3. FA vs. 5-MTHF
7.4. Is FA Beneficial for Pathologies Associated with Folate Deficiency?
7.5. FA, UMFA and Risk of Cancer
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Duthie, S.; Narayanan, S.; Brand, G.; Pirie, L.; Grant, G. Impact of folate deficiency on DNA stability. J. Nutr. 2002, 132 (Suppl. 8), 2444S–2449S. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.; Ayling, J.E. The extremely slow and variable activity of dihydrofolate reductase in human liver and its implications for high folic acid intake. Proc. Natl. Acad. Sci. USA 2009, 106, 15424–15429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, M.R.; McPartlin, J.; Weir, D.G.; Daly, S.; Pentieva, K.; Daly, L.; Scott, J.M. Evidence of unmetabolised folic acid in cord blood of newborn and serum of 4-day-old infants. Br. J. Nutr. 2005, 94, 727–730. [Google Scholar] [CrossRef]
- Sobczyńska-Malefora, A.; Harrington, D.J. Laboratory assessment of folate (vitamin B9) status. J. Clin. Pathol. 2018, 71, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Zuhra, K.; Augsburger, F.; Majtan, T.; Szabo, C. Cystathionine-β-Synthase: Molecular Regulation and Pharmacological Inhibition. Biomolecules 2020, 10, 697. [Google Scholar] [CrossRef]
- Menezo, Y.; Silvestris, E.; Dale, B. Elder K Oxidative stress and alterations in DNA methylation: Two sides of the same coin in reproduction. Reprod. Biomed. Online 2016, 33, 668–683. [Google Scholar] [CrossRef] [Green Version]
- Botto, L.D.; Yang, Q. 5,10-Methylenetetrahydrofolate reductase gene variants and congenital anomalies: A HuGE review. Am. J. Epidemiol. 2000, 151, 862–877. [Google Scholar] [CrossRef]
- van der Put, N.M.; Gabreëls, F.; Stevens, E.M.; Smeitink, J.A.; Trijbels, F.J.; Eskes, T.K.; van den Heuvel, L.P.; Blom, H.J. A second common mutation in the methylenetetrahydrofolate reductase gene: An additional risk factor for neural-tube defects? Am. J. Hum. Genet. 1998, 62, 1044–1051. [Google Scholar] [CrossRef] [Green Version]
- Tsang, B.L.; Devine, O.J.; Cordero, A.M.; Marchetta, C.M.; Mulinare, J.; Mersereau, P.; Guo, J.; Qi, Y.P.; Berry, R.J.; Rosenthal, J.; et al. Assessing the association between the methylenetetrahydrofolate reductase (MTHFR) 677C>T polymorphism and blood folate concentrations: A systematic review and meta-analysis of trials and observational studies. Am. J. Clin. Nutr. 2015, 101, 1286–1294. [Google Scholar] [CrossRef]
- Ménézo, Y.; Patrizio, P.; Alvarez, S.; Amar, E.; Brack, M.; Brami, C.; Chouteau, J.; Clement, A.; Clement, P.; Cohen, M.; et al. MTHFR (methylenetetrahydrofolate reductase: EC 1.5.1.20) SNPs (single-nucleotide polymorphisms) and homocysteine in patients referred for investigation of fertility. J. Assist. Reprod. Genet. 2021, 38, 2383–2389. [Google Scholar] [CrossRef]
- Aguilar-Lacasaña, S.; López-Flores, I.; González-Alzaga, B.; Giménez-Asensio, M.J.; Carmona, F.D.; Hernández, A.F.; López Gallego, M.F.; Romero-Molina, D.; Lacasaña, M. Methylenetetrahydrofolate Reductase (MTHFR) Gene Polymorphism and Infant’s Anthropometry at Birth. Nutrients 2021, 13, 831. [Google Scholar] [CrossRef] [PubMed]
- Camerota, M.; Graw, S.; Everson, T.M.; McGowan, E.C.; Hofheimer, J.A. Prenatal risk factors and neonatal DNA methylation in very preterm infants. Clin. Epigenetics 2021, 13, 171. [Google Scholar] [CrossRef]
- Bleich, S.; Semmler, A.; Frieling, H.; Thumfart, L.; Muschler, M.; Hillemacher, T.; Kornhuber, J.; Kallweit, U.; Simon, M.; Linnebank, M. Genetic variants of methionine metabolism and DNA methylation. Epigenomics 2014, 6, 585–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, M.R.; Staines, A.; Daly, L.; Traynor, A.; Daly, S.; Bailey, S.W.; Alverson, P.B.; Ayling, J.E.; Scott, J.M. Persistent circulating unmetabolised folic acid in a setting of liberal voluntary folic acid fortification. Implications for further mandatory fortification? BMC Public Health 2009, 9, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGowan, E.C.; Hong, X.; Selhub, J.; Paul, L.; Wood, R.A.; Matsui, E.C.; Keet, C.A.; Wang, X. Association Between Folate Metabolites and the Development of Food Allergy in Children. J. Allergy Clin. Immunol. Pract. 2020, 8, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Chen, Y.; Pei, J.; Pan, J. MTHFD1 gene polymorphisms as risk factors involved in orofacial cleft: An independent case-control study and a meta-analysis. Int. J. Clin. Exp. Med. 2015, 8, 7737–7745. [Google Scholar]
- Cornet, D.; Clement, A.; Clement, P.; Menezo, Y. High doses of folic acid induce a pseudo-methylenetetrahydrofolate syndrome. SAGE Open Med. Case Rep. 2019, 7, 2050313X1985043. [Google Scholar] [CrossRef]
- Christensen, K.E.; Mikael, L.G.; Leung, K.Y.; Lévesque, N.; Deng, L.R. High folic acid consumption leads to pseudo-MTHFR deficiency, altered lipid metabolism, and liver injury in mice. Am. J. Clin. Nutr. 2015, 101, 646–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidmar Golja, M.; Šmid, A.; Karas Kuželički, N.; Trontelj, J.; Geršak, K.; Mlinarič-Raščan, I. Folate Insufficiency Due to MTHFR Deficiency Is Bypassed by 5-Methyltetrahydrofolate. J. Clin. Med. 2020, 9, 2836. [Google Scholar] [CrossRef]
- Steinfeld, R.; Grapp, M.; Kraetzner, R.; Dreha-Kulaczewski, S.; Helms, G.; Dechent, P.; Wevers, R.; Grosso, S.; Gärtner, J. Folate receptor alpha defect causes cerebral folate transport deficiency: A treatable neurodegenerative disorder associated with disturbed myelin metabolism. Am. J. Hum. Genet. 2009, 85, 354–363. [Google Scholar] [CrossRef] [Green Version]
- Škovierová, H.; Vidomanová, E.; Mahmood, S.; Sopková, J.; Drgová, A.; Červeňová, T.; Halašová, E.; Lehotský, J. The Molecular and Cellular Effect of Homocysteine Metabolism Imbalance on Human Health. Int. J. Mol. Sci. 2016, 17, 1733. [Google Scholar] [CrossRef] [PubMed]
- Laanpere, M.; Altmäe, S.; Stavreus-Evers, A.; Nilsson, T.K.; Yngve, A.; Salumets, A. Folate-mediated one-carbon metabolism and its effect on female fertility and pregnancy viability. Nutr. Rev. 2010, 68, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Boxmeer, J.C.; Steegers-Theunissen, R.P.; Lindemans, J.; Wildhagen, M.F.; Martini, E.; Steegers, E.A.; Macklon, N.S. Homocysteine metabolism in the pre-ovulatory follicle during ovarian stimulation. Hum. Reprod. 2008, 23, 2570–2576. [Google Scholar]
- Kovács, T.; Szabó-Meleg, E.; Ábrahám, I.M. stradiol-Induced Epigenetically Mediated Mechanisms and Regulation of Gene Expression. Int. J. Mol. Sci. 2020, 21, 3177. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, A.; Usefzadeh, M.; Mianehsaz, E.; Foroozanfard, F.; Nikoueinejad, H.; Moraveji, S.A.; Nasiri, O.; Rajali, M. Comparison of oral folic acid and folinic acid on blood homocysteine level of patients on hemodialysis. Iran J. Kidney Dis. 2011, 5, 45–49. [Google Scholar]
- Hauser, A.C.; Hagen, W.; Rehak, P.H.; Buchmayer, H.; Födinger, M.; Papagiannopoulos, M.; Bieglmayer, C.; EKöller, E.; Ignatescu, M.; Hörl, W.H.; et al. Efficacy of folinic versus folic acid for the correction of hyperhomocysteinemia in hemodialysis patients. Am. J. Kidney Dis. 2001, 37, 758–765. [Google Scholar] [CrossRef]
- Hogeveen, M.; den Heijer, M.; Schonbeck, Y.; Ijland, M.; van Oppenraaij, D.; Gunnewiek, J.K.; Blom, H. The effect of folinic acid supplementation on homocysteine concentrations in newborns. Eur. J. Clin. Nutr. 2010, 64, 1266–1271. [Google Scholar] [CrossRef] [Green Version]
- Niederberger, K.E.; Dahms, I.; Broschard, T.H.; Boehni, R.; Moser, R. Safety evaluation of calcium L-methylfolate. Toxicol. Rep. 2019, 6, 1018–1030. [Google Scholar] [CrossRef]
- Venn, B.J.; Green, T.J.; Moser, R.; Mann, J.I. Comparison of the effect of low-dose supplementation with L-5-methyltetrahydrofolate or folic acid on plasma homocysteine: A randomized placebo-controlled study. Am. J. Clin. Nutr. 2003, 77, 658–662. [Google Scholar] [CrossRef] [Green Version]
- Troesch, B.; Demmelmair, J.; Gimpfl, M.; Hecht, C.; Lakovic, G.; Roehle, R.; Sipka, L.; Trisic, B.; Vusurovic, M.; Schoop, R.; et al. MEFOLIN Study Group. Suitability and safety of L-5- methyltetrahydrofolate as a folate source in infant formula: A randomized-controlled trial. PLoS ONE 2019, 14, e0216790. [Google Scholar] [CrossRef] [Green Version]
- Prinz-Langenoh, R.; Bramswig, S.; Tobolski, O.; Smulders, Y.M.; Smith, D.E.; Finglas, P.M.; Pietrzik, K. 5-MTHF increases plasma folate more effectively than folic acid in women with the homozygous or wild-type6777C ->T polymorphism of MTHFR. Br. J. Pharmacol. 2009, 158, 2014–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietrzik, K.; Bailey, L.; Shane, B. Folic acid and L-5-methyltetrahydrofolate: Comparison of clinical pharmacokinetics and pharmacodynamics. Clin. Pharmacokinet. 2010, 49, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Obeid, R.; Holzgreve, W.; Pietrzik, K. Is 5-methyltetrahydrofolate an alternative to folic acid for the prevention of neural tube defects? J. Perinat. Med. 2013, 41, 469–483. [Google Scholar] [CrossRef] [PubMed]
- Henderson, A.M.; Aleliunas, R.E.; Loh, S.P.; Khor, G.L.; Harvey-Leeson, S. 5-Methyltetrahydrofolate Supplementation Increases Blood Folate Concentrations to a Greater Extent than Folic Acid Supplementation in Malaysian Women. J. Nutr. 2018, 148, 885–890. [Google Scholar] [CrossRef] [Green Version]
- Cochrane, K.M.; Mayer, C.; Devlin, A.M.; Elango, R.; Hutcheon, J.A.; Karakochuk, C.D. Is natural (6S)-5-methyltetrahydrofolic acid as effective as synthetic folic acid in increasing serum and red blood cell folate concentrations during pregnancy? A proof-of-concept pilot study. Trials 2020, 21, 380. [Google Scholar] [CrossRef]
- Bailey, S.W.; Ayling, J.E. The pharmacokinetic advantage of 5-methyltetrahydrofolate for minimization of the risk for birth defects. Sci. Rep. 2018, 8, 4096. [Google Scholar] [CrossRef]
- Lamers, Y.; Prinz-Langenohl, R.; Bramswig, S.; Pietrzik, K. Red blood cell folate concentrations increase more after supplementation with [6S]-5-methyltetrahydrofolate than with folic acid in women of childbearing age. Am. J. Clin. Nutr. 2006, 84, 156–161. [Google Scholar] [CrossRef]
- Clément, A.; Menezo, Y.; Cohen, M.; Cornet, D.; Clément, P. 5-Methyltetrahydrofolate reduces blood homocysteine level significantly in C677T methyltetrahydrofolate reductase single-nucleotide polymorphism carriers consulting for infertility. J. Gynecol. Obstet. Hum. Reprod. 2020, 49, 101622. [Google Scholar] [CrossRef]
- Servy, E.; Menezo, Y. The Methylene Tetrahydrofolate Reductase (MTHFR)isoform challenge. High doses of folic acid are not a suitable option compared to 5 Methyltetrahydrofolate treatment. Clin. Obstet. Gynecol. Reprod. Med. 2017, 3, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Ferrazzi, E.; Tiso, G.; Di Martino, D. Folic acid versus 5- methyl tetrahydrofolate supplementation in pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 253, 312–319. [Google Scholar] [CrossRef]
- Blake, G.; Zhao, X.; Yung, H.W.; Burton, G.J.; Ferguson-Smith, A.C. Defective folate metabolism causes germline epigenetic instability and distinguishes Hira as a phenotype inheritance biomarker. Nat. Commun. 2021, 12, 3714. [Google Scholar] [CrossRef] [PubMed]
- Plumptre, L.; Masih, S.P.; Ly, A.; Aufreiter, S.; Sohn, K.J. High concentrations of folate and unmetabolized folic acid in a cohort of pregnant Canadian women and umbilical cord blood. Am. J. Clin. Nutr. 2015, 102, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, C.M.; Sternberg, M.R.; Fazili, Z.; Yetley, E.A.; Lacher, D.A.; Bailey, R.L.; Johnson, C.L. Unmetabolized folic acid is detected in nearly all serum samples from US children, adolescents, and adults. J. Nutr. 2015, 145, 520–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghavan, R.; Selhub, J.; Paul, L.; Ji, Y.; Wang, G.; Hong, X.; Zuckerman, B.; Fallin, M.D.; Wang, X. A prospective birth cohort study on cord blood folate subtypes and risk of autism spectrum disorder. Am. J. Clin. Nutr. 2020, 112, 1304–1317. [Google Scholar] [CrossRef]
- Cosín-Tomás, M.; Luan, Y.; Leclerc, D.; Malysheva, O.V.; Lauzon, N. Moderate Folic Acid Supplementation in Pregnant Mice results in Behavioral Alterations in Offspring with Sex-Specific Changes in Methyl Metabolism. Nutrients 2020, 12, 1716. [Google Scholar] [CrossRef]
- Gong, M.; Dong, W.; He, T.; Shi, Z.; Huang, G. MTHFR 677C>T polymorphism increases the male infertility risk: A meta-analysis involving 26 studies. PLoS ONE 2015, 10, e0121147. [Google Scholar] [CrossRef]
- El Aarabi, M.; Christensen, K.E.; Chan, D.; Leclerc, D.; Landry, M.; Ly, L.; Rozen, R.; Trasler, J. Testicular MTHFR deficiency may explain sperm DNA hypomethylation associated with high dose folic acid supplementation. Hum. Mol. Genet. 2018, 27, 1123–1135. [Google Scholar] [CrossRef] [Green Version]
- Mtiraoui, N.; Zammiti, W.; Ghazouani, L.; Braham, N.J.; Saidi, S.; Finan, R.R.; Almawi, W.Y.; Mahjoub, T. Methylenetetrahydrofolate reductase C677T and A1298C polymorphism and changes in homocysteine concentrations in women with idiopathic recurrent pregnancy losses. Reproduction 2006, 131, 395–401. [Google Scholar] [CrossRef] [Green Version]
- Tara, S.S.; Ghaemimanesh, F.; Zarei, S.; Reihani-Sabet, F.; Pahlevanzadeh, Z.; Modarresi, M.H.; Jeddi-Tehrani, M. Methylenetetrahydrofolate reductase C677T and A1298C polymorphisms in male partners of recurrent miscarriages couples. J. Reprod. Infertil. 2015, 16, 193–198. [Google Scholar]
- Enciso, M.; Sarasa, J.; Xanthopoulou, L.; Bristow, S.; Bowles, M.F.; Fragouli, E.; Delhanty, J.; Wells, D. Polymorphisms in the MTHFR gene influence embryo viability and the incidence of aneuploidy. Hum. Genet. 2016, 135, 555–568. [Google Scholar] [CrossRef]
- Servy, E.J.; Jacquesson-Fournols, L.; Cohen, M.; Menezo, Y. MTHFR isoform carriers. 5-MTHF (5-methyl tetrahydrofolate) vs folic acid: A key to pregnancy outcome: A case series. J. Assist. Reprod. Genet. 2018, 35, 1431–1435. [Google Scholar] [CrossRef] [PubMed]
- Denomme, M.M.; McCallie, B.R.; Parks, J.C.; Schoolcraft, W.B.; Katz-Jaffe, M.G. Alterations in the sperm histone-retained epigenome are associated with unexplained male factor infertility and poor blastocyst development in donor oocyte IVF cycles. Hum. Reprod. 2017, 32, 2443–2450. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Jia, C.; Shi, Q.; Zhu, Y.; Liu, Y. Hyperhomocysteinemia in men with a reproductive history of fetal neural tube defects: Three case reports and literature review. Medicine 2019, 98, e13998. [Google Scholar] [CrossRef] [PubMed]
- Jacquesson-Fournols, L.; Alvarez, S.; Cohen, M.; Clement, P.; Menezo, Y. A paternal effect of MTHFR SNPs on gametes and embryos should not be overlooked: Case reports. J. Assist. Reprod. Genet. 2019, 36, 1351–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choux, C.; Binquet, C.; Carmignac, V.; Bruno, C.; Chapusot, C.; Barberet, J.; Lamotte, M.; Sagot, P.; Bourc’his, D.; Fauque, P. The epigenetic control of transposable elements and imprinted genes in newborns is a_affected by the mode of conception: ART versus spontaneous conception without underlying infertility. Hum. Reprod. 2018, 33, 331–340. [Google Scholar] [CrossRef]
- Menezo, Y.; Servy, E. Advanced Paternal Age and Endocrine Disruptors: Two Causes of Psychiatric Disorders in Children, with DNA Methylation Dys-Regulation as a Common Biochemical Mechanism, 2017 in Psychiatric Disorders. pp. 1–12. Available online: www.smgebooks.com (accessed on 19 December 2021).
- Menezo, Y.J.; Elder, K.; Dale, B. Link between Increased Prevalence of Autism Spectrum Disorder Syndromes and Oxidative Stress, DNA Methylation, and Imprinting: The Impact of the Environment. JAMA Pediatr. 2015, 169, 1066–1067. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, A.; Sportelli, V.; Ziller, M.; Spengler, D. Epigenomics of Major Depressive Disorders and Schizophrenia: Early Life Decides. Int. J. Mol. Sci. 2017, 18, 1711. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, E.H. Methylfolate as adjunctive treatment in major depression. Am. J. Psychiatry 2013, 170, 560. [Google Scholar] [CrossRef]
- Goyco Ortiz, L.E.; Servy, E.J.; Menezo, Y. A successful treatment with 5 methyltetrahydrofolate of a 677 TT MTHFR woman suffering premature ovarian insufficiency post a NHL (non-Hodgkin’s lymphoma) and RPL (repeat pregnancy losses). J. Assist. Reprod. Genet. 2019, 36, 65–67. [Google Scholar] [CrossRef]
- Navarrete Muñoz, E.M.; Valera-Gran, D.; Garcia-de-la-Hera, M.; Gonzalez-Palacios, S. High doses of folic acid in the periconceptional period and risk of low weight for gestational age at birth in a population-based cohort study INMA Project. Eur. J. Nutr. 2019, 58, 241–251. [Google Scholar] [CrossRef]
- Valera-Grande, D.; Navarrete-Muñoz, E.M.; Garcia de la Hera, M.; Fernández-Somoano, A.; Tardón, A. INMA Project Effect of maternal high dosages of folic acid supplements on neurocognitive development in children at 4-5 y of age: The prospective birth cohort Infancia y Medio Ambiente (INMA) study. Am. J. Clin. Nutr. 2017, 106, 878–887. [Google Scholar]
- Li, Y.; Huang, T.; Zheng, Y.; Muka, T.; Troup, J.; Hu, F.B. Folic Acid Supplementation and the Risk of Cardiovascular Diseases: A Meta-Analysis of Randomized Controlled Trials. J. Am. Heart. Assoc. 2016, 5, e003768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, A.K.; Dhalla, N.S. Effectiveness of Some Vitamins in the Prevention of Cardiovascular Disease: A Narrative Review. Front. Physiol. 2021, 12, 729255. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, E.H. Benefits and risks of folic acid to the nervous system. J. Neurol. Neurosurg. Psychiatry 2002, 72, 567–571. [Google Scholar] [CrossRef] [Green Version]
- Martí-Carvajal, A.J.; Solà, I.; Lathyris, D.; Dayer, M. Homocysteine-lowering interventions for preventing cardiovascular events. Cochrane Database Syst. Rev. 2017, 8, CD006612. [Google Scholar]
- Yang, H.T.; Lee, M.; Hong, K.S.; Ovbiagele, B.; Saver, J.L. Efficacy of folic acid supplementation in cardiovascular disease prevention: An updated meta-analysis of randomized controlled trials. Eur. J. Intern. Med. 2012, 23, 745–754. [Google Scholar] [CrossRef]
- Jeeja, M.C.; Jayakrishnan, T.; Narayanan, P.V.; Kumar, M.S.; Thejus, T. Folic acid supplementation on homocysteine levels in children taking antiepileptic drugs: A randomized controlled trial. J. Pharmacol. Pharmacother. 2014, 5, 93–99. [Google Scholar]
- Auger, N.; Bilodeau-Bertrand, M.; Tith, R.M.; Arbour, L. Bariatric surgery and the risk of congenital anomalies in subsequent pregnancies. Am. J. Clin. Nutr. 2019, 110, 1168–1174. [Google Scholar] [CrossRef]
- Morgan, S.L.; Oster, R.A.; Lee, J.Y.; Alarcón, G.S.; Baggott, J.E. The effect of folic acid and folinic acid supplements on purine metabolism in methotrexate-treated rheumatoid arthritis. Arthritis. Rheum. 2004, 50, 3104–3111. [Google Scholar] [CrossRef]
- Baylin, S.B.; Jones, P.A. Epigenetic Determinants of Cancer. Cold Spring Harb. Perspect. Biol. 2016, 8, a019505. [Google Scholar] [CrossRef] [Green Version]
- Koenig, K.L.; Scarmo, S.; Afanasyeva, Y.; Clendenen, T.V.; Ueland, P.M.; Zeleniuch-Jacquotte, A. Circulating unmetabolized folic acid and 5-methyltetrahydrofolate and risk of breast cancer: A nested case-control study. Eur. J. Clin. Nutr. 2020, 74, 1306–1315. [Google Scholar] [CrossRef] [PubMed]
- Moazzen, S.; Dolatkhah, R.; Tabrizi, J.S.; Shaarbafi, J.; Alizadeh, B.Z. Folic acid intake and folate status and colorectal cancer risk: A systematic review and meta-analysis. Clin. Nutr. 2018, 37, 1926–1934. [Google Scholar] [CrossRef]
- Oliai- Araghi, S.; Kiefte-de Jong, J.C.; van Dijk, S.C.; Swart, K.M.A.; van Laarhoven, H.W.; van Schoor, N.M.; de Groot, L.C.P.G.M.; Lemmens, V.; Stricker, B.H.; Uitterlinden, A.G.; et al. Folic Acid and Vitamin B12 Supplementation and the Risk of Cancer: Long-term Follow-up of the B Vitamins for the Prevention of Osteoporotic Fractures (B-PROOF) Trial. Cancer Epidemiol. Biomarkers Prev. 2019, 28, 275–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rycyna, K.J.; Bacich, D.J.; O’Keefe, D.S. Opposing roles of folate in prostate cancer. Urology 2013, 82, 1197–1203. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, J.C. Folic acid and risk of prostate cancer: Results from a randomized clinical trial. J. Natl. Cancer Inst. 2009, 101, 432–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirsch, S.; Sanchez, H.; Albala, C.; de la Maza, M.P.; Barrera, G.; Leiva, L.; Bunout, D. Colon cancer in Chile before and after the start of the flour fortification program with folic acid. Eur. J. Gastroenterol. Hepatol. 2009, 21, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Hornstra, J.; Rocha, M.; Jansen, G.; Assaraf, Y. Folic Acid Impairs the Uptake of 5-Methyltetrahydrofolate in Human Umbilical Vascular Endothelial Cells. J. Cardiovasc. Pharmacol. 2017, 70, 271–275. [Google Scholar] [CrossRef] [Green Version]
- Padmanabhan, N.; Menelaou, K.; Gao, J.; Anderson, A.; Blake, G.; Daw, B.N.; Watson, E.D. Abnormal folate metabolism causes age-, sex- and parent-of-origin-specific haematological defects in mice. J. Physiol. 2018, 596, 4341–4360. [Google Scholar] [CrossRef]
- Ly, L.; Chan, D.; Trasler, J.M. Developmental windows of susceptibility for epigenetic inheritance through the male germline. Semin Cell Dev. Biol. 2015, 43, 96–105. [Google Scholar] [CrossRef]
- Pickell, L.; Brown, K.; Li, D.; Wang, X.L.; Deng, L.; Wu, Q.; Selhub, J.; Luo, L.; Jerome-Majewska, L.; Rozen, R. High intake of folic acid disrupts embryonic development in mice. Birth Defects Res. A Clin. Mol. Teratol. 2011, 91, 8–19. [Google Scholar] [CrossRef]
- Manikkam, M.; Tracey, R.; Guerrero-Bosagna, C.; Skinner, M.K. Plastics derived endocrine disruptors (BPA, DEHP and DBP) induce epigenetic transgenerational inheritance of obesity, reproductive disease and sperm epimutations. PLoS ONE 2013, 8, e55387. [Google Scholar] [CrossRef] [PubMed]
- Ben Maamar, M.; Nilsson, E.E.; Skinner, M.K. Epigenetic transgenerational inheritance, gametogenesis and germline development. Biol. Reprod. 2021, 105, 570–592. [Google Scholar] [CrossRef] [PubMed]
- Menezo, Y.; Dale, B.; Elder, K. The negative impact of the environment on methylation/epigenetic marking in gametes and embryos: A plea for action to protect the fertility of future generations. Mol. Reprod. Dev. 2019, 86, 1273–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menezo, Y.; Elder, K.; Clement, A.; Clement, P. Folic Acid, Folinic Acid, 5 Methyl TetraHydroFolate Supplementation for Mutations That Affect Epigenesis through the Folate and One-Carbon Cycles. Biomolecules 2022, 12, 197. https://doi.org/10.3390/biom12020197
Menezo Y, Elder K, Clement A, Clement P. Folic Acid, Folinic Acid, 5 Methyl TetraHydroFolate Supplementation for Mutations That Affect Epigenesis through the Folate and One-Carbon Cycles. Biomolecules. 2022; 12(2):197. https://doi.org/10.3390/biom12020197
Chicago/Turabian StyleMenezo, Yves, Kay Elder, Arthur Clement, and Patrice Clement. 2022. "Folic Acid, Folinic Acid, 5 Methyl TetraHydroFolate Supplementation for Mutations That Affect Epigenesis through the Folate and One-Carbon Cycles" Biomolecules 12, no. 2: 197. https://doi.org/10.3390/biom12020197
APA StyleMenezo, Y., Elder, K., Clement, A., & Clement, P. (2022). Folic Acid, Folinic Acid, 5 Methyl TetraHydroFolate Supplementation for Mutations That Affect Epigenesis through the Folate and One-Carbon Cycles. Biomolecules, 12(2), 197. https://doi.org/10.3390/biom12020197