
Decapeptide from Potato Hydrolysate Induces Myogenic Differentiation and Ameliorates High Glucose-Associated Modulations in Protein Synthesis and Mitochondrial Biogenesis in C2C12 Cells

 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture
2.3. MTT Assay
2.4. Differentiation of C2C12 Cells
2.5. Western Blotting Analysis
2.6. Statistical Analysis
3. Results
3.1. Effect of DI-10 on Myoblast Cell Viability
3.2. DI-10 Activates ERK, Akt, mTOR, and FOXO3a Proteins and Facilitates Protein Synthesis in Myoblast Cells
3.3. DI-10 Improves the Viability of C2C12 Cells under High Glucose Stress
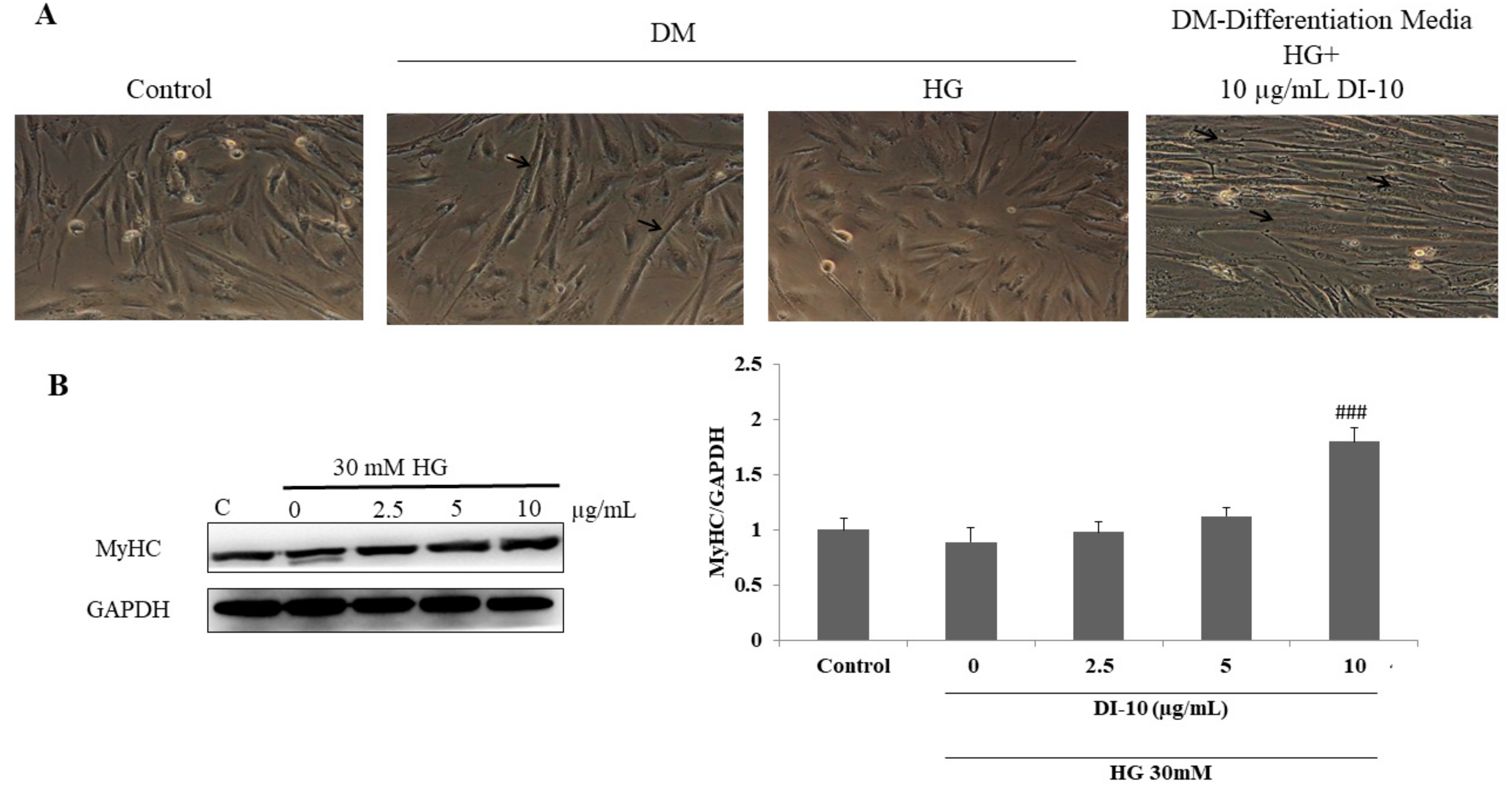
3.4. Decapeptide DI-10 Promotes Myoblast Differentiation under High-Glucose Conditions
3.5. Effect of DI-10 on Akt, mTOR, and AMPK Phosphorylation under Hyperglycemic Conditions in C2C12 Cells
3.6. DI-10 Inhibits Muscle Atrophy under Hyperglycemic Conditions through Modulating MAFbx and MuRF1
3.7. DI-10 Improves the Expression of Mitochondrial Biogenesis-Regulating Factors in Myotubes in High-Glucose Conditions through NRF-1 and TFam
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cruz-Jentoft, A.J.; Landi, F.; Schneider, S.M.; Zúñiga, C.; Arai, H.; Boirie, Y.; Chen, L.-K.; Fielding, R.A.; Martin, F.C.; Michel, J.-P. Prevalence of and interventions for sarcopenia in ageing adults: A systematic review. Report of the International Sarcopenia Initiative (EWGSOP and IWGS). Age Ageing 2014, 43, 748–759. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, S.K. Sarcopenia: A contemporary health problem among older adult populations. Nutrients 2020, 12, 1293. [Google Scholar] [CrossRef] [PubMed]
- Bowen, T.S.; Schuler, G.; Adams, V. Skeletal muscle wasting in cachexia and sarcopenia: Molecular pathophysiology and impact of exercise training. J. Cachexia Sarcopenia Muscle 2015, 6, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-J.; Chang, C.-F.; Angayarkanni, J.; Lin, W.-T. Alcalase Potato Protein Hydrolysate-PPH902 Enhances Myogenic Differentiation and Enhances Skeletal Muscle Protein Synthesis under High Glucose Condition in C2C12 Cells. Molecules 2021, 26, 6577. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, R. Nutrition and sarcopenia. J. Clin. Densitom. 2015, 18, 483–487. [Google Scholar] [CrossRef]
- Fukada, S.-I.; Akimoto, T.; Sotiropoulos, A. Role of damage and management in muscle hypertrophy: Different behaviors of muscle stem cells in regeneration and hypertrophy. Biochim. Biophys. Acta BBA Mol. Cell Res. 2020, 1867, 118742. [Google Scholar] [CrossRef]
- Rommel, C.; Bodine, S.C.; Clarke, B.A.; Rossman, R.; Nunez, L.; Stitt, T.N.; Yancopoulos, G.D.; Glass, D.J. Mediation of IGF-1-induced skeletal myotube hypertrophy by PI (3) K/Akt/mTOR and PI (3) K/Akt/GSK3 pathways. Nat. Cell Biol. 2001, 3, 1009–1013. [Google Scholar] [CrossRef]
- Drummond, M.J.; Fry, C.S.; Glynn, E.L.; Dreyer, H.C.; Dhanani, S.; Timmerman, K.L.; Volpi, E.; Rasmussen, B.B. Rapamycin administration in humans blocks the contraction-induced increase in skeletal muscle protein synthesis. J. Physiol. 2009, 587, 1535–1546. [Google Scholar] [CrossRef]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef]
- Ji, L.L.; Kang, C. Role of PGC-1α in sarcopenia: Etiology and potential intervention-a mini-review. Gerontology 2015, 61, 139–148. [Google Scholar] [CrossRef]
- Leduc-Gaudet, J.-P.; Picard, M.; Pelletier, F.S.-J.; Sgarioto, N.; Auger, M.-J.; Vallée, J.; Robitaille, R.; St-Pierre, D.H.; Gouspillou, G. Mitochondrial morphology is altered in atrophied skeletal muscle of aged mice. Oncotarget 2015, 6, 17923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilodeau, P.A.; Coyne, E.S.; Wing, S.S. The ubiquitin proteasome system in atrophying skeletal muscle: Roles and regulation. Am. J. Physiol. Cell Physiol. 2016, 311, C392–C403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, R. Ubiquitin-proteasome pathway and muscle atrophy. Muscle Atrophy 2018, 235–248. [Google Scholar]
- Altun, M.; Besche, H.C.; Overkleeft, H.S.; Piccirillo, R.; Edelmann, M.J.; Kessler, B.M.; Goldberg, A.L.; Ulfhake, B. Muscle wasting in aged, sarcopenic rats is associated with enhanced activity of the ubiquitin proteasome pathway. J. Biol. Chem. 2010, 285, 39597–39608. [Google Scholar] [CrossRef] [Green Version]
- Rom, O.; Reznick, A.Z. The role of E3 ubiquitin-ligases MuRF-1 and MAFbx in loss of skeletal muscle mass. Free Radic. Biol. Med. 2016, 98, 218–230. [Google Scholar] [CrossRef]
- Wiedmer, P.; Jung, T.; Castro, J.P.; Pomatto, L.C.; Sun, P.Y.; Davies, K.J.; Grune, T. Sarcopenia–Molecular mechanisms and open questions. Ageing Res. Rev. 2021, 65, 101200. [Google Scholar] [CrossRef] [PubMed]
- Takase, T.; Ikeuchi, S.; Inoue, T.; Mukai, R. Eriocitrin Contained in Lemon Peel Ameliorates Disuse Muscle Atrophy by Suppressing the Expression of Atrogin-1 and MuRF-1 in Denervated Mice. J. Nat. Prod. 2021, 84, 2048–2052. [Google Scholar] [CrossRef]
- Bellanti, F.; Lo Buglio, A.; Vendemiale, G. Mitochondrial Impairment in Sarcopenia. Biology 2021, 10, 31. [Google Scholar] [CrossRef]
- Kjøbsted, R.; Hingst, J.R.; Fentz, J.; Foretz, M.; Sanz, M.N.; Pehmøller, C.; Shum, M.; Marette, A.; Mounier, R.; Treebak, J.T. AMPK in skeletal muscle function and metabolism. FASEB J. 2018, 32, 1741–1777. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Meng, X.; Kang, S.Y.; Zhang, J.; Jung, H.W.; Park, Y.-K. Regulatory effects of the fruit extract of Lycium chinense and its active compound, betaine, on muscle differentiation and mitochondrial biogenesis in C2C12 cells. Biomed. Pharmacother. 2019, 118, 109297. [Google Scholar] [CrossRef]
- Ma, J.; Kang, S.Y.; Meng, X.; Kang, A.N.; Park, J.H.; Park, Y.-K.; Jung, H.W. Effects of rhizome extract of Dioscorea batatas and its active compound, allantoin, on the regulation of myoblast differentiation and mitochondrial biogenesis in C2C12 myotubes. Molecules 2018, 23, 2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Zepeda, M.U.; Sgaravatti, A.; Dent, E. Sarcopenia and post-hospital outcomes in older adults: A longitudinal study. Arch. Gerontol. Geriatr. 2017, 69, 105–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, T.-T.; Tan, Y.-N.; Ee, K.-Y.; Xiao, J.; Wong, F.-C. Seeds, fermented foods, and agricultural by-products as sources of plant-derived antibacterial peptides. Crit. Rev. Food Sci. Nutr. 2019, 59, S162–S177. [Google Scholar] [CrossRef] [PubMed]
- Lammi, C.; Aiello, G.; Boschin, G.; Arnoldi, A. Multifunctional peptides for the prevention of cardiovascular disease: A new concept in the area of bioactive food-derived peptides. J. Funct. Foods 2019, 55, 135–145. [Google Scholar] [CrossRef]
- Kudoh, K.; Matsumoto, M.; Onodera, S.; Takeda, Y.; Ando, K.; Shiomi, N. Antioxidative activity and protective effect against ethanol-induced gastric mucosal damage of a potato protein hydrolysate. J. Nutr. Sci. Vitaminol. 2003, 49, 451–455. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.Y.; Chiang, W.-D.; Pai, P.; Lin, W.-T. Potato protein hydrolysate attenuates high fat diet-induced cardiac apoptosis through SIRT1/PGC-1a/Akt signalling. J. Funct. Foods 2015, 12, 389–398. [Google Scholar] [CrossRef]
- Baskaran, R.; Poornima, P.; Huang, C.Y.; Padma, V.V. Neferine prevents NF-κB translocation and protects muscle cells from oxidative stress and apoptosis induced by hypoxia. Biofactors 2016, 42, 407–417. [Google Scholar] [CrossRef]
- Nicastro, H.; Zanchi, N.E.; da Luz, C.R.; de Moraes, W.M.; Ramona, P.; de Siqueira Filho, M.A.; Chaves, D.F.; Medeiros, A.; Brum, P.C.; Dardevet, D. Effects of leucine supplementation and resistance exercise on dexamethasone-induced muscle atrophy and insulin resistance in rats. Nutrition 2012, 28, 465–471. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.Y.; Nithiyanantham, S.; Liao, J.Y.; Lin, W.T. Bioactive peptides attenuate cardiac hypertrophy and fibrosis in spontaneously hypertensive rat hearts. J. Food Drug Anal. 2020, 28, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Lai, P.-F.; Baskaran, R.; Kuo, C.-H.; Day, C.H.; Chen, R.-J.; Ho, T.-J.; Yeh, Y.-L.; Padma, V.V.; Lai, C.-H.; Huang, C.-Y. Bioactive dipeptide from potato protein hydrolysate combined with swimming exercise prevents high fat diet induced hepatocyte apoptosis by activating PI3K/Akt in SAMP8 mouse. Mol. Biol. Rep. 2021, 48, 2629–2637. [Google Scholar] [CrossRef]
- Lin, W.T.; Nithiyanantham, S.; Hsieh, D.J.Y.; Chen, R.J.; Day, C.H.; Liao, J.Y.; Kuo, C.H.; Mahalakshmi, B.; Kuo, W.W.; Huang, C.Y. Bioactive peptides attenuate cardiac apoptosis in spontaneously hypertensive rat hearts through activation of autophagy and mitochondrial biogenesis pathway. Environ. Toxicol. 2020, 35, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Dideriksen, K.; Reitelseder, S.; Holm, L. Influence of amino acids, dietary protein, and physical activity on muscle mass development in humans. Nutrients 2013, 5, 852–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreyer, H.C.; Drummond, M.J.; Pennings, B.; Fujita, S.; Glynn, E.L.; Chinkes, D.L.; Dhanani, S.; Volpi, E.; Rasmussen, B.B. Leucine-enriched essential amino acid and carbohydrate ingestion following resistance exercise enhances mTOR signaling and protein synthesis in human muscle. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E392–E400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, D.K.; Dickinson, J.M.; Timmerman, K.L.; Drummond, M.J.; Reidy, P.T.; Fry, C.S.; Gundermann, D.M.; Rasmussen, B.B. Exercise, amino acids and aging in the control of human muscle protein synthesis. Med. Sci. Sports Exerc. 2011, 43, 2249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, M.J.; Dreyer, H.C.; Pennings, B.; Fry, C.S.; Dhanani, S.; Dillon, E.L.; Sheffield-Moore, M.; Volpi, E.; Rasmussen, B.B. Skeletal muscle protein anabolic response to resistance exercise and essential amino acids is delayed with aging. J. Appl. Physiol. 2008, 104, 1452–1461. [Google Scholar] [CrossRef] [PubMed]
- Demontis, F.; Perrimon, N. FOXO/4E-BP signaling in Drosophila muscles regulates organism-wide proteostasis during aging. Cell 2010, 143, 813–825. [Google Scholar] [CrossRef] [Green Version]
- Murtaza, G.; Khan, A.K.; Rashid, R.; Muneer, S.; Hasan, S.M.F.; Chen, J. FOXO transcriptional factors and long-term living. Oxidative Med. Cell. Longev. 2017, 2017, 3494289. [Google Scholar] [CrossRef]
- Christian, C.J.; Benian, G.M. Animal models of sarcopenia. Aging Cell 2020, 19, e13223. [Google Scholar] [CrossRef]
- Sandri, M.; Sandri, C.; Gilbert, A.; Skurk, C.; Calabria, E.; Picard, A.; Walsh, K.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 2004, 117, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Salih, D.A.; Brunet, A. FoxO transcription factors in the maintenance of cellular homeostasis during aging. Curr. Opin. Cell Biol. 2008, 20, 126–136. [Google Scholar] [CrossRef] [Green Version]
- Ferry, A.; Messéant, J.; Parlakian, A.; Lemaitre, M.; Roy, P.; Delacroix, C.; Lilienbaum, A.; Hovhannisyan, Y.; Furling, D.; Klein, A. Desmin prevents muscle wasting, exaggerated weakness and fragility, and fatigue in dystrophic mdx mouse. J. Physiol. 2020, 598, 3667–3689. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.-H.; Kim, K.M.; Oh, H.T.; Yoo, G.D.; Jeong, M.G.; Lee, H.; Park, J.; Jeong, K.; Kim, Y.K.; Ko, Y.-G. TAZ links exercise to mitochondrial biogenesis via mitochondrial transcription factor A. Nat. Commun. 2022, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.F.; Bishop, D.J. Transcription Factor Movement and Exercise-Induced Mitochondrial Biogenesis in Human Skeletal Muscle: Current Knowledge and Future Perspectives. Int. J. Mol. Sci. 2022, 23, 1517. [Google Scholar] [CrossRef]
- Jeon, S.-H.; Choung, S.-Y. Oyster Hydrolysates Attenuate Muscle Atrophy via Regulating Protein Turnover and Mitochondria Biogenesis in C2C12 Cell and Immobilized Mice. Nutrients 2021, 13, 4385. [Google Scholar] [CrossRef] [PubMed]
- Islam, H.; Bonafiglia, J.T.; Turnbull, P.C.; Simpson, C.A.; Perry, C.G.; Gurd, B.J. The impact of acute and chronic exercise on Nrf2 expression in relation to markers of mitochondrial biogenesis in human skeletal muscle. Eur. J. Appl. Physiol. 2020, 120, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Cederroth, C.R.; Vinciguerra, M.; Gjinovci, A.; Kuhne, F.; Klein, M.; Cederroth, M.; Caille, D.; Suter, M.; Neumann, D.; James, R.W. Dietary phytoestrogens activate AMP-activated protein kinase with improvement in lipid and glucose metabolism. Diabetes 2008, 57, 1176–1185. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-J.; Baskaran, R.; Chang, C.F.; Mohammedsaleh, Z.M.; Lin, W.-T. Decapeptide from Potato Hydrolysate Induces Myogenic Differentiation and Ameliorates High Glucose-Associated Modulations in Protein Synthesis and Mitochondrial Biogenesis in C2C12 Cells. Biomolecules 2022, 12, 565. https://doi.org/10.3390/biom12040565
Chen Y-J, Baskaran R, Chang CF, Mohammedsaleh ZM, Lin W-T. Decapeptide from Potato Hydrolysate Induces Myogenic Differentiation and Ameliorates High Glucose-Associated Modulations in Protein Synthesis and Mitochondrial Biogenesis in C2C12 Cells. Biomolecules. 2022; 12(4):565. https://doi.org/10.3390/biom12040565
Chicago/Turabian StyleChen, Yi-Ju, Rathinasamy Baskaran, Ching Fang Chang, Zuhair M. Mohammedsaleh, and Wan-Teng Lin. 2022. "Decapeptide from Potato Hydrolysate Induces Myogenic Differentiation and Ameliorates High Glucose-Associated Modulations in Protein Synthesis and Mitochondrial Biogenesis in C2C12 Cells" Biomolecules 12, no. 4: 565. https://doi.org/10.3390/biom12040565
APA StyleChen, Y. -J., Baskaran, R., Chang, C. F., Mohammedsaleh, Z. M., & Lin, W. -T. (2022). Decapeptide from Potato Hydrolysate Induces Myogenic Differentiation and Ameliorates High Glucose-Associated Modulations in Protein Synthesis and Mitochondrial Biogenesis in C2C12 Cells. Biomolecules, 12(4), 565. https://doi.org/10.3390/biom12040565