
Ibuprofen in Therapeutic Concentrations Affects the Secretion of Human Bone Marrow Mesenchymal Stromal Cells, but Not Their Proliferative and Migratory Capacity

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Isolation and Culture
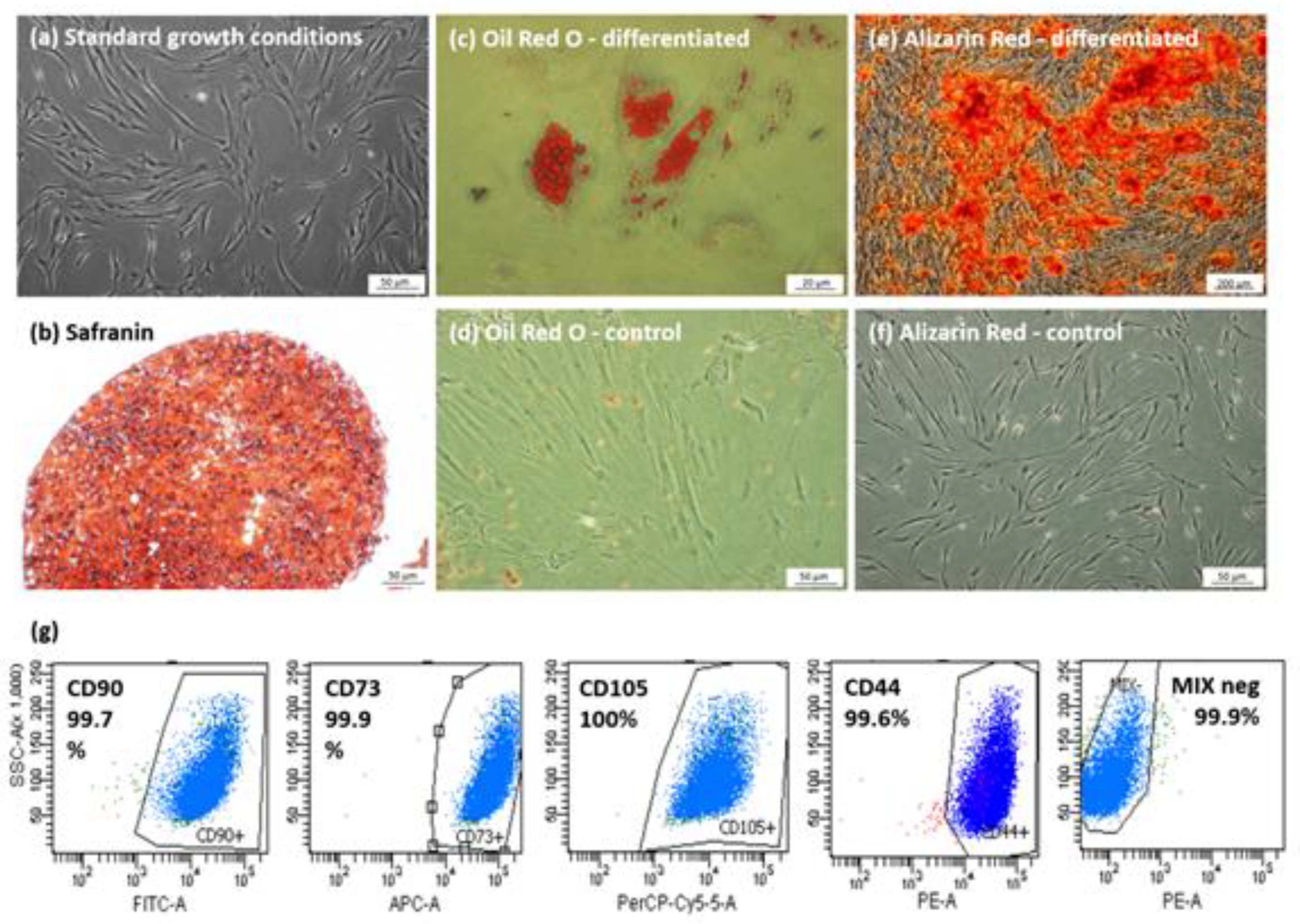
2.2. hBM-MSCs Identification
2.2.1. Antigen Characterisation
2.2.2. Multilinear Differentiation
2.3. Ibuprofen Treatment
2.3.1. MTT Assay
2.3.2. PGE2 ELISA Immunoassay
2.4. BrdU Proliferation Assay
2.5. Transwell Migration Assay
2.6. Cell Migration Scratch Assay—Live Cell Imaging
2.7. Cytokine Secretion Analysis
2.7.1. Proteome Profiler Antibody Assay
2.7.2. Luminex Multiplex Immunoassay
2.7.3. BCA Protein Assay
2.8. Statistical Analysis
3. Results
3.1. Characterization of Isolated hBM-MSCs Populations
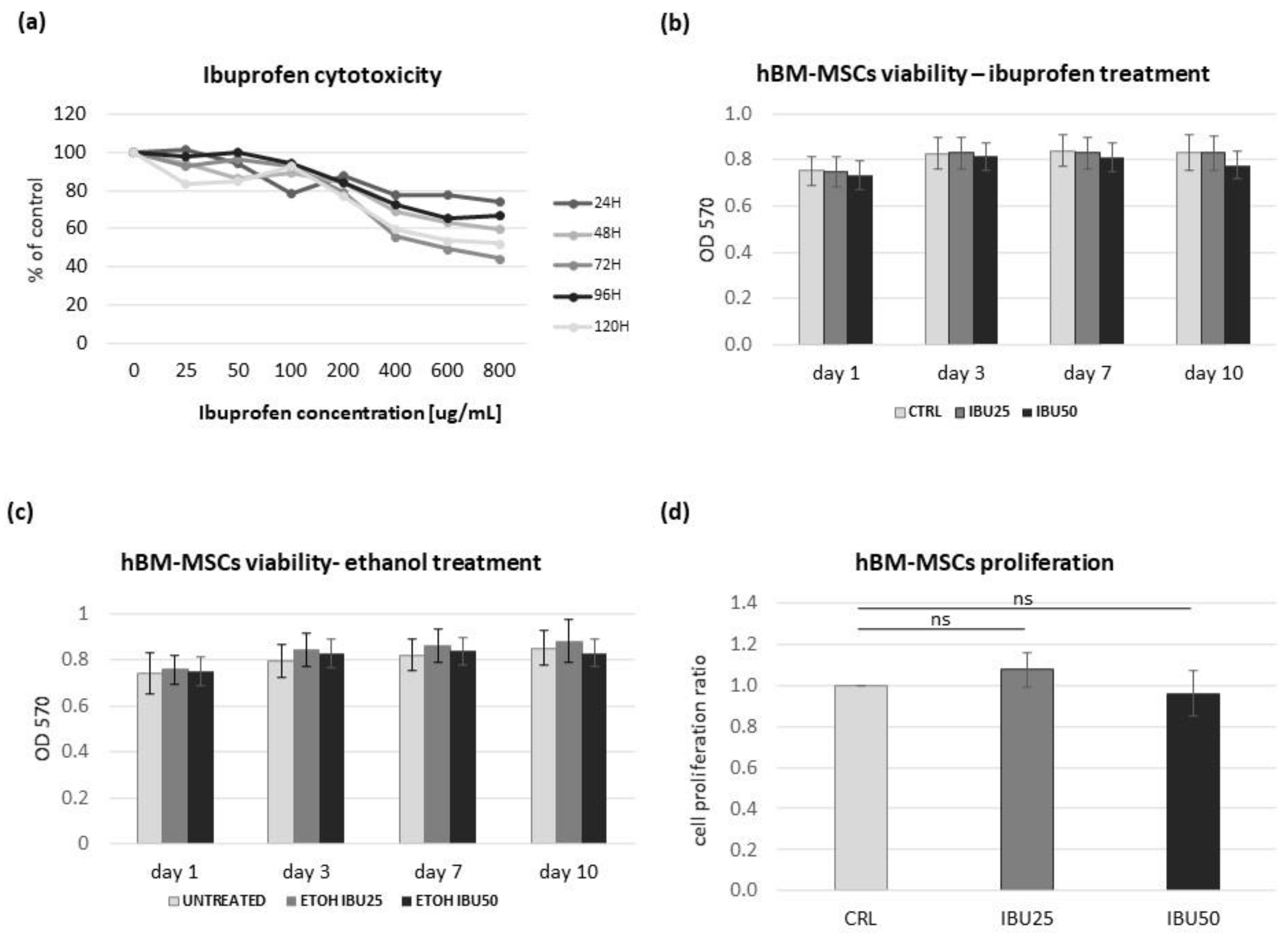
3.2. Ibuprofen Reduces the Viability of hBM-MSCs in a Dose-Dependent but Not a Time-Dependent Manner
3.3. Ibuprofen Does Not Affect the hBM-MSCs Proliferation
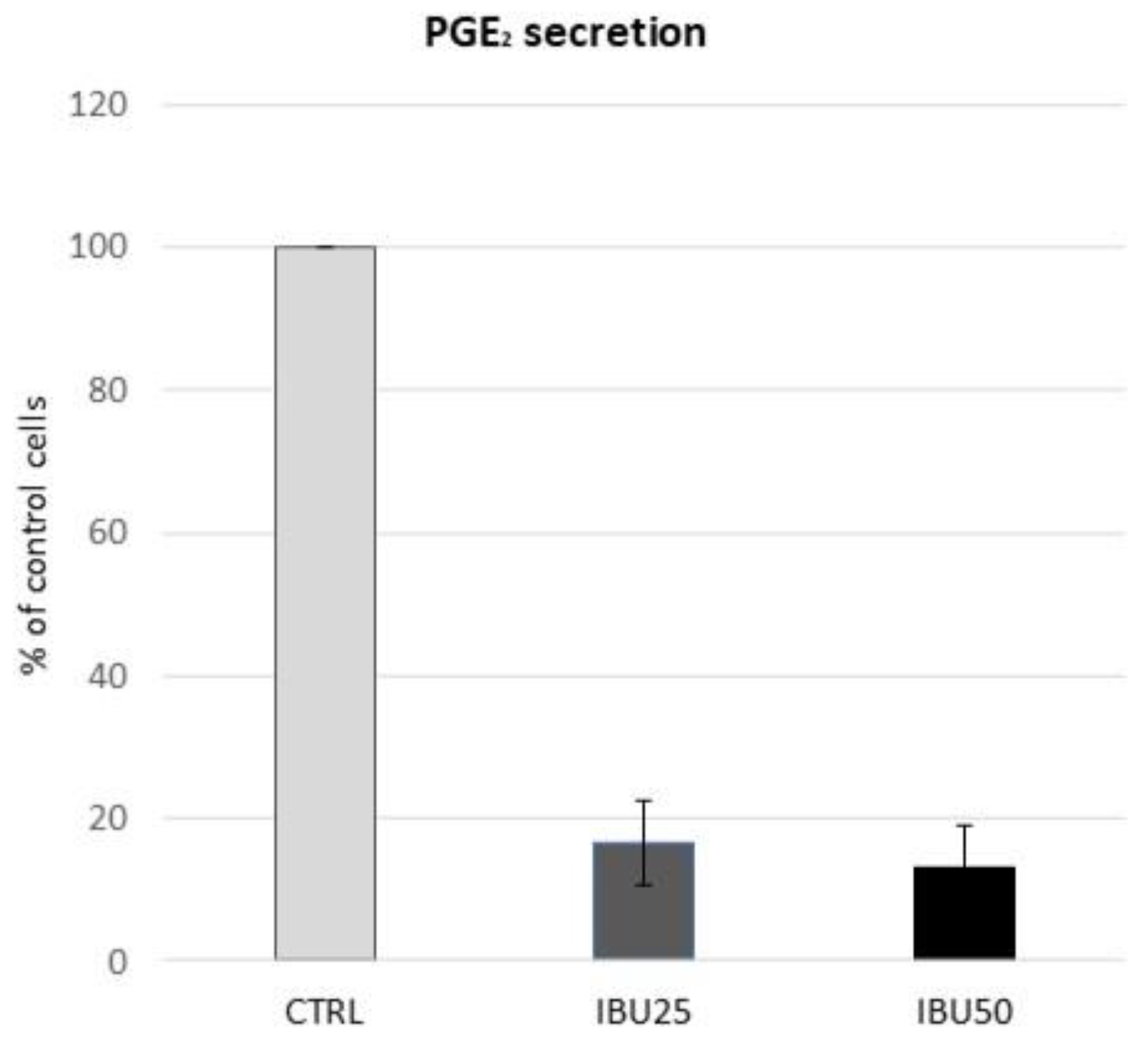
3.4. Ibuprofen in Therapeutic Doses Substantially Reduce the Secretion of PGE2 by hBM-MSCs
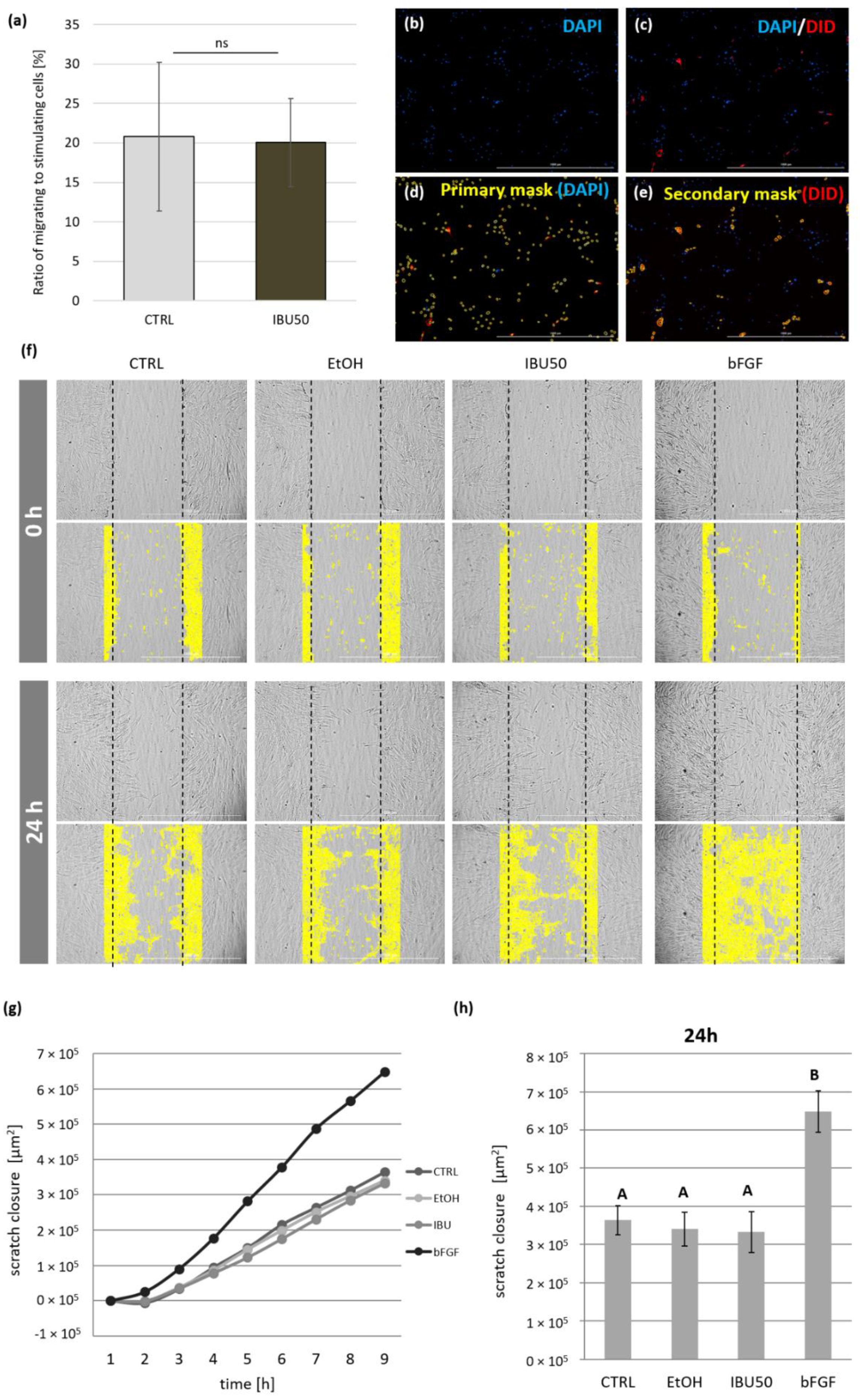
3.5. Ibuprofen Does Not Affect the hBM-MSCs Migration
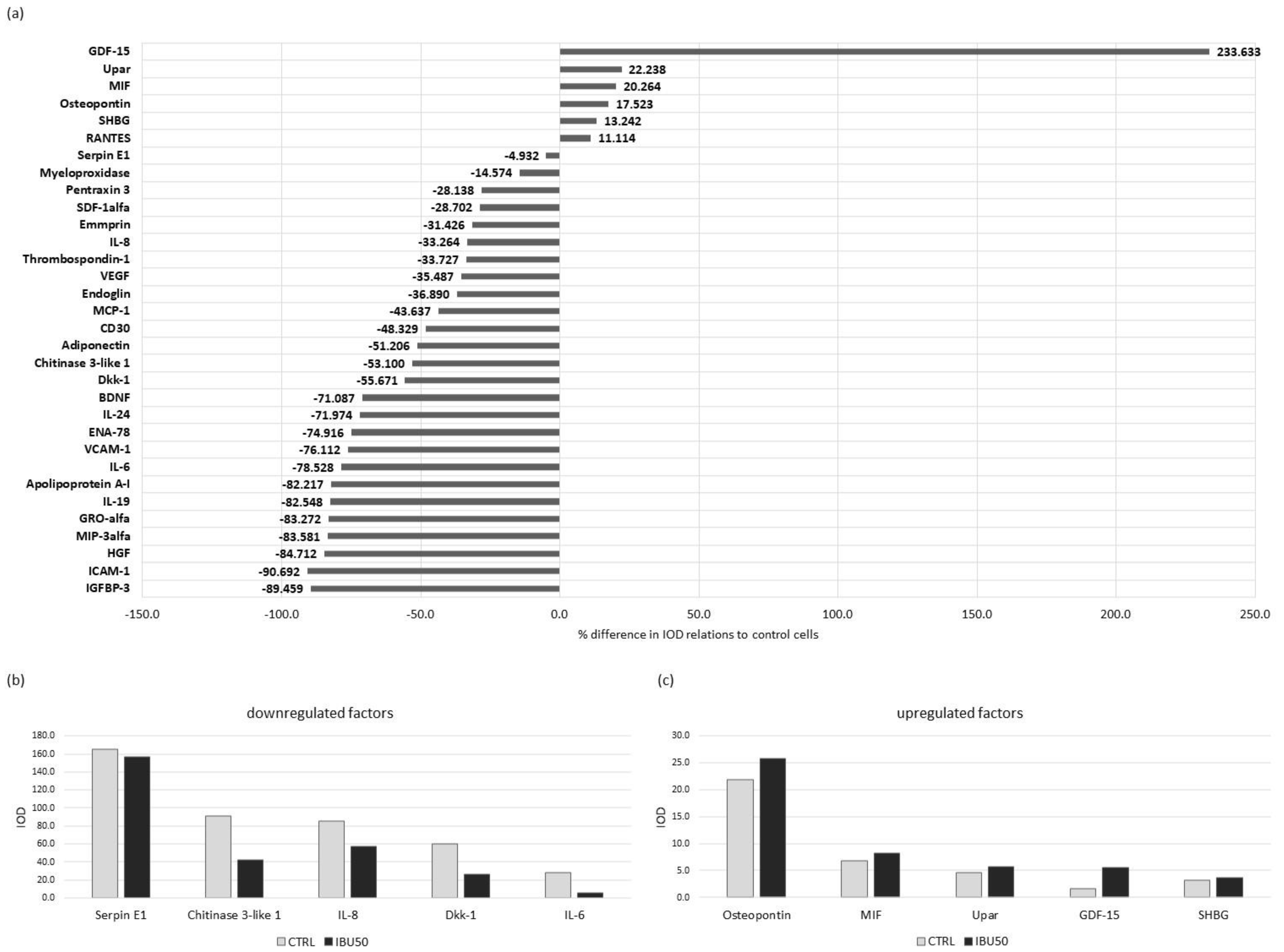
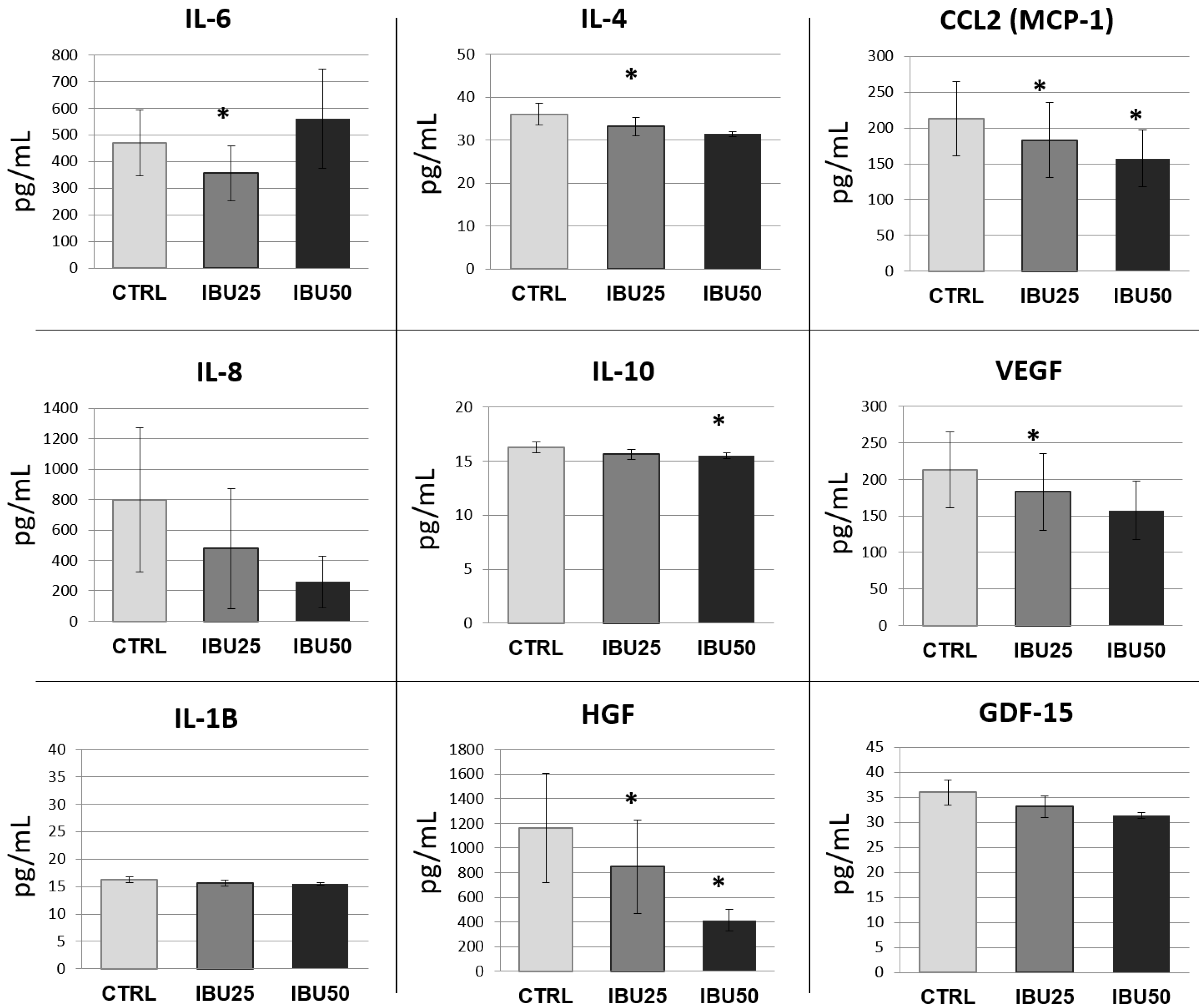
3.6. Ibuprofen Affects hBM-MSCs Secretion Profile
3.6.1. Ibuprofen Changes Protein Secretion in BM-MSC
3.6.2. Ibuprofen Significantly Reduces the Secretion of Selected Cytokines
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boilly, B.; Vercoutter-Edouart, A.S.; Hondermarck, H.; Nurcombe, V.; Le Bourhis, X. FGF signals for cell proliferation and migration through different pathways. Cytokine Growth Factor Rev. 2000, 11, 295–302. [Google Scholar] [CrossRef]
- Rennert, R.C.; Sorkin, M.; Garg, R.K.; Gurtner, G.C. Stem cell recruitment after injury: Lessons for regenerative medicine. Regen. Med. 2012, 7, 833–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pojda, Z.; Machaj, E.; Kurzyk, A.; Mazur, S.; Debski, T.; Gilewicz, J.; Wysocki, J. Mesenchymal stem cells. Postepy Biochem. 2013, 59, 187–197. [Google Scholar] [PubMed]
- Matheakakis, A.; Batsali, A.; Papadaki, H.A.; Pontikoglou, C.G. Therapeutic Implications of Mesenchymal Stromal Cells and Their Extracellular Vesicles in Autoimmune Diseases: From Biology to Clinical Applications. Int. J. Mol. Sci. 2021, 22, 132. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chen, X.; Wang, W.E.; Zeng, C. How to Improve the Survival of Transplanted Mesenchymal Stem Cell in Ischemic Heart? Stem Cells Int. 2016, 2016, 9682757. [Google Scholar] [CrossRef] [Green Version]
- Prockop, D.J. Repair of tissues by adult stem/progenitor cells (MSCs): Controversies, myths, and changing paradigms. Mol. Ther. 2009, 17, 939–946. [Google Scholar] [CrossRef]
- Lee, S.; Choi, E.; Cha, M.J.; Hwang, K.C. Cell adhesion and long-term survival of transplanted mesenchymal stem cells: A prerequisite for cell therapy. Oxid. Med. Cell. Longev. 2015, 2015, 632902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phinney, D.G.; Prockop, D.J. Concise review: Mesenchymal stem/multipotent stromal cells: The state of transdifferentiation and modes of tissue repair–current views. Stem Cells 2007, 25, 2896–2902. [Google Scholar] [CrossRef]
- Andreeva, E.; Bobyleva, P.; Gornostaeva, A.; Buravkova, L. Interaction of multipotent mesenchymal stromal and immune cells: Bidirectional effects. Cytotherapy 2017, 19, 1152–1166. [Google Scholar] [CrossRef]
- Nauta, A.J.; Fibbe, W.E. Immunomodulatory properties of mesenchymal stromal cells. Blood 2007, 110, 3499–3506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Girolamo, L.; Lucarelli, E.; Alessandri, G.; Avanzini, M.A.; Bernardo, M.E.; Biagi, E.; Brini, A.T.; D’Amico, G.; Fagioli, F.; Ferrero, I.; et al. Mesenchymal stem/stromal cells: A new ‘‘cells as drugs’’ paradigm. Efficacy and critical aspects in cell therapy. Curr. Pharm. Des. 2013, 19, 2459–2473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Blanc, K.; Mougiakakos, D. Multipotent mesenchymal stromal cells and the innate immune system. Nat. Rev. Immunol. 2012, 12, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Meirelles Lda, S.; Fontes, A.M.; Covas, D.T.; Caplan, A.I. Mechanisms involved in the therapeutic properties of mesenchymal stem cells. Cytokine Growth Factor Rev. 2009, 20, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Szala, S.; Wisniewska, E.; Czapla, J. Mesenchymal stromal cells. Postepy Hig. Med. Dosw. 2014, 68, 1287–1298. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Pillinger, M.H.; Abramson, S.B. Prostaglandin E2 synthesis and secretion: The role of PGE2 synthases. Clin. Immunol. 2006, 119, 229–240. [Google Scholar] [CrossRef]
- English, K. Mechanisms of mesenchymal stromal cell immunomodulation. Immunol. Cell Biol. 2013, 91, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, S.; Pittenger, M.F. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood 2005, 105, 1815–1822. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.X.; Zhang, Y.; Liu, B.; Zhang, S.X.; Wu, Y.; Yu, X.D.; Mao, N. Human mesenchymal stem cells inhibit differentiation and function of monocyte-derived dendritic cells. Blood 2005, 105, 4120–4126. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, K.; Leelahavanichkul, A.; Yuen, P.S.; Mayer, B.; Parmelee, A.; Doi, K.; Robey, P.G.; Leelahavanichkul, K.; Koller, B.H.; Brown, J.M.; et al. Bone marrow stromal cells attenuate sepsis via prostaglandin E(2)-dependent reprogramming of host macrophages to increase their interleukin-10 production. Nat. Med. 2009, 15, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.C.; Kim, H.S.; Shin, T.H.; Kang, I.; Lee, J.Y.; Kim, J.J.; Kang, H.K.; Seo, Y.; Lee, S.; Yu, K.R.; et al. PGE2 maintains self-renewal of human adult stem cells via EP2-mediated autocrine signaling and its production is regulated by cell-to-cell contact. Sci. Rep. 2016, 6, 26298. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Han, J.; Xu, X.; Xu, J.; Liu, L.; Huang, Y.; Yang, Y.; Qiu, H. PGE2 Promotes the Migration of Mesenchymal Stem Cells through the Activation of FAK and ERK1/2 Pathway. Stem Cells Int. 2017, 2017, 8178643. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Lu, X.; Zou, L.; Xu, X.; Qiu, H. E-Prostanoid 2 Receptor Overexpression Promotes Mesenchymal Stem Cell Attenuated Lung Injury. Hum. Gene Ther. 2016, 27, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Kota, D.J.; Prabhakara, K.S.; Toledano-Furman, N.; Bhattarai, D.; Chen, Q.; DiCarlo, B.; Smith, P.; Triolo, F.; Wenzel, P.L.; Cox, C.S., Jr.; et al. Prostaglandin E2 Indicates Therapeutic Efficacy of Mesenchymal Stem Cells in Experimental Traumatic Brain Injury. Stem Cells 2017, 35, 1416–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varrassi, G.; Pergolizzi, J.V.; Dowling, P.; Paladini, A. Ibuprofen Safety at the Golden Anniversary: Are all NSAIDs the Same? A Narrative Review. Adv. Ther. 2020, 37, 61–82. [Google Scholar] [CrossRef] [Green Version]
- Sienko, D.; Klimczak-Tomaniak, D.; Kulesza, A.; Symonides, H.; Kuch, M.; Paczek, L.; Burdzinska, A. The influence of oxygen deprivation and donor age on the effect of statins on human mesenchymal stromal cells. Tissue Cell 2020, 67, 101427. [Google Scholar] [CrossRef]
- Chen, M.R.; Dragoo, J.L. The effect of nonsteroidal anti-inflammatory drugs on tissue healing. Knee Surg. Sports Traumatol. Arthrosc. 2013, 21, 540–549. [Google Scholar] [CrossRef]
- Kawaguchi, H.; Pilbeam, C.C.; Harrison, J.R.; Raisz, L.G. The role of prostaglandins in the regulation of bone metabolism. Clin. Orthop. Relat. Res. 1995, 36–46. [Google Scholar]
- Mackey, A.L.; Kjaer, M.; Dandanell, S.; Mikkelsen, K.H.; Holm, L.; Dossing, S.; Kadi, F.; Koskinen, S.O.; Jensen, C.H.; Schroder, H.D.; et al. The influence of anti-inflammatory medication on exercise-induced myogenic precursor cell responses in humans. J. Appl. Physiol. 2007, 103, 425–431. [Google Scholar] [CrossRef] [Green Version]
- Mikkelsen, U.R.; Langberg, H.; Helmark, I.C.; Skovgaard, D.; Andersen, L.L.; Kjaer, M.; Mackey, A.L. Local NSAID infusion inhibits satellite cell proliferation in human skeletal muscle after eccentric exercise. J. Appl. Physiol. 2009, 107, 1600–1611. [Google Scholar] [CrossRef] [Green Version]
- Mallick, E.; Scutt, N.; Scutt, A.; Rolf, C. Passage and concentration-dependent effects of Indomethacin on tendon derived cells. J. Orthop. Surg. Res. 2009, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Christensen, B.; Dandanell, S.; Kjaer, M.; Langberg, H. Effect of anti-inflammatory medication on the running-induced rise in patella tendon collagen synthesis in humans. J. Appl. Physiol. 2011, 110, 137–141. [Google Scholar] [CrossRef] [Green Version]
- Muller, M.; Raabe, O.; Addicks, K.; Wenisch, S.; Arnhold, S. Effects of non-steroidal anti-inflammatory drugs on proliferation, differentiation and migration in equine mesenchymal stem cells. Cell Biol. Int. 2011, 35, 235–248. [Google Scholar] [CrossRef]
- Styner, M.; Sen, B.; Xie, Z.; Case, N.; Rubin, J. Indomethacin promotes adipogenesis of mesenchymal stem cells through a cyclooxygenase independent mechanism. J. Cell Biochem. 2010, 111, 1042–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, N.; Kim, S.; Hosoya, K.; Okumura, M. Compensatory cellular reactions to nonsteroidal anti-inflammatory drugs on osteogenic differentiation in canine bone marrow-derived mesenchymal stem cells. J. Vet. Med. Sci. 2014, 76, 629–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pountos, I.; Giannoudis, P.V.; Jones, E.; English, A.; Churchman, S.; Field, S.; Ponchel, F.; Bird, H.; Emery, P.; McGonagle, D. NSAIDS inhibit in vitro MSC chondrogenesis but not osteogenesis: Implications for mechanism of bone formation inhibition in man. J. Cell. Mol. Med. 2011, 15, 525–534. [Google Scholar] [CrossRef] [Green Version]
- Burdzinska, A.; Dybowski, B.; Zarychta-Wisniewska, W.; Kulesza, A.; Zagozdzon, R.; Gajewski, Z.; Paczek, L. The Anatomy of Caprine Female Urethra and Characteristics of Muscle and Bone Marrow Derived Caprine Cells for Autologous Cell Therapy Testing. Anat. Rec. 2017, 300, 577–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonab, M.M.; Alimoghaddam, K.; Talebian, F.; Ghaffari, S.H.; Ghavamzadeh, A.; Nikbin, B. Aging of mesenchymal stem cell in vitro. BMC Cell Biol. 2006, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Zielniok, K.; Burdzinska, A.; Kaleta, B.; Zagozdzon, R.; Paczek, L. Vadadustat, a HIF Prolyl Hydroxylase Inhibitor, Improves Immunomodulatory Properties of Human Mesenchymal Stromal Cells. Cells 2020, 9, 2396. [Google Scholar] [CrossRef] [PubMed]
- Davies, N.M. Clinical pharmacokinetics of ibuprofen. The first 30 years. Clin. Pharmacokinet. 1998, 34, 101–154. [Google Scholar] [CrossRef]
- Dymowska, M.; Aksamit, A.; Zielniok, K.; Kniotek, M.; Kaleta, B.; Roszczyk, A.; Zych, M.; Dabrowski, F.; Paczek, L.; Burdzinska, A. Interaction between Macrophages and Human Mesenchymal Stromal Cells Derived from Bone Marrow and Wharton’s Jelly-A Comparative Study. Pharmaceutics 2021, 13, 1822. [Google Scholar] [CrossRef]
- Sotiropoulou, P.A.; Perez, S.A.; Salagianni, M.; Baxevanis, C.N.; Papamichail, M. Characterization of the optimal culture conditions for clinical scale production of human mesenchymal stem cells. Stem Cells 2006, 24, 462–471. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Song, N.; Scholtemeijer, M.; Shah, K. Mesenchymal Stem Cell Immunomodulation: Mechanisms and Therapeutic Potential. Trends Pharmacol. Sci. 2020, 41, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Perrott, D.A.; Piira, T.; Goodenough, B.; Champion, G.D. Efficacy and safety of acetaminophen vs ibuprofen for treating children’s pain or fever: A meta-analysis. Arch. Pediatr. Adolesc. Med. 2004, 158, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Potthast, H.; Dressman, J.B.; Junginger, H.E.; Midha, K.K.; Oeser, H.; Shah, V.P.; Vogelpoel, H.; Barends, D.M. Biowaiver monographs for immediate release solid oral dosage forms: Ibuprofen. J. Pharm. Sci. 2005, 94, 2121–2131. [Google Scholar] [CrossRef]
- Di Nicola, M.; Carlo-Stella, C.; Magni, M.; Milanesi, M.; Longoni, P.D.; Matteucci, P.; Grisanti, S.; Gianni, A.M. Human bone marrow stromal cells suppress T-lymphocyte proliferation induced by cellular or nonspecific mitogenic stimuli. Blood 2002, 99, 3838–3843. [Google Scholar] [CrossRef]
- Cottrell, J.; O’Connor, J.P. Effect of Non-Steroidal Anti-Inflammatory Drugs on Bone Healing. Pharmaceuticals 2010, 3, 1668–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, W.C.; Hsu, C.C.; Chen, C.P.; Chen, M.J.; Lin, M.S.; Pang, J.H. Ibuprofen inhibition of tendon cell migration and down-regulation of paxillin expression. J. Orthop. Res. 2006, 24, 551–558. [Google Scholar] [CrossRef]
- Brooks, G.; Yu, X.M.; Wang, Y.; Crabbe, M.J.; Shattock, M.J.; Harper, J.V. Non-steroidal anti-inflammatory drugs (NSAIDs) inhibit vascular smooth muscle cell proliferation via differential effects on the cell cycle. J. Pharm. Pharmacol. 2003, 55, 519–526. [Google Scholar] [CrossRef]
- Duffy, M.M.; Pindjakova, J.; Hanley, S.A.; McCarthy, C.; Weidhofer, G.A.; Sweeney, E.M.; English, K.; Shaw, G.; Murphy, J.M.; Barry, F.P.; et al. Mesenchymal stem cell inhibition of T-helper 17 cell- differentiation is triggered by cell-cell contact and mediated by prostaglandin E2 via the EP4 receptor. Eur. J. Immunol. 2011, 41, 2840–2851. [Google Scholar] [CrossRef]
- Jang, M.W.; Yun, S.P.; Park, J.H.; Ryu, J.M.; Lee, J.H.; Han, H.J. Cooperation of Epac1/Rap1/Akt and PKA in prostaglandin E(2) -induced proliferation of human umbilical cord blood derived mesenchymal stem cells: Involvement of c-Myc and VEGF expression. J. Cell. Physiol. 2012, 227, 3756–3767. [Google Scholar] [CrossRef] [PubMed]
- Kleiveland, C.R.; Kassem, M.; Lea, T. Human mesenchymal stem cell proliferation is regulated by PGE2 through differential activation of cAMP-dependent protein kinase isoforms. Exp. Cell Res. 2008, 314, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Zarychta-Wisniewska, W.; Burdzinska, A.; Zielniok, K.; Koblowska, M.; Gala, K.; Pedzisz, P.; Iwanicka-Nowicka, R.; Fogtman, A.; Aksamit, A.; Kulesza, A.; et al. The Influence of Cell Source and Donor Age on the Tenogenic Potential and Chemokine Secretion of Human Mesenchymal Stromal Cells. Stem Cells Int. 2019, 2019, 1613701. [Google Scholar] [CrossRef] [PubMed]
- Scutera, S.; Salvi, V.; Lorenzi, L.; Piersigilli, G.; Lonardi, S.; Alotto, D.; Casarin, S.; Castagnoli, C.; Dander, E.; D’Amico, G.; et al. Adaptive Regulation of Osteopontin Production by Dendritic Cells Through the Bidirectional Interaction With Mesenchymal Stromal Cells. Front. Immunol. 2018, 9, 1207. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T. IL-6 in inflammation, autoimmunity and cancer. Int. Immunol. 2021, 33, 127–148. [Google Scholar] [CrossRef]
- Lee, S.; Kim, O.J.; Lee, K.O.; Jung, H.; Oh, S.H.; Kim, N.K. Enhancing the Therapeutic Potential of CCL2-Overexpressing Mesenchymal Stem Cells in Acute Stroke. Int. J. Mol. Sci. 2020, 21, 7795. [Google Scholar] [CrossRef]
- Nakamura, T.; Sakai, K.; Nakamura, T.; Matsumoto, K. Hepatocyte growth factor twenty years on: Much more than a growth factor. J. Gastroenterol. Hepatol. 2011, 26 (Suppl. 1), 188–202. [Google Scholar] [CrossRef] [Green Version]
- Gallelli, L.; Galasso, O.; Falcone, D.; Southworth, S.; Greco, M.; Ventura, V.; Romualdi, P.; Corigliano, A.; Terracciano, R.; Savino, R.; et al. The effects of nonsteroidal anti-inflammatory drugs on clinical outcomes, synovial fluid cytokine concentration and signal transduction pathways in knee osteoarthritis. A randomized open label trial. Osteoarthr. Cartil. 2013, 21, 1400–1408. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Dependent Correlations | |
|---|---|
| Ibuprofen Concentration [ug/mL] | Pearson’s Correlation Coefficient (r) |
| 25 | 0.46 |
| 50 | 0.5 |
| 100 | 0.24 |
| 200 | 0.72 |
| 400 | 0.65 |
| 600 | 0.7 |
| 800 | 0.53 |
| average | 0.54 |
| Dose dependent correlations | |
| treatment time [h] | Pearson’s correlation coefficient (r) |
| 24 | 0.9 |
| 48 | 0.96 |
| 72 | 0.95 |
| 96 | 0.94 |
| 120 | 0.89 |
| average | 0.92 |
| PGE2 Concentration [pg/mL] | |||
|---|---|---|---|
| CTRL | IBU25 | IBU50 | |
| Population 1 | 12.59 | underdetection | underdetection |
| Population 2 | 525.54 | 12.48 | 11.62 |
| Population 3 | 68.21 | 8.34 | 9.92 |
| Population 4 | 38.55 | 8.84 | 8.58 |
| Population 5 | 32.14 | 9.15 | underdetection |
| Population 6 | 28.78 | underdetection | underdetection |
| mean | 117.63 | 9.71 | 10.04 |
| SD | 200.66 | 1.88 | 1.52 |
| SEM | 81.92 | 0.77 | 0.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulesza, A.; Zielniok, K.; Hawryluk, J.; Paczek, L.; Burdzinska, A. Ibuprofen in Therapeutic Concentrations Affects the Secretion of Human Bone Marrow Mesenchymal Stromal Cells, but Not Their Proliferative and Migratory Capacity. Biomolecules 2022, 12, 287. https://doi.org/10.3390/biom12020287
Kulesza A, Zielniok K, Hawryluk J, Paczek L, Burdzinska A. Ibuprofen in Therapeutic Concentrations Affects the Secretion of Human Bone Marrow Mesenchymal Stromal Cells, but Not Their Proliferative and Migratory Capacity. Biomolecules. 2022; 12(2):287. https://doi.org/10.3390/biom12020287
Chicago/Turabian StyleKulesza, Agnieszka, Katarzyna Zielniok, Jakub Hawryluk, Leszek Paczek, and Anna Burdzinska. 2022. "Ibuprofen in Therapeutic Concentrations Affects the Secretion of Human Bone Marrow Mesenchymal Stromal Cells, but Not Their Proliferative and Migratory Capacity" Biomolecules 12, no. 2: 287. https://doi.org/10.3390/biom12020287
APA StyleKulesza, A., Zielniok, K., Hawryluk, J., Paczek, L., & Burdzinska, A. (2022). Ibuprofen in Therapeutic Concentrations Affects the Secretion of Human Bone Marrow Mesenchymal Stromal Cells, but Not Their Proliferative and Migratory Capacity. Biomolecules, 12(2), 287. https://doi.org/10.3390/biom12020287