
HIV-1 Trans Infection via TNTs Is Impeded by Targeting C5aR



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Generation of Human Monocyte-Derived DCs and CD4+ T Cells
2.3. Multicolor Fluorescence-Activated Cell Sorting (FACS) Analyses
2.4. Plasmids
2.5. Virus Production
2.6. Virus Opsonization
2.7. p24 ELISA
2.8. Virus Capture Assay
2.9. Infection of Cells
2.10. Confocal Analysis
2.11. Statistical Analysis
3. Results
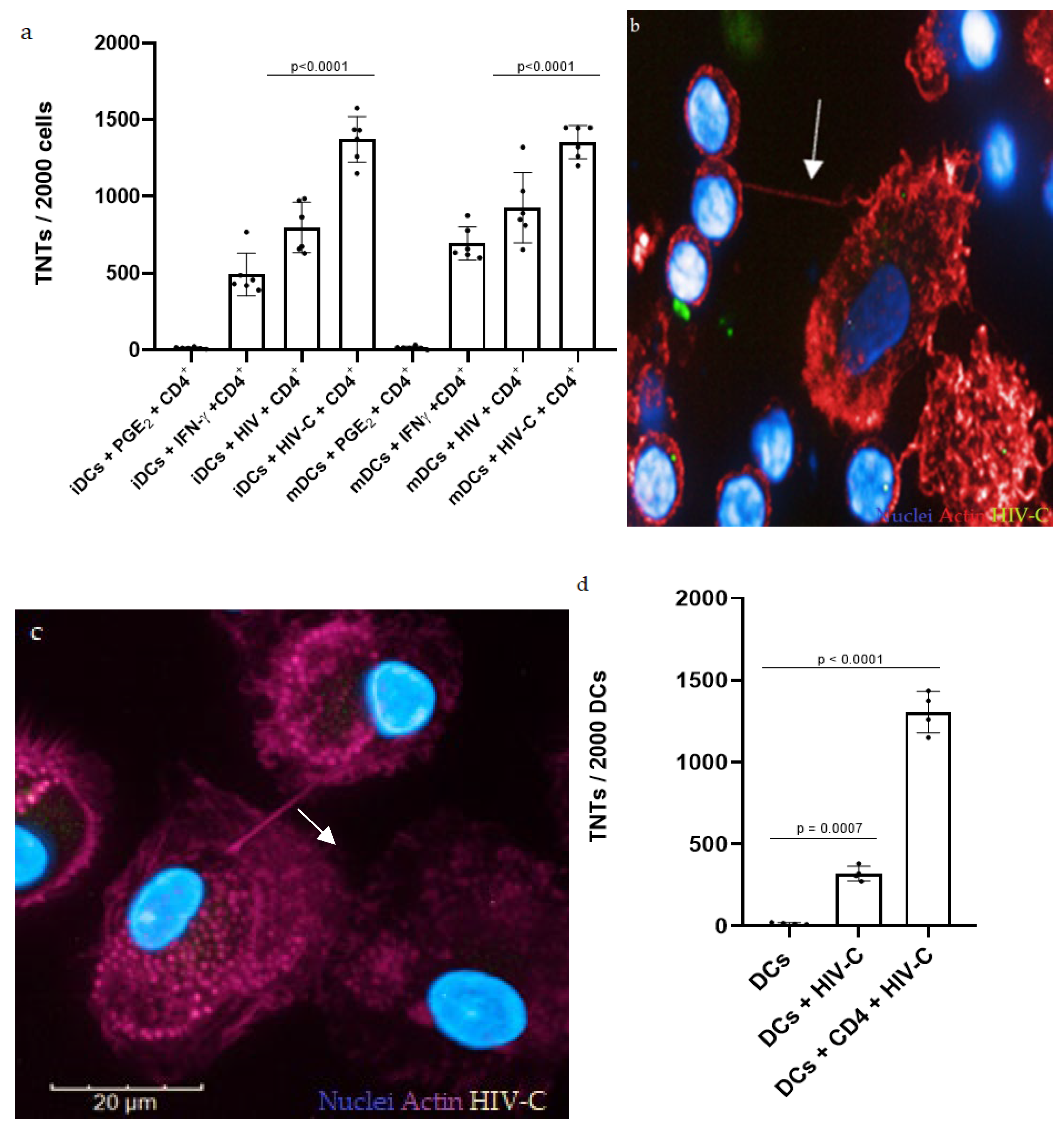
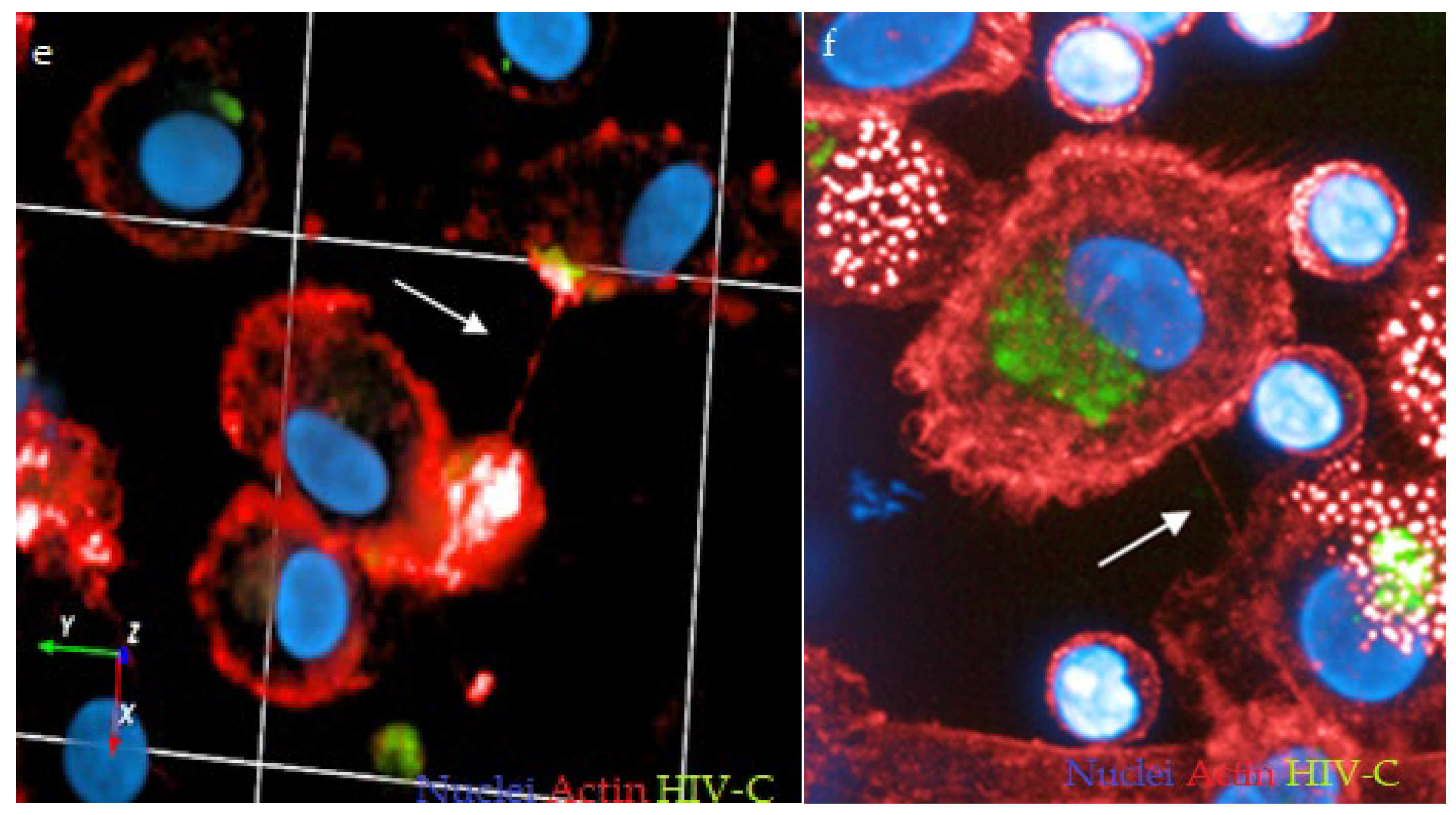
3.1. HIV-C Enhances TNT Formation in DCs and DC/CD4+ T-Cell Co-Cultures
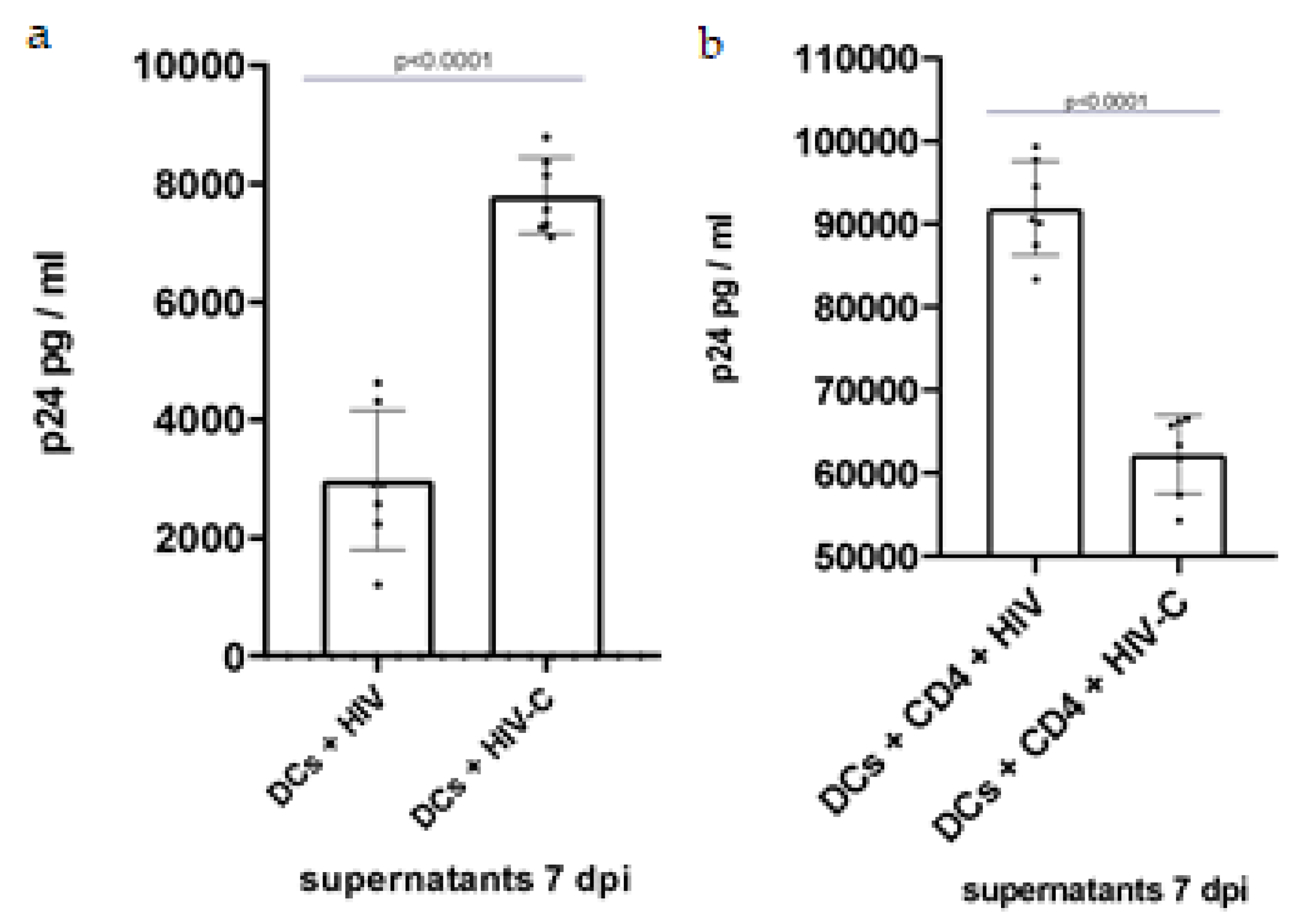
3.2. Investigating DC and DC/CD4+ T-Cell Infection with HIV and HIV-C
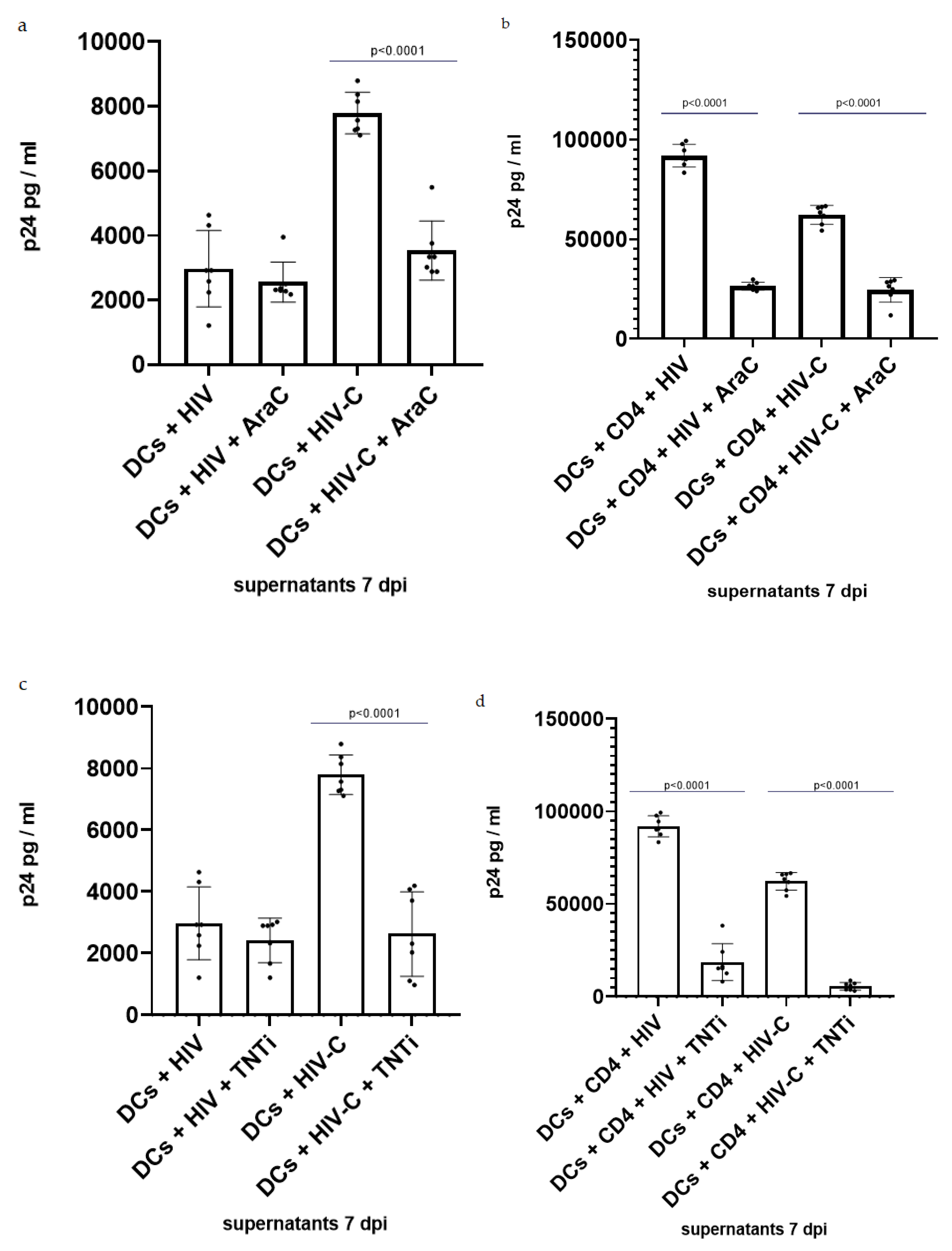
3.3. Inhibition of TNT Formation Decreases Infection in DCs and DC/CD4+ Co-Cultures
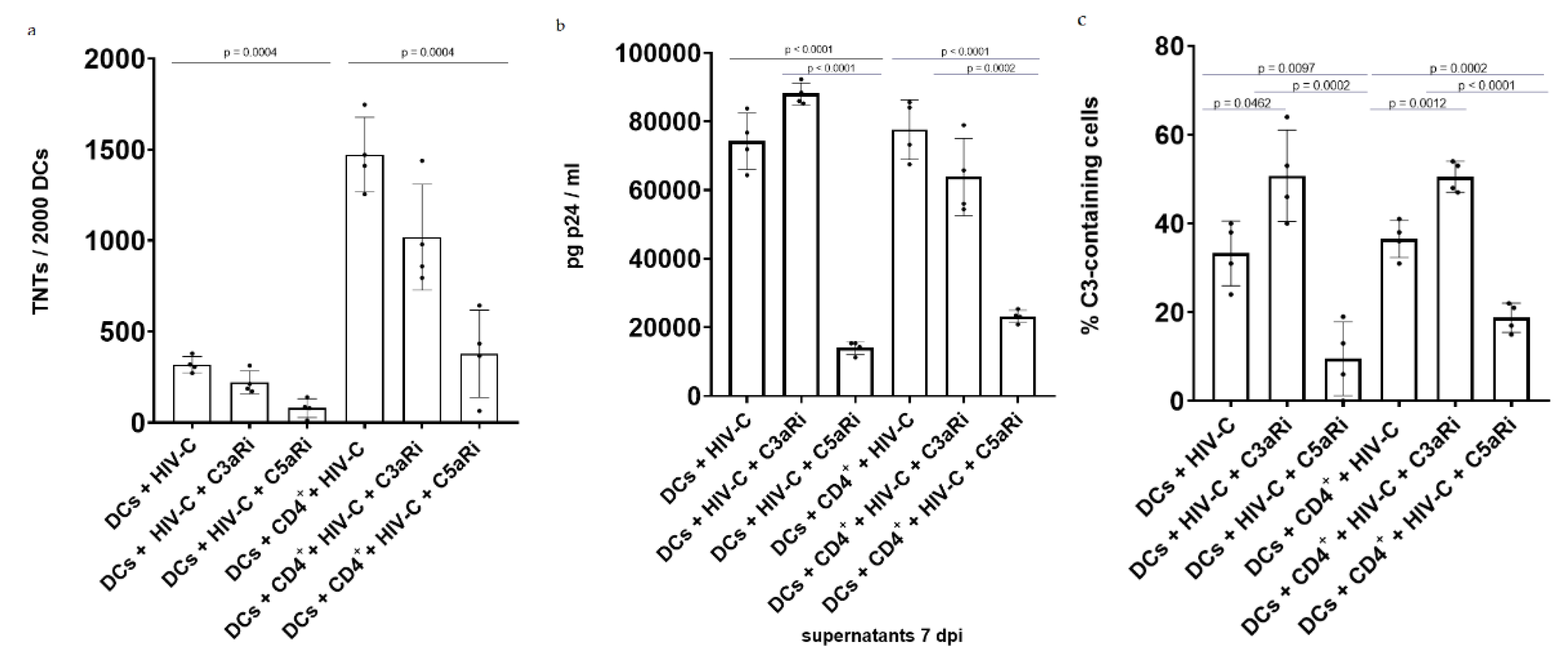
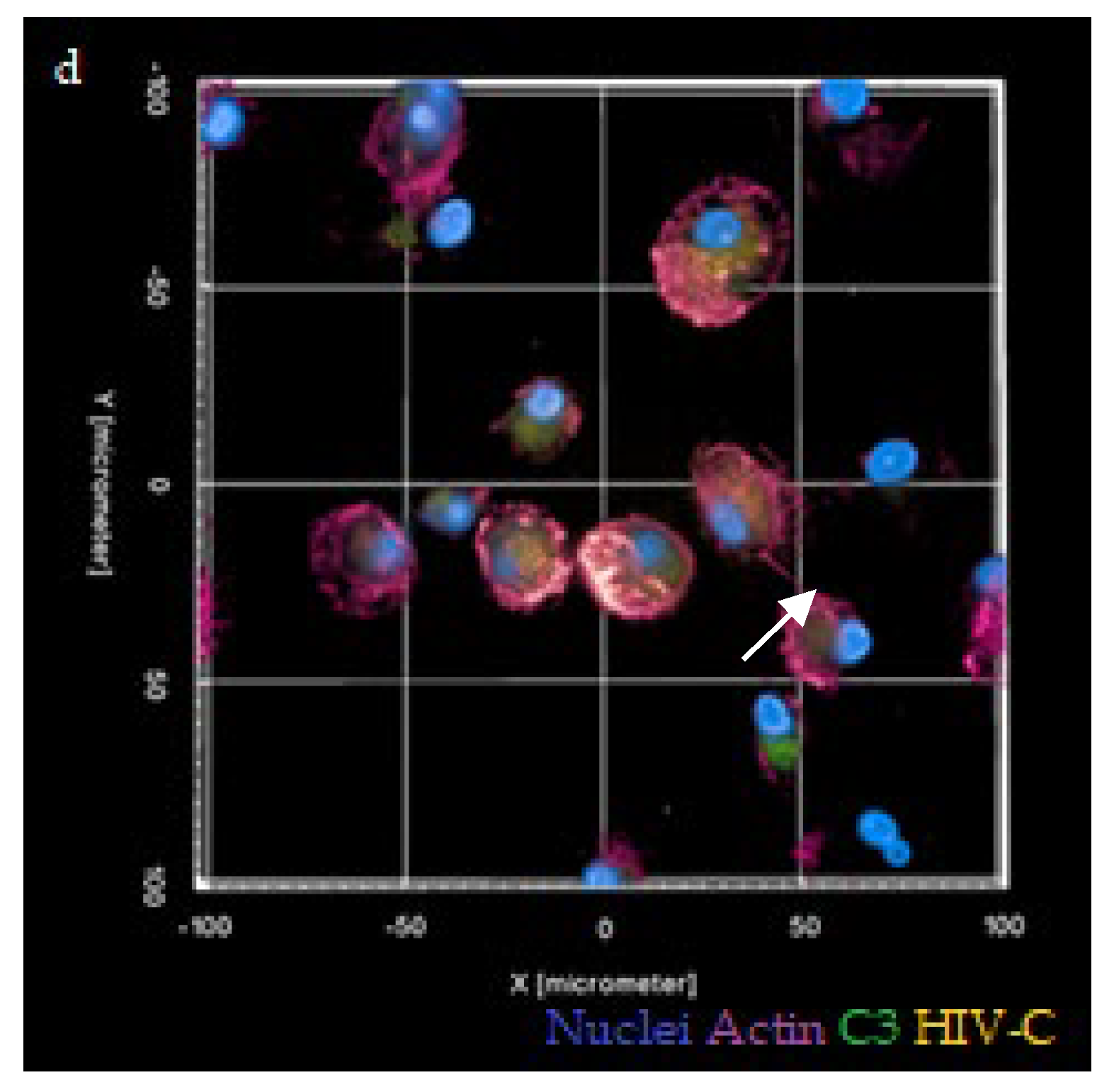
3.4. Antagonizing C5aR Reduces TNT Formation, Infection and Local C3 Production in HIV-C-Infected DCs and DC/CD4+ T-Cell Co-Cultures
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Joint United Nations Programme on HIV/AIDS. Global AIDS update. In Confronting Inequalities. Lessons for Pandemic Responses from 40 Years of AIDS; UNAIDS: Geneva, Switzerland, 2021; p. 14. [Google Scholar]
- Hemelaar, J. The origin and diversity of the HIV-1 pandemic. Trends Mol. Med. 2012, 18, 182–192. [Google Scholar] [CrossRef]
- Steinman, R.M.; Hemmi, H. Dendritic cells: Translating innate to adaptive immunity. Curr. Top. Microbiol. Immunol. 2006, 311, 17–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinman, R.M. Decisions about dendritic cells: Past, present, and future. Annu. Rev. Immunol. 2012, 30, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, R.; Richter, H.E.; Smith, P.D. Early HIV-1 target cells in human vaginal and ectocervical mucosa. Am. J. Reprod. Immunol. 2011, 65, 261–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Andrian, U.; Mempel, T. Homing and cellular traffic in lymph nodes. Nat. Rev. Immunol. 2003, 3, 867–878. [Google Scholar] [CrossRef]
- Bousso, P.; Robey, E. Dynamics of CD8+ T cell priming by dendritic cells in intact lymph nodes. Nat. Immunol. 2003, 4, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Lutz, M.; Schuler, G. Immature, semi-mature and fully mature dendritic cells: Which signals induce tolerance or immunity? Trends Immunol. 2002, 23, 445–449. [Google Scholar] [CrossRef]
- Bertram, K.M.; Botting, R.A.; Baharlou, H.; Rhodes, J.W.; Rana, H.; Graham, J.D.; Patrick, E.; Fletcher, J.; Plasto, T.M.; Truong, N.R.; et al. Identification of HIV transmitting CD11c+ human epidermal dendritic cells. Nat. Commun. 2019, 10, 2759. [Google Scholar] [CrossRef]
- Smed-Sorensen, A.; Loré, K.; Vasudevan, J.; Louder, M.K.; Andersson, J.; Mascola, J.R.; Spetz, A.L.; Koup, R.A. Differential susceptibility to human immunodeficiency virus type 1 infection of myeloid and plasmacytoid dendritic cells. J. Virol. 2005, 79, 8861–8869. [Google Scholar] [CrossRef] [Green Version]
- Kirchhoff, F. Immune evasion and counteraction of restriction factors by HIV-1 and other primate lentiviruses. Cell Host Microbe 2010, 8, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Hrecka, K.; Hao, C.; Gierszewska, M.; Swanson, S.K.; Kesik-Brodacka, M.; Srivastava, S.; Florens, L.; Washburn, M.P.; Skowronski, J. Vpx relieves inhibition of HIV-1 infection of macrophages mediated by the SAMHD1 protein. Nature 2011, 474, 658–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laguette, N.; Sobhian, B.; Casartelli, N.; Ringeard, M.; Chable-Bessia, C.; Segeral, E.; Yatim, A.; Emiliani, S.; Schwartz, O.; Benkirane, M. SAMHD1 is the dendritic-and myeloid-cell-specific HIV-1 restriction factor counteracted by Vpx. Nature 2011, 474, 654–657. [Google Scholar] [CrossRef] [PubMed]
- Descours, B.; Cribier, A.; Chable-Bessia, C.; Ayinde, D.; Rice, G.; Crow, Y.; Yatim, A.; Schwartz, O.; Laguette, N.; Benkirane, M. SAMHD1 restricts HIV-1 reverse transcription in quiescent CD4(+) T-cells. Retrovirology 2012, 9, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahouassa, H.; Daddacha, W.; Hofmann, H.; Ayinde, D.; Logue, E.C.; Dragin, L.; Bloch, N.; Maudet, C.; Bertrand, M.; Gramberg, T.; et al. SAMHD1 restricts the replication of human immunodeficiency virus type 1 by depleting the intracellular pool of deoxynucleoside triphosphates. Nat. Immunol. 2012, 13, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Goldstone, D.; Ennis-Adeniran, V.; Hedden, J.J.; Groom, H.C.T.; Rice, G.I.; Christodoulou, E.; Walker, P.A.; Kelly, G.; Haire, L.F.; Yap, M.W. HIV-1 restriction factor SAMHD1 is a deoxynucleoside triphosphate triphosphohydrolase. Nature 2011, 480, 379–382. [Google Scholar] [CrossRef]
- Posch, W.; Steger, M.; Knackmuss, U.; Blatzer, M.; Baldauf, H.M.; Doppler, W.; White, T.E.; Hörtnagl, P.; Diaz-Griffero, F.; Lass-Flörl, C.; et al. Complement-opsonized HIV-1 overcomes restriction in dendritic cells. PLoS Pathog. 2015, 11, e1005005. [Google Scholar] [CrossRef]
- Bermejo-Jambrina, M.; Blatzer, M.; Jauregui-Onieva, P.; Yordanov, T.E.; Hörtnagl, P.; Valovka, T.; Huber, L.A.; Wilflingseder, D.; Posch, W. CR4 Signaling Contributes to a DC-Driven Enhanced Immune Response Against Complement-Opsonized HIV-1. Front. Immunol. 2020, 11, 2010. [Google Scholar] [CrossRef]
- Posch, W.; Bermejo-Jambina, M.; Steger, M.; Witting, C.; Gabriel, D.; Hörtnagl, P.; Hubert, H.; Lass-Flörl, C.; Huber, L.A.; Geijtenbeek, T.B.H.; et al. Complement Potentiates Immune Sensing of HIV-1 and Early Type I Interferon Responses. mBio 2021, 12, 5. [Google Scholar] [CrossRef]
- Wilflingseder, D.; Bánki, Z.; Garcia, E.; Prünster, M.; Pfister, G.; Müllauer, B.; Nikolic, D.S.; Gassner, C.; Ammann, C.G.; Dierich, M.P.; et al. IgG Opsonization of HIV Impedes Provirus Formation in and Infection of Dendritic Cells and Subsequent Long-Term Transfer to T Cells. J. Immunol. 2007, 178, 7840–7848. [Google Scholar] [CrossRef]
- Prünster, M.; Wilflingseder, D.; Bánki, Z.; Ammann, C.G.; Müllauer, B.; Meyer, M.; Speth, C.; Dierich, M.P.; Stoiber, H. C-type lectin-independent interaction of complement opsonized HIV with monocyte-derived dendritic cells. Eur. J. Immunol. 2005, 35, 2691–2698. [Google Scholar] [CrossRef]
- Bajtay, Z.; Speth, C.; Erdei, A.; Dierich, M.P. Cutting Edge: Productive HIV-1 Infection of Dendritic Cells via Complement Receptor Type 3 (CR3, CD11b/CD18). J. Immunol. 2004, 173, 4775–4778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouhlal, H.; Chomont, N.; Réquena, M.; Nasreddine, N.; Saidi, H.; Legoff, J.; Kazatchkine, M.D.; Bélec, L.; Hochini, H. Opsonization of HIV with Complement Enhances Infection of Dendritic Cells and Viral Transfer to CD4 T Cells in a CR3 and DC-SIGN-Dependent Manner. J. Immunol. 2007, 178, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Harman, A.N.; Nasr, N.; Feetham, A.; Galoyan, A.; Alshehri, A.A.; Rambukwelle, D.; Botting, R.A.; Hiener, B.M.; Diefenbach, E.; Diefenbach, R.J.; et al. HIV Blocks Interferon Induction in Human Dendritic Cells and Macrophages by Dysregulation of TBK1. J. Virol. 2015, 89, 6575–6584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, P.; Ellegård, R.; Khalid, M.; Svanberg, C.; Govender, M.; Keita, Å.V.; Söderholm, J.D.; Myrelid, P.; Shankar, E.M.; Nyström, S.; et al. Complement opsonization of HIV affects primary infection of human colorectal mucosa and subsequent activation of T cells. eLife 2020, 9, e57869. [Google Scholar] [CrossRef]
- Stoiber, H.; Bánki, Z.; Wilflingseder, D.; Dierich, M.P. Complement HIV interactions during all steps of viral pathogenesis. Vaccine 2008, 26, 3046–3054. [Google Scholar] [CrossRef]
- Bouhlal, H.; Chomont, N.; Haeffner-Cavaillon, N.; Kazatchkine, M.; Belec, L.; Hocini, H. Opsonization of HIV-1 by semen complement enhances infection of human epithelial cells. J. Immunol. 2002, 169, 3301–3306. [Google Scholar] [CrossRef] [Green Version]
- Stoiber, H.; Pinter, C.; Siccardi, A.G.; Clivio, A.; Dierich, M.P. Efficient destruction of human immunodeficiency virus in human serum by inhibiting the protective action of complement factor H and decay accelerating factor (DAF, CD55). J. Exp. Med. 1996, 183, 307–310. [Google Scholar] [CrossRef] [Green Version]
- Frank, I.; Stoiber, H.; Godar, S.; Stockinger, H.; Steind, F.; Katinger, H.W.; Dierich, M.P. Acquisition of host cell-surface-derived molecules by HIV-1. AIDS 1996, 10, 1611–1620. [Google Scholar] [CrossRef]
- Stoiber, H.; Söderholm, A.; Wilflingseder, D.; Gusenbauer, S.; Hildgartner, A.; Dierich, M.P. Complement and antibodies: A dangerous liaison in HIV infection? Vaccine 2008, 26 (Suppl. 8), 79–85. [Google Scholar] [CrossRef]
- Ebenbichler, C.F.; Thielens, N.M.; Vornhagen, R.; Marschang, P.; Arlaud, G.J.; Dierich, M.P. Human immunodeficiency virus type 1 activates the classical pathway of complement by direct C1 binding through specific sites in the transmembrane glycoprotein gp41. J. Exp. Med. 1991, 174, 1417–1424. [Google Scholar] [CrossRef] [Green Version]
- Stoiber, H.; Thielens, N.M.; Ebenbichler, C.; Arlaud, G.J.; Dierich, M.P. The envelope glycoprotein of HIV-1 gp120 and human complement protein C1q bind to the same peptides derived from three different regions of gp41, the transmembrane glycoprotein of HIV-1, and share antigenic homology. Eur. J. Immunol. 1994, 24, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Prohászka, Z.; Hidvégi, T.; Ujhelyi, E.; Stoiber, H.; Dierich, M.P.; Süsal, C.; Füst, G. Interaction of complement and specific antibodies with the external glycoprotein 120 of HIV-1. Immunology 1995, 85, 184–189. [Google Scholar] [PubMed]
- Saifuddin, M.; Hedayati, T.; Atkinson, J.P.; Holguin, M.H.; Parker, C.J.; Spear, G.T. Human immunodeficiency virus type 1 incorporates both glycosyl phosphatidylinositol-anchored CD55 and CD59 and integral membrane CD46 at levels that protect from complement-mediated destruction. J. Gen. Virol. 1997, 78, 1907–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, S.D.; Sørensen, A.M.; Schønning, K.; Lund, O.; Nielsen, J.O.; Hansen, J.E. Complement-mediated enhancement of HIV-1 infection in peripheral blood mononuclear cells. Scand. J. Infect. Dis. 1997, 29, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Rustom, A.; Saffrich, R.; Markovic, I.; Walther, P.; Gerdes, H.H. Nanotubular highways for intercellular organelle transport. Science 2004, 303, 1007–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariazi, J.; Benowitz, A.; De Biasi, V.; Den Boer, M.L.; Cherqui, S.; Cui, H.; Douillet, N.; Eugenin, E.A.; Favre, D.; Goodman, S.; et al. Tunneling nanotubes and gap junctions-their role in long-range intercellular communication during development, health, and disease conditions. Front. Mol. Neurosci. 2017, 10, 333. [Google Scholar] [CrossRef] [PubMed]
- Rustom, A. The missing link: Does tunnelling nanotube-based supercellularity provide a new understanding of chronic and lifestyle diseases? Open Biol. 2016, 6, 160057. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Shi, Y.; You, J. Immune Cell Connection by Tunneling Nanotubes: The Impact of Intercellular Cross-Talk on the Immune Response and Its Therapeutic Applications. Mol. Pharm. 2021, 18, 772–786. [Google Scholar] [CrossRef]
- Gerdes, H.H.; Bukoreshtliev, N.V.; Barroso, J.F. Tunneling nanotubes: A new route for the exchange of components between animal cells. FEBS Lett. 2007, 581, 2194–2201. [Google Scholar] [CrossRef] [Green Version]
- Rescigno, M.; Urbano, M.; Valzasina, B.; Francolini, M.; Rotta, G.; Bonasio, R.; Granucci, F.; Kraehenbuhl, J.P.; Ricciardi-Castagnoli, P. Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nat. Immunol. 2001, 2, 361–367. [Google Scholar] [CrossRef]
- Eugenin, E.A.; Gaskill, P.J.; Berman, J.W. Tunneling nanotubes (TNT) are induced by HIV-infection of macrophages: A potential mechanism for intercellular HIV trafficking. Cell. Immunol. 2009, 254, 142–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, M.; Bhuyan, F.; Hiyoshi, M.; Noyori, O.; Nasser, H.; Miyazaki, M.; Saito, T.; Kondoh, Y.; Oada, H.; Kimura, S.; et al. Potential Role of the Formation of Tunneling Nanotubes in HIV-1 Spread in Macrophages. J. Immunol. 2016, 196, 1832–1841. [Google Scholar] [CrossRef] [PubMed]
- Lotfi, S.; Nasser, H.; Noyori, O.; Hiyoshi, M.; Takeuchi, H.; Koyanagi, Y.; Suzu, S. M-Sec facilitates intercellular transmission of HIV-1 through multiple mechanisms. Retrovirology 2020, 17, 20. [Google Scholar] [CrossRef]
- Lachambre, S.; Chopard, C.; Beaumelle, B. Preliminary characterisation of nanotubes connecting T-cells and their use by HIV-1. Biol. Cell 2014, 106, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Souriant, S.; Balboa, L.; Dupont, M.; Pingris, K.; Kviatcovsky, D.; Cougoule, C.; Lastrucci, C.; Bah, A.; Gasser, R.; Poincloux, R.; et al. Tuberculosis Exacerbates HIV-1 Infection through IL-10/STAT3-Dependent Tunneling Nanotube Formation in Macrophages. Cell Rep. 2019, 26, 3586–3599. [Google Scholar] [CrossRef] [Green Version]
- Dupont, M.; Souriant, S.; Balboa, L.; V Manh, T.P.; Pingris, K.; Rousset, S.; Cougoule, C.; Rombouts, Y.; Poincloux, R.; Ben Neji, M.; et al. Tuberculosis-associated IFN-I induces Siglec-1 on tunneling nanotubes and favors HIV-1 spread in macrophages. eLife 2020, 9, e52535. [Google Scholar] [CrossRef] [Green Version]
- Kadiu, I.; Gendelman, H.E. Human immunodeficiency virus type 1 endocytic trafficking through macrophage bridging conduits facilitates spread of infection. J. Neuroimmune Pharmacol. 2011, 6, 658–675. [Google Scholar] [CrossRef] [Green Version]
- Sowinski, S.; Jolly, C.; Berninghausen, O.; Purbhoo, M.A.; Chauveau, A.; Köhler, K.; Oddos, S.; Eissmann, P.; Brodsky, F.M.; Hopkins, C.; et al. Membrane nanotubes physically connect T cells over long distances presenting a novel route for HIV-1 transmission. Nat. Cell. Biol. 2008, 10, 211–219. [Google Scholar] [CrossRef]
- Aggarwal, A.; Hitchen, T.L.; Ootes, L.; McAllery, S.; Wong, A.; Nguyen, K.; McCluskey, A.; Robinson, P.J.; Turville, S.G. HIV infection is influenced by dynamin at 3 independent points in the viral life cycle. Traffic 2017, 18, 392–410. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, A.; Iemma, T.L.; Shih, I.; Newsome, T.P.; McAllery, S.; Cunningham, A.L.; Turville, S.G. Mobilization of HIV spread by diaphanous 2 dependent filopodia in infected dendritic cells. PLoS Pathog. 2012, 8, e1002762. [Google Scholar] [CrossRef] [Green Version]
- Zaccard, C.R.; Watkins, S.C.; Kalinski, P.; Fecek, R.J.; Yates, A.L.; Salter, R.D.; Ayyavoo, V.; Rinalso, C.R.; Mailliard, R.B. CD40L Induces Functional Tunneling Nanotube Networks Exclusively in Dendritic Cells Programmed by Mediators of Type 1 Immunity. J. Immunol. 2015, 194, 1047–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omsland, M.; Bruserud, Ø.; Gjertsen, B.T.; Andresen, V. Tunneling nanotube (TNT) formation is downregulated by cytarabine and NF-κB inhibition in acute myeloid leukemia (AML). Oncotarget 2017, 8, 7946–7963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posch, W.; Lass-Flörl, C.; Wilflingseder, D. Generation of Human Monocyte-derived Dendritic Cells from Whole Blood. J. Vis. Exp. 2016, 118, 54968. [Google Scholar] [CrossRef] [PubMed]
- Posch, W.; Cardinaud, S.; Hamimi, C.; Fletcher, A.; Muhlbacher, A.; Loacker, K.; Eichberger, P.; Dierich, M.P.; PAncino, G.; Lass-Flörl, C.; et al. Antibodies attenuate the capacity of dendritic cells to stimulate HIV-specific cytotoxic T lymphocytes. J. Allergy Clin. Immunol. 2012, 130, 1368–1374. [Google Scholar] [CrossRef] [Green Version]
- FACSDiva; Version 8; BD Biosciences: Franklin Lakes, NJ, USA, 2020; Available online: https://www.bdbiosciences.com/en-eu/products/software/instrument-software/bd-facsdiva-software#Overview (accessed on 15 December 2021).
- Purtscher, M.; Trkola, A.; Gruber, G.; Buchacher, A.; Predl, R.; Steindl, F.; Tauer, C.; Berger, R.; Barrett, N.; Jungbauer, A.; et al. broadly neutralizing human monoclonal antibody against gp41 of human immunodeficiency virus type 1. AIDS Res. Hum. Retroviruses 1994, 10, 1651–1658. [Google Scholar] [CrossRef]
- Harmony Software, Version 4.8, Perkin Elmer; Available online: https://www.perkinelmer.com/uk/product/harmony-4-8-office-hh17000001 (accessed on 20 December 2021).
- Hanna, S.J.; McCoy-Simandle, K.; Miskolci, V.; Guo, P.; Cammer, M.; Hodgson, L.; Cox, D. The Role of Rho-GTPases and actin polymerization during Macrophage Tunneling Nanotube Biogenesis. Sci. Rep. 2017, 7, 8547. [Google Scholar] [CrossRef]
- Graphpad Prism; Version 9; GraphPad Software Inc.: San Diego, CA, USA; Available online: https://www.graphpad.com (accessed on 21 January 2021).
- Gerdes, H.H.; Rustom, A. Cell-Cell Channels. In Tunneling Nanotubes; Springer: New York, NY, USA, 2006. [Google Scholar] [CrossRef]
- Omsland, M.; Pise-Masison, C.; Fujikawa, D.; Galli, V.; Fenizia, C.; Washington Parks, R.; Gjertsen, B.T.; Franchini, G.; Andresen, V. Inhibition of Tunneling Nanotube (TNT) Formation and Human T-cell Leukemia Virus Type 1 (HTLV-1) Transmission by Cytarabine. Sci. Rep. 2018, 8, 11118. [Google Scholar] [CrossRef]
- Shih, V.F.; Davis-Turak, J.; Macal, M.; Huang, J.Q.; Ponomarenko, J.; Kearns, J.D.; Yu, T.; Fagerlund, R.; Asagiri, M.; Zuniga, E.I.; et al. Control of RelB during dendritic cell activation integrates canonical and noncanonical NF-κB pathways. Nat. Immunol. 2012, 13, 1162–1170. [Google Scholar] [CrossRef] [Green Version]
- Oh, H.; Ghosh, S. NF-κB: Roles and regulation in different CD4(+) T-cell subsets. Immunol. Rev. 2013, 252, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Uhl, J.; Gujarathi, S.; Waheed, A.A.; Gordon, A.; Freed, E.O.; Gousset, K. Myosin-X is essential to the intercellular spread of HIV-1 Nef through tunneling nanotubes. J. Cell. Commun. Signal. 2019, 13, 209–224. [Google Scholar] [CrossRef]
- Mukerji, J.; Olivieri, K.C.; Misra, V.; Agopian, K.A.; Gabuzda, D. Proteomic Analysis of HIV-1 Nef Cellular Binding Partners Reveals a Role for Exocyst Complex Proteins in Mediating Enhancement of Intercellular Nanotube Formation. Retrovirology 2012, 9, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaranta, M.G.; Mattioli, B.; Spadaro, F.; Straface, E.; Giordani, L.; Ramoni, C.; Malorni, W.; Viora, M. HIV-1 Nef triggers Vav-mediated signaling pathway leading to functional and morphological differentiation of dendritic cells. FASEB J. 2003, 17, 2025–2036. [Google Scholar] [CrossRef] [PubMed]
- Dhainaut, M.; Moser, M. Regulation of immune reactivity by intercellular transfer. Front. Immunol. 2014, 5, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campana, S.; De Pasquale, C.; Carrega, P.; Ferlazzo, G.; Bonaccorsi, I. Cross-dressing: An alternative mechanism for antigen presentation. Immunol. Lett. 2015, 168, 349–354. [Google Scholar] [CrossRef]
- Zaccard, C.R.; Rinaldo, C.R.; Mailliard, R.B. Linked in: Immunologic membrane nanotube networks. J. Leukoc. Biol. 2016, 100, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Kim, J.H.; Ranjan, P.; Metcalfe, M.G.; Cao, W.; Mishina, M.; Gangappa, S.; Guo, Z.; Boyden, S.E.; Zaki, S.; et al. Influenza Virus Exploits Tunneling Nanotubes for Cell-to-Cell Spread. Sci. Rep. 2017, 7, 40360. [Google Scholar] [CrossRef]
- Schneider, C.; Oellerich, T.; Baldauf, H.M.; Schwarz, S.M.; Thomas, D.; Flick, R.; Bohnenberger, H.; Kaderali, L.; Stegmann, L.; Cremer, A.; et al. SAMHD1 is a biomarker for cytarabine response and a therapeutic target in acute myeloid leukemia. Nat. Med. 2017, 23, 250–255. [Google Scholar] [CrossRef]
- Gröschel, B.; Kaufmann, A.; Cinatl, J.; Doerr, H.W. Cytarabine treatment of human T-lymphoid cells induces decreased HIV-1 receptor expression and reduced HIV-1 susceptibility. Nucleosides Nucleotides Nucleic Acids 2001, 20, 1433–1437. [Google Scholar] [CrossRef]
- Soederholm, A.; Bánki, Z.; Wilflingseder, D.; Gassner, C.; Zwirner, J.; López-Trascasa, M.; Falkensammer, B.; Dierich, M.; Stoiber, H. HIV-1 induced generation of C5a attracts immature dendritic cells and promotes infection of autologous T cells. Eur. J. Immunol. 2007, 37, 2156–2163. [Google Scholar] [CrossRef]
- Wilflingseder, D.; Schroll, A.; Hackl, H.; Gallasch, R.; Frampton, D.; Lass-Flörl, C.; Pancino, G.; Saez-Cirion, A.; Lambotte, O.; Weiss, L.; et al. Immediate T-Helper 17 Polarization Upon Triggering CD11b/c on HIV-Exposed Dendritic Cells. J. Infect. Dis. 2015, 212, 44–56. [Google Scholar] [CrossRef]
- Posch, W.; Vosper, J.; Noureen, A.; Zaderer, V.; Witting, C.; Bertacchi, G.; Gstir, R.; Filipek, P.A.; Bonn, G.K.; Huber, L.A.; et al. C5aR inhibition of nonimmune cells suppresses inflammation and maintains epithelial integrity in SARS-CoV-2–infected primary human airway epithelia. J. Allergy Clin. Immunol. 2021, 147, 2083–2097. [Google Scholar] [CrossRef] [PubMed]
- Vaibhav, T.; Raghuram, K.; Greer, R.; Ananya, S.; Deepak, S. Role of Tunneling Nanotubes in Viral Infection, Neurodegenerative Disease, and Cancer. Front. Immunol. 2021, 12, 2256. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertacchi, G.; Posch, W.; Wilflingseder, D. HIV-1 Trans Infection via TNTs Is Impeded by Targeting C5aR. Biomolecules 2022, 12, 313. https://doi.org/10.3390/biom12020313
Bertacchi G, Posch W, Wilflingseder D. HIV-1 Trans Infection via TNTs Is Impeded by Targeting C5aR. Biomolecules. 2022; 12(2):313. https://doi.org/10.3390/biom12020313
Chicago/Turabian StyleBertacchi, Giulia, Wilfried Posch, and Doris Wilflingseder. 2022. "HIV-1 Trans Infection via TNTs Is Impeded by Targeting C5aR" Biomolecules 12, no. 2: 313. https://doi.org/10.3390/biom12020313
APA StyleBertacchi, G., Posch, W., & Wilflingseder, D. (2022). HIV-1 Trans Infection via TNTs Is Impeded by Targeting C5aR. Biomolecules, 12(2), 313. https://doi.org/10.3390/biom12020313