
The Influence of Heterochronic Non-Myeloablative Bone Marrow Transplantation on the Immune System, Frailty, General Health, and Longevity of Aged Murine Recipients

 ,
, 
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
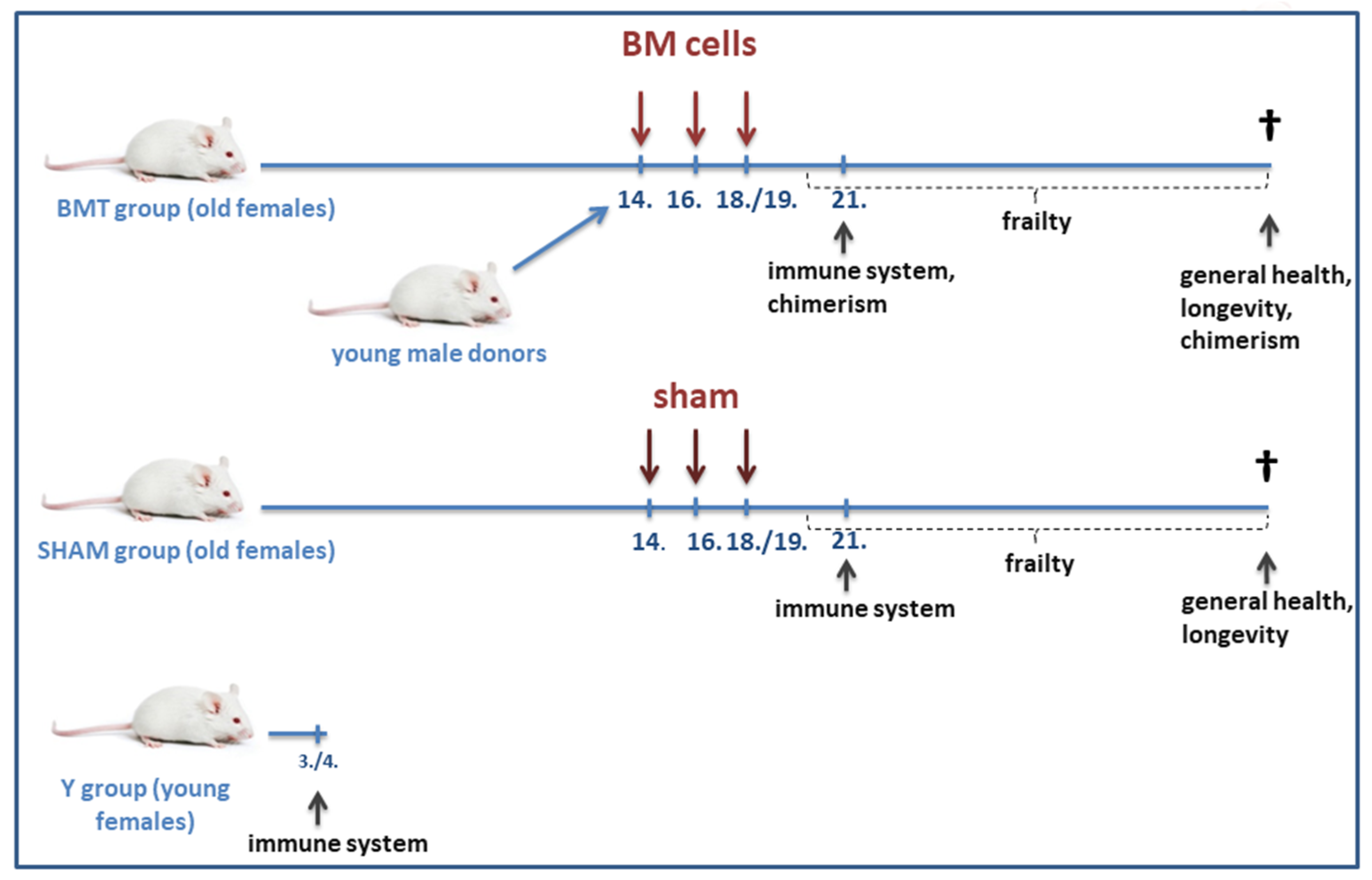
2.2. The Experimental Scheme
2.3. Heterochronic Non-Conditioned Bone Marrow Transplantation
2.4. Bone Marrow Cell Isolation
2.5. Cell Isolation from the Peritoneal Cavity
2.6. Cell Isolation from Spleen
2.7. Phenotypic Characterization with Flow Cytometry and Fluorescence-Activated Cell Sorting
2.8. DNA Isolation
2.9. Chimerism Determination with Quantitative Real-Time PCR (qPCR)
2.10. Donor Chimerism in the Hematopoietic Colony-Forming Units
2.11. Endocytosis Studies
2.12. Cytokine Release Assay
2.13. Splenocyte Proliferative Response
2.14. In Vivo Immunization
2.15. Antibody Detection Analysis
2.16. Frailty Evaluation
2.17. Pathomorphological Investigation
2.18. Longevity
2.19. Statistical Analysis
3. Results
3.1. Chimerism and HSC Number
3.2. Analysis of the Immune System
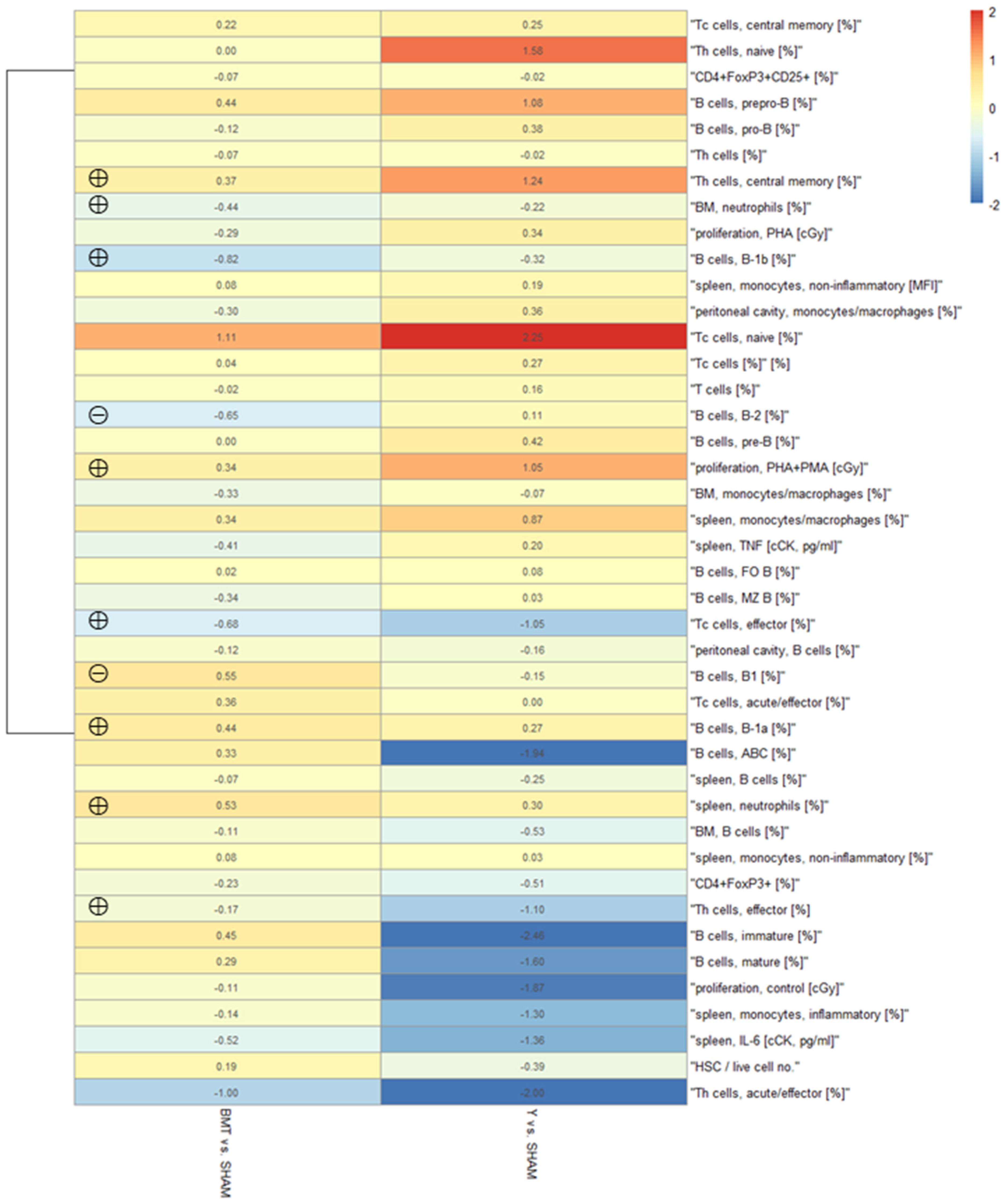
Heatmap of the Immune Parameters
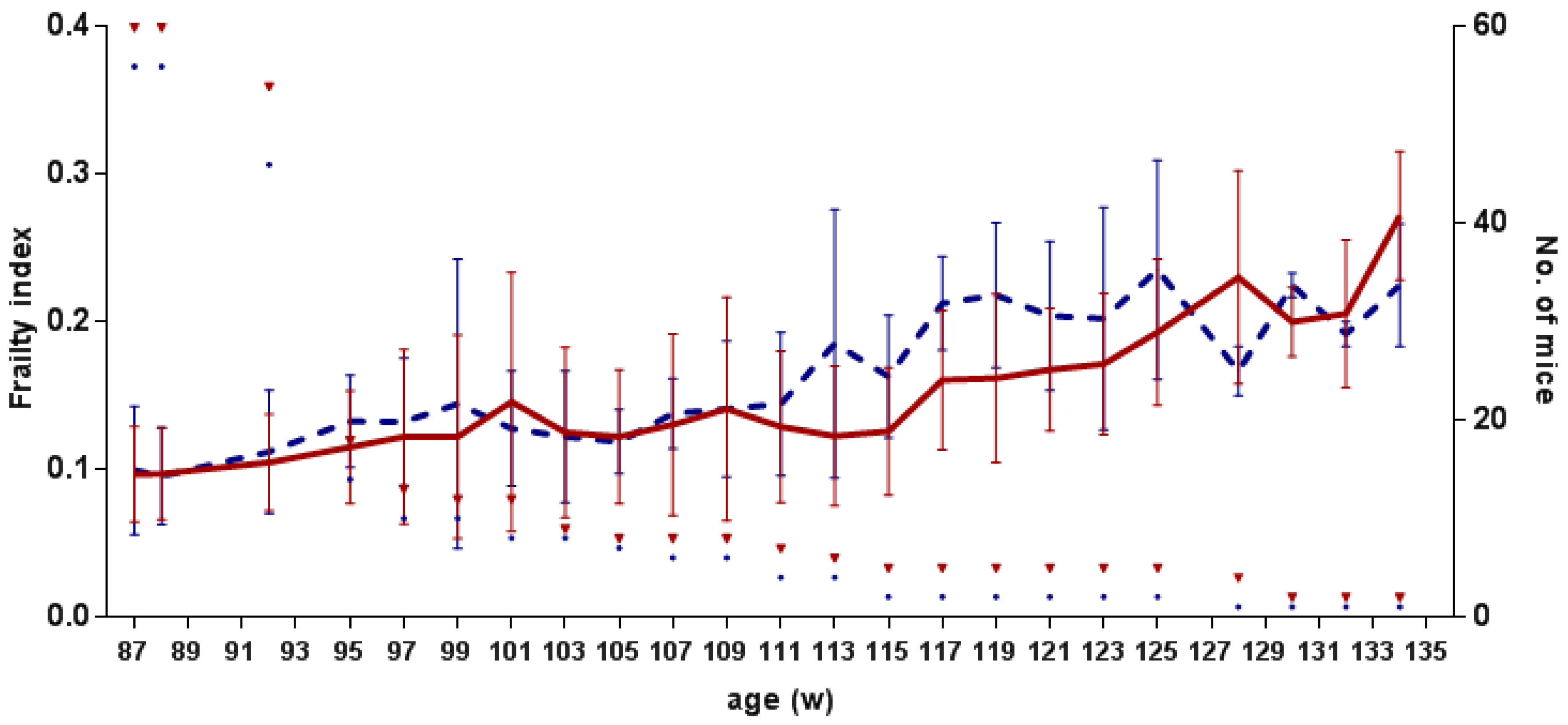
3.3. Frailty and Pathomorphological Changes
3.3.1. Frailty
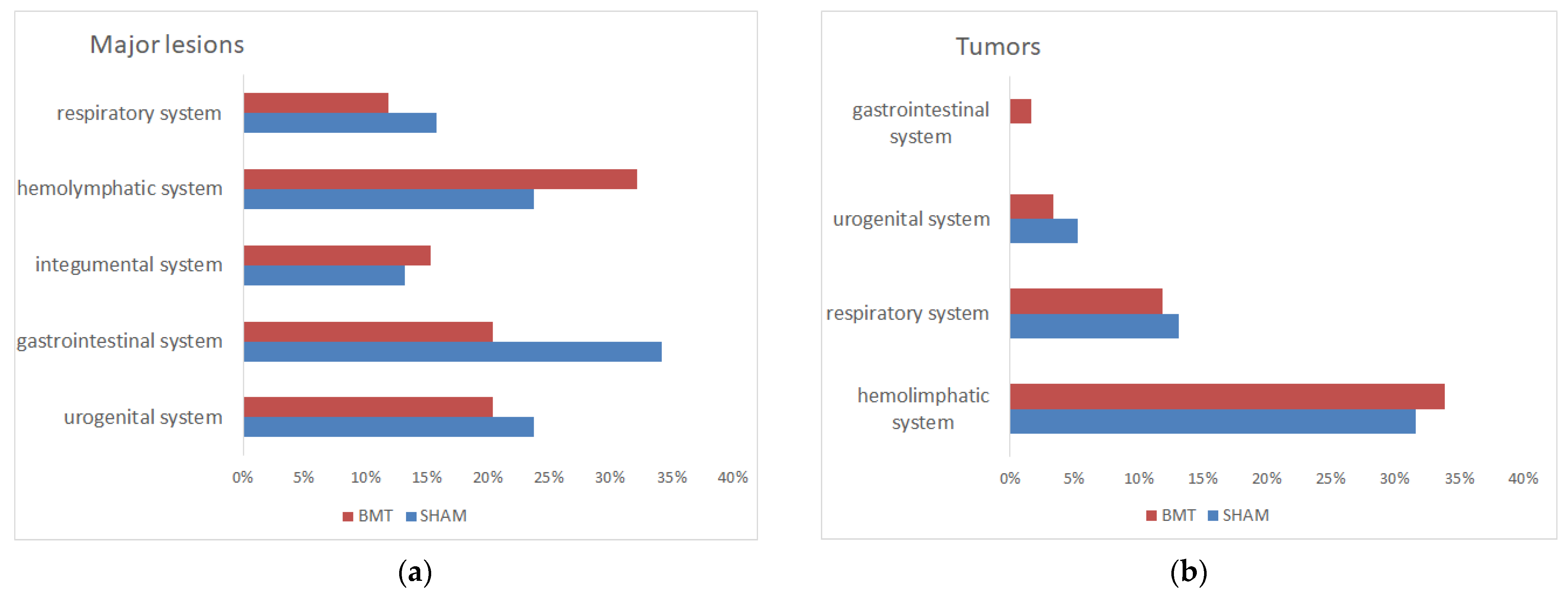
3.3.2. Pathomorphological Changes
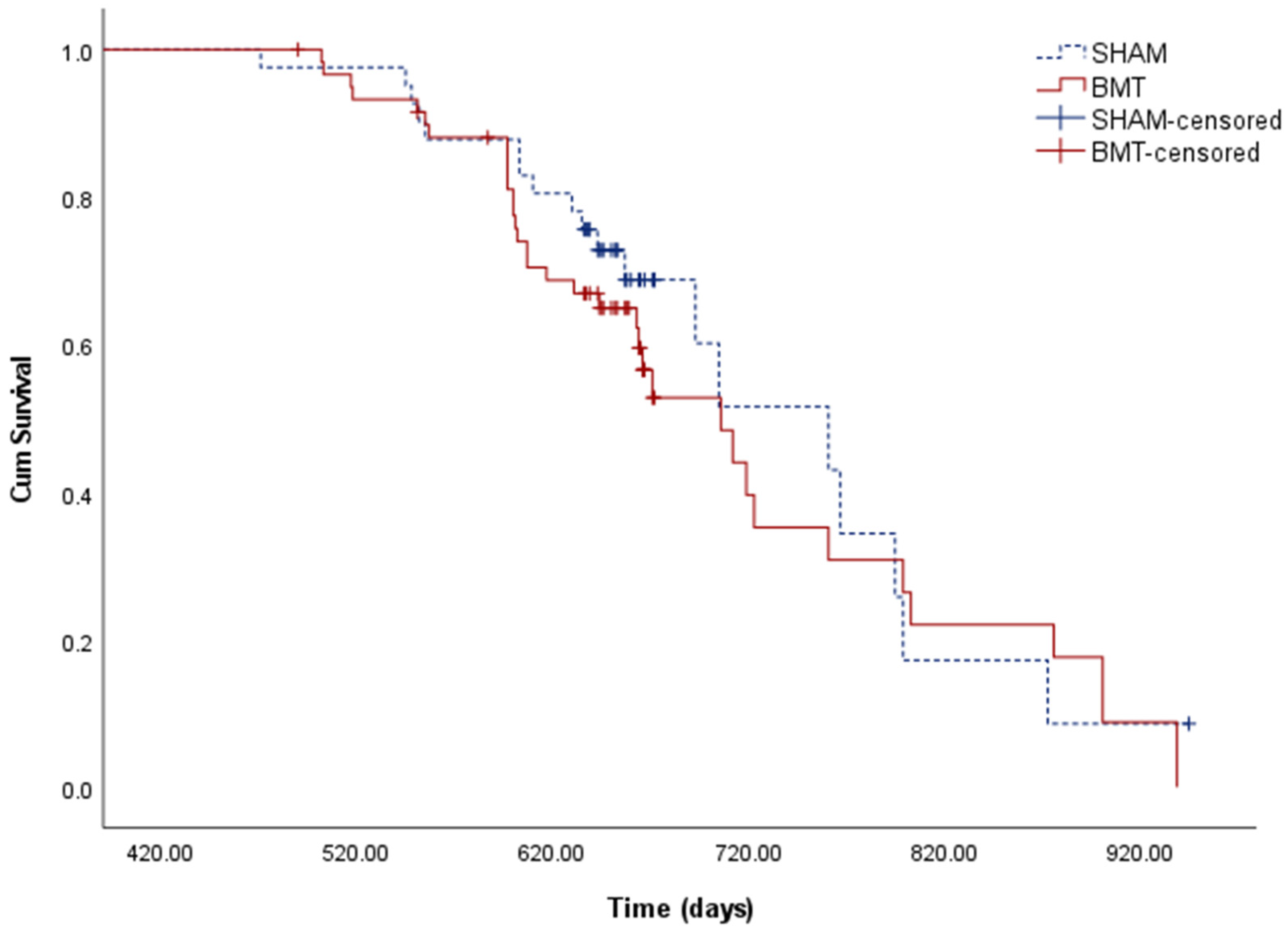
3.4. Longevity
4. Discussion
4.1. Chimerism after the Heterochronic BMT in Non-Conditioned Mice
4.2. Influence of the Heterochronic BMT on the Immune System
4.2.1. Influence of the Heterochronic BMT on the Innate Immune System
4.2.2. Influence of the Heterochronic BMT on the Adaptive Immune System
4.3. Influence of the Heterochronic BMT on the Frailty and Pathomorphological Changes
4.4. Influence of the Heterochronic BMT on the Longevity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Eurostat. Major Causes of Death, EU-28, 2016; Eurostat: Luxembourg, 2019.
- Menon, M.; Jaroslow, R.N.; Koesterer, R. The decline of cell-mediated immunity in aging mice. J Gerontol 1974, 29, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Stutman, O. Cell-mediated immunity and aging. Fed. Proc. 1974, 33, 2028–2032. [Google Scholar] [PubMed]
- Nordin, A.A.; Makinodan, T. Humoral immunity in aging. Fed. Proc. 1974, 33, 2033–2035. [Google Scholar] [PubMed]
- Albright, J.W.; Makinodan, T. Decline in the growth potential of spleen-colonizing bone marrow stem cells of long-lived aging mice. J. Exp. Med. 1976, 144, 1204–1213. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, P.A.; Bennett, M. Aging of natural and acquired immunity of mice. II. Decreased T cell responses to syngeneic tumor cells and parental-strain spleen cells. Cancer Investig. 1983, 1, 139–149. [Google Scholar] [CrossRef]
- Hirokawa, K.; Albright, J.W.; Makinodan, T. Restoration of impaired immune functions in aging animals. I. Effect of syngeneic thymus and bone marrow grafts. Clin. Immunol. Immunopathol. 1976, 5, 371–376. [Google Scholar] [CrossRef]
- Alonso-Fernandez, P.; De la Fuente, M. Role of the immune system in aging and longevity. Curr. Aging Sci. 2011, 4, 78–100. [Google Scholar] [CrossRef]
- Xu, W.; Larbi, A. Immunity and Inflammation: From Jekyll to Hyde. Exp. Gerontol. 2018, 107, 98–101. [Google Scholar] [CrossRef]
- Sharpless, N.E.; Depinho, R.A. How stem cells age and why this makes us grow old. Nat. Rev. Mol. Cell Biol. 2007, 8, 703–713. [Google Scholar] [CrossRef]
- Ahmed, A.S.; Sheng, M.H.; Wasnik, S.; Baylink, D.J.; Lau, K.W. Effect of aging on stem cells. World J. Exp. Med. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Pritz, T.; Weinberger, B.; Grubeck-Loebenstein, B. The aging bone marrow and its impact on immune responses in old age. Immunol. Lett. 2014, 162 Pt B, 310–315. [Google Scholar] [CrossRef]
- Rozman, P. The potential of non-myeloablative heterochronous autologous hematopoietic stem cell transplantation for extending a healthy life span. Geroscience 2018, 40, 221–242. [Google Scholar] [CrossRef] [PubMed]
- Rozman, P. How Could We Slow or Reverse the Human Aging Process and Extend the Healthy Life Span with Heterochronous Autologous Hematopoietic Stem Cell Transplantation. Rejuvenation Res. 2020, 23, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, S.K.; Dooner, M.S.; Quesenberry, P.J. Synchronized cell-cycle induction of engrafting long-term repopulating stem cells. Blood 1997, 90, 4646–4650. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, S.K.; Dooner, M.S.; Weier, H.U.; Frenkel, B.; Lian, J.B.; Stein, G.S.; Quesenberry, P.J. Cells capable of bone production engraft from whole bone marrow transplants in nonablated mice. Int. J. Clin. Exp. Med. 1999, 189, 729–734. [Google Scholar] [CrossRef]
- Bubnic, S.J.; Keating, A. Donor stem cells home to marrow efficiently and contribute to short- and long-term hematopoiesis after low-cell-dose unconditioned bone marrow transplantation. Exp. Hematol. 2002, 30, 606–611. [Google Scholar] [CrossRef]
- Jazbec, K.; Jez, M.; Smrekar, B.; Miceska, S.; Rozman, J.Z.; Svajger, U.; Zavrsnik, J.; Malovrh, T.; Rozman, P. Chimerism and gene therapy—Lessons learned from non-conditioned murine bone marrow transplantation models. Eur. J. Haematol. 2018, 100, 372–382. [Google Scholar] [CrossRef]
- Lountzi, D.; Henzel, K.; Jazbec, K.; Bano, D.; Krauss, S.; Rozman, P.; Ehninger, D. Effects of heterochronic, non-myeloablative bone marrow transplantation on age-related behavioural changes in mice. Mech. Ageing Dev. 2020, 191, 111327. [Google Scholar] [CrossRef]
- Justin, M.; Jez, M.; Kosir, A.; Miceska, S.; Rozman, P.; Jazbec, K. Application of the 3R principles: Vertebrae as an additional source of murine bone-marrow cells. Lab. Anim. 2021, 55, 43–52. [Google Scholar] [CrossRef]
- Ray, A.; Dittel, B.N. Isolation of mouse peritoneal cavity cells. J. Vis. Exp. 2010, 35, e1488. [Google Scholar] [CrossRef]
- An, N.; Kang, Y. Using quantitative real-time PCR to determine donor cell engraftment in a competitive murine bone marrow transplantation model. J. Vis. Exp. 2013, e50193. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, J.C.; Hildebrand, B.A.; Sun, M.; Rockwood, M.R.; Rose, R.A.; Rockwood, K.; Howlett, S.E. A clinical frailty index in aging mice: Comparisons with frailty index data in humans. J. Gerontol. -Ser. A Biol. Sci. Med. Sci. 2014, 69, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Ray, M.A.; Johnston, N.A.; Verhulst, S.; Trammell, R.A.; Toth, L.A. Identification of markers for imminent death in mice used in longevity and aging research. J. Am. Assoc. Lab. Anim. Sci. 2010, 49, 282–288. [Google Scholar] [PubMed]
- Kolde, R. Package ‘pheatmap’ in Pretty Heatmaps. Implementation of Heatmaps that Offers More Control over Dimensions and Appearance. 2018. Available online: https://cran.r-project.org/web/packages/pheatmap/index.html (accessed on 5 January 2022).
- R Core Team, R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020.
- Palmer, D.B. The Effect of Age on Thymic Function. Front. Immunol. 2013, 4, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, H.E.; Goldberg, G.L.; Chidgey, A.; Van den Brink, M.R.; Boyd, R.; Sempowski, G.D. Thymic involution and immune reconstitution. Trends Immunol. 2009, 30, 366–373. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.D.; Keating, A. Hematopoietic stem cells engraft in untreated transplant recipients. Exp. Hematol. 1993, 21, 251–256. [Google Scholar]
- Abkowitz, J.L.; Golinelli, D.; Harrison, D.E.; Guttorp, P. In vivo kinetics of murine hemopoietic stem cells. Blood 2000, 96, 3399–3405. [Google Scholar] [CrossRef]
- McCarthy, K.F. Population size and radiosensitivity of murine hematopoietic endogenous long-term repopulating cells. Blood 1997, 89, 834–841. [Google Scholar] [CrossRef]
- Colvin, G.A.; Lambert, J.F.; Abedi, M.; Hsieh, C.C.; Carlson, J.E.; Stewart, F.M.; Quesenberry, P.J. Murine marrow cellularity and the concept of stem cell competition: Geographic and quantitative determinants in stem cell biology. Leukemia 2004, 18, 575–583. [Google Scholar] [CrossRef]
- Tomay, F.; Wells, K.; Duong, L.; Tsu, J.W.; Dye, D.E.; Radley-Crabb, H.G.; Grounds, M.D.; Shavlakadze, T.; Metharom, P.; Nelson, D.J.; et al. Aged neutrophils accumulate in lymphoid tissues from healthy elderly mice and infiltrate T- and B-cell zones. Immunol. Cell Biol. 2018, 96, 831–840. [Google Scholar] [CrossRef]
- Franceschi, C.; Bonafe, M. Centenarians as a model for healthy aging. Biochem. Soc. Trans. 2003, 31, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Bueno, V.; Sant’Anna, O.A.; Lord, J.M. Ageing and myeloid-derived suppressor cells: Possible involvement in immunosenescence and age-related disease. Age 2014, 36, 9729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacs, E.J.; Palmer, J.L.; Fortin, C.F.; Fulop, T., Jr.; Goldstein, D.R.; Linton, P.J. Aging and innate immunity in the mouse: Impact of intrinsic and extrinsic factors. Trends Immunol. 2009, 30, 319–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hearps, A.C.; Martin, G.E.; Angelovich, T.A.; Cheng, W.J.; Maisa, A.; Landay, A.L.; Jaworowski, A.; Crowe, S.M. Aging is associated with chronic innate immune activation and dysregulation of monocyte phenotype and function. Aging Cell 2012, 11, 867–875. [Google Scholar] [CrossRef]
- Desai, A.; Grolleau-Julius, A.; Yung, R. Leukocyte function in the aging immune system. J. Leukoc. Biol. 2010, 87, 1001–1009. [Google Scholar] [CrossRef]
- Franceschi, C.; Garagnani, P.; Vitale, G.; Capri, M.; Salvioli, S. Inflammaging and ‘Garb-aging’. Trends Endocrinol. Metab. 2017, 28, 199–212. [Google Scholar] [CrossRef] [Green Version]
- Enioutina, E.Y.; Bareyan, D.; Daynes, R.A. A role for immature myeloid cells in immune senescence. J. Immunol. Res. 2011, 186, 697–707. [Google Scholar] [CrossRef] [Green Version]
- Hagen, M.; Derudder, E. Inflammation and the Alteration of B-Cell Physiology in Aging. Gerontology 2020, 66, 105–113. [Google Scholar] [CrossRef]
- Frasca, D.; Riley, R.L.; Blomberg, B.B. Humoral immune response and B-cell functions including immunoglobulin class switch are downregulated in aged mice and humans. Semin. Immunol. 2005, 17, 378–384. [Google Scholar] [CrossRef]
- Labi, V.; Derudder, E. Cell signaling and the aging of B cells. Exp. Gerontol. 2020, 138, 110985. [Google Scholar] [CrossRef]
- Hida, D.; Ishiguro, N.; Haneda, M.; Ishida, Y.; Suzuki, H.; Isobe, K. Intra-bone marrow bone marrow transplantation rejuvenates the B-cell lineage in aged mice. Immunol. Cell Biol. 2010, 88, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; O’Neill, P.; Naradikian, M.S.; Scholz, J.L.; Cancro, M.P. A B-cell subset uniquely responsive to innate stimuli accumulates in aged mice. Blood 2011, 118, 1294–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratliff, M.; Alter, S.; Frasca, D.; Blomberg, B.B.; Riley, R.L. In senescence, age-associated B cells secrete TNFalpha and inhibit survival of B-cell precursors. Aging Cell 2013, 12, 303–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratliff, M.; Alter, S.; McAvoy, K.; Frasca, D.; Wright, J.A.; Zinkel, S.S.; Khan, W.N.; Blomberg, B.B.; Riley, R.L. In aged mice, low surrogate light chain promotes pro-B-cell apoptotic resistance, compromises the PreBCR checkpoint, and favors generation of autoreactive, phosphorylcholine-specific B cells. Aging Cell 2015, 14, 382–390. [Google Scholar] [CrossRef] [Green Version]
- Rubtsova, K.; Rubtsov, A.V.; Cancro, M.P.; Marrack, P. Age-Associated B Cells: A T-bet-Dependent Effector with Roles in Protective and Pathogenic Immunity. J. Immunol. Res. 2015, 195, 1933–1937. [Google Scholar] [CrossRef]
- Keren, Z.; Naor, S.; Nussbaum, S.; Golan, K.; Itkin, T.; Sasaki, Y.; Schmidt-Supprian, M.; Lapidot, T.; Melamed, D. B-cell depletion reactivates B lymphopoiesis in the BM and rejuvenates the B lineage in aging. Blood 2011, 117, 3104–3112. [Google Scholar] [CrossRef] [Green Version]
- Bouamama, S.; Merzouk, H.; Medjdoub, A.; Merzouk-Saidi, A.; Merzouk, S.A. Effects of exogenous vitamins A, C, and E and NADH supplementation on proliferation, cytokines release, and cell redox status of lymphocytes from healthy aged subjects. Abbr. Appl. Physiol. Nutr. Metab. 2017, 42, 579–587. [Google Scholar] [CrossRef]
- Lages, C.S.; Lewkowich, I.; Sproles, A.; Wills-Karp, M.; Chougnet, C. Partial restoration of T-cell function in aged mice by in vitro blockade of the PD-1/ PD-L1 pathway. Aging Cell 2010, 9, 785–798. [Google Scholar] [CrossRef] [Green Version]
- Pieren, D.K.J.; Smits, N.A.M.; van de Garde, M.D.B.; Guichelaar, T. Response kinetics reveal novel features of ageing in murine T cells. Sci. Rep. 2019, 9, 5587. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Zhang, J.; Wu, H.; Tang, X.; Liu, J.; Cheng, G.; Li, P. The influences of age on T lymphocyte subsets in C57BL/6 mice. Saudi J. Biol. Sci. 2017, 24, 108–113. [Google Scholar] [CrossRef] [Green Version]
- Harms, R.; Morsey, B.; Boyer, C.W.; Fox, H.S.; Sarvetnick, N. Methamphetamine administration targets multiple immune subsets and induces phenotypic alterations suggestive of immunosuppression. PLoS ONE 2012, 7, e49897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Concepcion, A.R.; Salas, J.T.; Saez, E.; Sarvide, S.; Ferrer, A.; Portu, A.; Uriarte, I.; Hervas-Stubbs, S.; Oude Elferink, R.P.; Prieto, J.; et al. CD8+ T cells undergo activation and programmed death-1 repression in the liver of aged Ae2a,b-/- mice favoring autoimmune cholangitis. Oncotarget 2015, 6, 28588–28606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Yang, L.; Debidda, M.; Witte, D.; Zheng, Y. Cdc42 GTPase-activating protein deficiency promotes genomic instability and premature aging-like phenotypes. Proc. Natl. Acad. Sci. USA 2007, 104, 1248–1253. [Google Scholar] [CrossRef] [Green Version]
- Beswick, M.; Pachnio, A.; Lauder, S.N.; Sweet, C.; Moss, P.A. Antiviral therapy can reverse the development of immune senescence in elderly mice with latent cytomegalovirus infection. J. Virol. 2013, 87, 779–789. [Google Scholar] [CrossRef] [Green Version]
- Sckisel, G.D.; Mirsoian, A.; Minnar, C.M.; Crittenden, M.; Curti, B.; Chen, J.Q.; Blazar, B.R.; Borowsky, A.D.; Monjazeb, A.M.; Murphy, W.J. Differential phenotypes of memory CD4 and CD8 T cells in the spleen and peripheral tissues following immunostimulatory therapy. J. ImmunoTherapy Cancer 2017, 5, 33. [Google Scholar] [CrossRef] [PubMed]
- Vokali, E.; Yu, S.S.; Hirosue, S.; Rincon-Restrepo, M.; Scherer, S.; Corthesy-Henrioud, P.; Kilarski, W.W.; Mondino, A.; Zehn, D.; Hugues, S.; et al. Lymphatic endothelial cells prime naive CD8(+) T cells into memory cells under steady-state conditions. Nat. Commun. 2020, 11, 538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu Eid, R.; Ahmad, S.; Lin, Y.; Webb, M.; Berrong, Z.; Shrimali, R.; Kumai, T.; Ananth, S.; Rodriguez, P.C.; Celis, E.; et al. Enhanced Therapeutic Efficacy and Memory of Tumor-Specific CD8 T Cells by Ex Vivo PI3K-delta Inhibition. Cancer Res. 2017, 77, 4135–4145. [Google Scholar] [CrossRef] [Green Version]
- Darrigues, J.; van Meerwijk, J.P.M.; Romagnoli, P. Age-Dependent Changes in Regulatory T Lymphocyte Development and Function: A Mini-Review. Gerontology 2018, 64, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Garg, S.K.; Delaney, C.; Toubai, T.; Ghosh, A.; Reddy, P.; Banerjee, R.; Yung, R. Aging is associated with increased regulatory T-cell function. Aging Cell 2014, 13, 441–448. [Google Scholar] [CrossRef] [Green Version]
- Gregg, R.; Smith, C.M.; Clark, F.J.; Dunnion, D.; Khan, N.; Chakraverty, R.; Nayak, L.; Moss, P.A. The number of human peripheral blood CD4+ CD25high regulatory T cells increases with age. Clin. Exp. Immunol. 2005, 140, 540–546. [Google Scholar] [CrossRef]
- Fagiolo, U.; Cossarizza, A.; Scala, E.; Fanales-Belasio, E.; Ortolani, C.; Cozzi, E.; Monti, D.; Franceschi, C.; Paganelli, R. Increased cytokine production in mononuclear cells of healthy elderly people. Eur. J. Immunol. 1993, 23, 2375–2378. [Google Scholar] [CrossRef] [PubMed]
- Simioni, P.U.; Costa, E.H.; Tamashiro, W.M. Aging reduces the primary humoral response and the in vitro cytokine production in mice. Braz. J. Med. Biol. Res. 2007, 40, 1111–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, L.; Yuan, J.; Ni, S.; Zhou, Y.; Wang, X.; Chen, Y.; Zhang, S. Enhancement of adaptive immune responses of aged mice by dietary intake of beta-glucans, with special emphasis on anti-aging activity. Mol. Immunol. 2020, 117, 160–167. [Google Scholar] [CrossRef]
- Smith, T.P.; Kennedy, S.L.; Fleshner, M. Influence of age and physical activity on the primary in vivo antibody and T cell-mediated responses in men. J. Appl. Physiol. 2004, 97, 491–498. [Google Scholar] [CrossRef]
- Howlett, S.E. Assessment of Frailty in Animal Models. Interdiscip. Top. Gerontol. Geriatr. 2015, 41, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Mitnitski, A.B.; Mogilner, A.J.; Rockwood, K. Accumulation of deficits as a proxy measure of aging. Sci. World J. 2001, 1, 323–336. [Google Scholar] [CrossRef] [Green Version]
- Schlom, J.; Michalides, R.; Kufe, D.; Hehlmann, R.; Spiegelman, S.; Bentvelzen, P.; Hageman, P. A comparative study of the biological and molecular basis of murine mammary carcinoma. A model for human breast cancer. J. Nat. Cancer Inst. 1973, 51, 541–551. [Google Scholar]
- Sass, B.; Peters, R.L.; Kelloff, G.J. Differences in tumor incidence in two substrains of Claude BALB/c (BALB/cfCd) mice, emphasizing renal, mammary, pancreatic, and synovial tumors. Lab. Anim. Sci. 1976, 26, 736–741. [Google Scholar]
- Festing, M.F.; Blackmore, D.K. Life span of specified-pathogen-free (MRC category 4) mice and rats. Lab. Anim. 1971, 5, 179–192. [Google Scholar] [CrossRef]
- Mitchell, S.J.; Scheibye-Knudsen, M.; Longo, D.L.; de Cabo, R. Animal models of aging research: Implications for human aging and age-related diseases. Annu. Rev. Anim. Biosci. 2015, 3, 283–303. [Google Scholar] [CrossRef] [Green Version]
- Santulli, G.; Borras, C.; Bousquet, J.; Calza, L.; Cano, A.; Illario, M.; Franceschi, C.; Liotta, G.; Maggio, M.; Molloy, W.D.; et al. Models for preclinical studies in aging-related disorders: One is not for all. Transl. Med. UniSa 2015, 13, 4–12. [Google Scholar]
- Shen, J.; Tsai, Y.T.; Dimarco, N.M.; Long, M.A.; Sun, X.; Tang, L. Transplantation of mesenchymal stem cells from young donors delays aging in mice. Sci. Rep. 2011, 1, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.; Mohsin, S.; Khan, S.N.; Riazuddin, S. Repair of senescent myocardium by mesenchymal stem cells is dependent on the age of donor mice. J. Cell Mol. Med. 2011, 15, 1515–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.W.; Shin, I.S.; Park, S.Y.; Park, J.H.; Kim, J.S.; Yoon, E.J.; Kang, S.K.; Ra, J.C.; Hong, S.H. Reversal of serologic, immunologic, and histologic dysfunction in mice with systemic lupus erythematosus by long-term serial adipose tissue-derived mesenchymal stem cell transplantation. Arthritis Rheum. 2012, 64, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Yang, G.; Bae, D.K.; Lee, S.H.; Yang, Y.H.; Kyung, J.; Kim, D.; Choi, E.K.; Choi, K.C.; Kim, S.U.; et al. Human adipose tissue-derived mesenchymal stem cells improve cognitive function and physical activity in ageing mice. J. Neurosci. Res. 2013, 91, 660–670. [Google Scholar] [CrossRef]
- Lavasani, M.; Robinson, A.R.; Lu, A.; Song, M.; Feduska, J.M.; Ahani, B.; Tilstra, J.S.; Feldman, C.H.; Robbins, P.D.; Niedernhofer, L.J.; et al. Muscle-derived stem/progenitor cell dysfunction limits healthspan and lifespan in a murine progeria model. Nat. Commun. 2012, 3, 608. [Google Scholar] [CrossRef] [Green Version]
- Kovina, M.V.; Zuev, V.A.; Kagarlitskiy, G.O.; Khodarovich, Y.M. Effect on lifespan of high yield non-myeloablating transplantation of bone marrow from young to old mice. Front. Genet. 2013, 4, 144. [Google Scholar] [CrossRef] [Green Version]
- Kamminga, L.M.; van Os, R.; Ausema, A.; Noach, E.J.; Weersing, E.; Dontje, B.; Vellenga, E.; de Haan, G. Impaired hematopoietic stem cell functioning after serial transplantation and during normal aging. Stem Cells 2005, 23, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Selesniemi, K.; Lee, H.J.; Niikura, T.; Tilly, J.L. Young adult donor bone marrow infusions into female mice postpone age-related reproductive failure and improve offspring survival. Aging 2009, 1, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Birkenmeier, E.H.; Barker, J.E.; Vogler, C.A.; Kyle, J.W.; Sly, W.S.; Gwynn, B.; Levy, B.; Pegors, C. Increased life span and correction of metabolic defects in murine mucopolysaccharidosis type VII after syngeneic bone marrow transplantation. Blood 1991, 78, 3081–3092. [Google Scholar] [CrossRef] [Green Version]
- Singh, L.; Brennan, T.A.; Kim, J.H.; Egan, K.P.; McMillan, E.A.; Chen, Q.; Hankenson, K.D.; Zhang, Y.; Emerson, S.G.; Johnson, F.B.; et al. Long-term functional engraftment of mesenchymal progenitor cells in a mouse model of accelerated aging. Stem Cells 2013, 31, 607–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovina, M.V.; Karnaukhov, A.V.; Krasheninnikov, M.E.; Kovin, A.L.; Gazheev, S.T.; Sergievich, L.A.; Karnaukhova, E.V.; Bogdanenko, E.V.; Balyasin, M.V.; Khodarovich, Y.M.; et al. Extension of Maximal Lifespan and High Bone Marrow Chimerism After Nonmyeloablative Syngeneic Transplantation of Bone Marrow From Young to Old Mice. Front. Genet. 2019, 10, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Kueppers, S.T.; Kooreman, N.G.; Gravina, A.; Wang, D.; Tediashvili, G.; Schlickeiser, S.; Frentsch, M.; Nikolaou, C.; Thiel, A.; et al. The H-Y Antigen in Embryonic Stem Cells Causes Rejection in Syngeneic Female Recipients. Stem Cells Dev. 2020, 29, 1179–1189. [Google Scholar] [CrossRef]
- Poulos, M.G.; Ramalingam, P.; Gutkin, M.C.; Llanos, P.; Gilleran, K.; Rabbany, S.Y.; Butler, J.M. Endothelial transplantation rejuvenates aged hematopoietic stem cell function. J. Clin. Investig. 2017, 127, 4163–4178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, Y.H.; Mendez-Ferrer, S. Microenvironmental contributions to hematopoietic stem cell aging. Haematologica 2020, 105, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Frobel, J.; Rahmig, S.; Franzen, J.; Waskow, C.; Wagner, W. Epigenetic aging of human hematopoietic cells is not accelerated upon transplantation into mice. Clin. Epigen. 2018, 10, 67. [Google Scholar] [CrossRef] [PubMed]
- Weidner, C.I.; Ziegler, P.; Hahn, M.; Brummendorf, T.H.; Ho, A.D.; Dreger, P.; Wagner, W. Epigenetic aging upon allogeneic transplantation: The hematopoietic niche does not affect age-associated DNA methylation. Leukemia 2015, 29, 985–988. [Google Scholar] [CrossRef]
- Li, X.; Zeng, X.; Xu, Y.; Wang, B.; Zhao, Y.; Lai, X.; Qian, P.; Huang, H. Mechanisms and rejuvenation strategies for aged hematopoietic stem cells. J. Hematol. Oncol. Pharm. 2020, 13, 31. [Google Scholar] [CrossRef]
- Blomberg, M.; Rao, S.; Reilly, J.; Tiarks, C.; Peters, S.; Kittler, E.; Quesenberry, P. Repetitive bone marrow transplantation in nonmyeloablated recipients. Exp. Hematol. 1998, 26, 320–324. [Google Scholar]
- Takaki, T.; Hosaka, N.; Miyake, T.; Cui, W.; Nishida, T.; Inaba, M.; Ikehara, S. Presence of donor-derived thymic epithelial cells in [B6-->MRL/lpr] mice after allogeneic intra-bone marrow-bone marrow transplantation (IBM-BMT). J. Autoimmun. 2008, 31, 408–415. [Google Scholar] [CrossRef]
- Li, Y.; Hisha, H.; Inaba, M.; Lian, Z.; Yu, C.; Kawamura, M.; Yamamoto, Y.; Nishio, N.; Toki, J.; Fan, H.; et al. Evidence for migration of donor bone marrow stromal cells into recipient thymus after bone marrow transplantation plus bone grafts: A role of stromal cells in positive selection. Exp. Hematol. 2000, 28, 950–960. [Google Scholar] [CrossRef]
- Grove, J.E.; Bruscia, E.; Krause, D.S. Plasticity of bone marrow-derived stem cells. Stem Cells 2004, 22, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Huang, B. The multi-differentiation potential of peripheral blood mononuclear cells. Stem Cell Res. Ther. 2012, 3, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, A.; Saito, T.; Sato, Y.; Tsuchiya, T.; Kenjo, A.; Kimura, T.; Kanno, R.; Suzuki, H.; Kogure, M.; Hoshino, Y.; et al. Traffic of infused bone marrow cells after genetically-labeled syngeneic bone marrow transplantation following lethal irradiation in mice. Fukushima J. Med. Sci. 2008, 54, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Wang, L.; Liu, G.; Zhang, X.; Yang, J.; Pan, Y.; Luo, J. The Reparative Effects of Human Adipose-Derived Mesenchymal Stem Cells in the Chemotherapy-Damaged Thymus. Stem Cells Dev. 2019, 28, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Krueger, A. Thymus Colonization: Who, How, How Many? Arch. Immunol. Exp. 2018, 66, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Ucar, O.; Li, K.; Dvornikov, D.; Kreutz, C.; Timmer, J.; Matt, S.; Brenner, L.; Smedley, C.; Travis, M.A.; Hofmann, T.G.; et al. A Thymic Epithelial Stem Cell Pool Persists throughout Ontogeny and Is Modulated by TGF-beta. Cell Rep. 2016, 17, 448–457. [Google Scholar] [CrossRef] [Green Version]
- Chaudhry, M.S.; Velardi, E.; Malard, F.; van den Brink, M.R. Immune Reconstitution after Allogeneic Hematopoietic Stem Cell Transplantation: Time To T Up the Thymus. J. Immunol. 2017, 198, 40–46. [Google Scholar] [CrossRef]
- Fischer, J.C.; Lin, C.C.; Heidegger, S.; Wintges, A.; Schlapschy, M.; Beudert, M.; Combs, S.E.; Bassermann, F.; Skerra, A.; Haas, T.; et al. Regeneration After Radiation- and Immune-Mediated Tissue Injury Is Not Enhanced by Type III Interferon Signaling. Int. J. Radiat. Oncol. Biol. Phys. 2019, 103, 970–976. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Populations in the Spleen n (BMT) = 11, n (SHAM) = 12 | Heterochronic BMT Effects |
|---|---|
| CD11b+Ly6G-monocytes/macrophages | ns |
| CD11b+Ly6G-Ly6Chi inflammatory monocytes | ns |
| CD11b+Ly6G-Ly6C-/lo non-inflammatory monocytes | ns |
| CD11b+Ly6G+ neutrophils | ⊕ * |
| CD19+CD21/35-CD23-CD43-AA4.1-ABCs | ns |
| CD19+CD21/35+CD23+CD43-AA4.1-FO B cells | ns |
| CD19+CD21/35+CD23-CD43-AA4.1-MZ B cells | ns |
| CD3+ T cells | ns |
| CD3+CD4+ Th cells | ns |
| CD3+CD4+CD44-CD62L+ naïve Th cells | ns |
| CD3+CD4+CD44+CD62L+ central memory Th cells | ⊕ * |
| CD3+CD4+CD44+CD62L- effector/effector memory Th cells | ⊕ (*) |
| CD3+CD8a+ Tc cells | ns |
| CD3+CD8a+CD44-CD62L+ naïve Tc cells | ns |
| CD3+CD8a+CD44+CD62L+ central memory Tc cells | ns |
| CD3+CD8a+CD44+CD62L- effector/effector memory Tc cells | ⊕ * |
| CD3+CD8a+CD44-CD62L-acute/activated effector cells Tc cells | ns |
| CD4+FoxP3+ | ns |
| CD4+FoxP3+CD25+ Treg | ns |
| Cell Populations in the Bone Marrow n (BMT) = 12, n (SHAM) = 12 | Heterochronic BMT Effects |
|---|---|
| CD11b+Ly6G+ neutrophils | ⊕ * |
| CD11b+Ly6G- monocytes/macrophages | ns |
| B220loCD43+CD19-sIgM-sIgD-prepro-B | ns |
| B220loCD43-CD19+sIgM-sIgD-pre-B | ns |
| B220loCD43+CD19+sIgM-sIgD-pro-B | ns |
| B220loCD43-CD19+sIgM+sIgD-immature B cells | ns |
| B220hiCD43-CD19+sIgM+sIgD+ mature B cells | ns |
| B220+ B cells | ns |
| Cell Populations in the Peritoneal Cavity n (BMT) = 8, n (SHAM) = 6 | Heterochronic BMT Effects |
|---|---|
| B220+ B cells | ns |
| B220-CD23+ B2 cells | ⊝ * |
| B220+CD23- B1 cells | ⊝ * |
| B220+CD23-CD11b+CD5+ B1a cells | ⊕ * |
| B220+CD23-CD11b+CD5- B1b cells B220-CD11b+ monocytes/macrophages | ⊕ ** ns |
| In Vitro Endocytosis Evaluationn n (BMT) = 12, n (SHAM) = 13 | Heterochronic BMT Effects |
|---|---|
| CD11b+Ly6G-Ly6Chi inflammatory monocytes | ns |
| CD11b+Ly6G-Ly6C-/lo non-inflammatory monocytes | ns |
| CD11b+Ly6G+ neutrophils | ns |
| In Vitro Cytokine Release Assay, Spleen Cells n (BMT) = 13, n (SHAM) = 13 | Heterochronic BMT Effects |
|---|---|
| IL-6 | ns |
| TNF | ns |
| In Vitro Cytokine Release Assay, Peritoneal Cells n (BMT) = 10, n (SHAM) = 10 | Heterochronic BMT Effects |
|---|---|
| IL-6 | ns |
| MCP-1 | ns |
| TNF | ns |
| In Vitro Splenocyte Proliferative Response n (BMT) = 15, n (SHAM) = 15 | Heterochronic BMT Effects |
|---|---|
| PHA | ns |
| PHA+PMA | ⊕ (*) |
| In Vivo Immunization n (BMT) = 9, n (SHAM) = 10 | Heterochronic BMT Effects |
|---|---|
| Absorbance at 105 titre | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jazbec, K.; Jež, M.; Švajger, U.; Smrekar, B.; Miceska, S.; Rajčevič, U.; Justin, M.; Završnik, J.; Malovrh, T.; Švara, T.; et al. The Influence of Heterochronic Non-Myeloablative Bone Marrow Transplantation on the Immune System, Frailty, General Health, and Longevity of Aged Murine Recipients. Biomolecules 2022, 12, 595. https://doi.org/10.3390/biom12040595
Jazbec K, Jež M, Švajger U, Smrekar B, Miceska S, Rajčevič U, Justin M, Završnik J, Malovrh T, Švara T, et al. The Influence of Heterochronic Non-Myeloablative Bone Marrow Transplantation on the Immune System, Frailty, General Health, and Longevity of Aged Murine Recipients. Biomolecules. 2022; 12(4):595. https://doi.org/10.3390/biom12040595
Chicago/Turabian StyleJazbec, Katerina, Mojca Jež, Urban Švajger, Boštjan Smrekar, Simona Miceska, Uroš Rajčevič, Mojca Justin, Janja Završnik, Tadej Malovrh, Tanja Švara, and et al. 2022. "The Influence of Heterochronic Non-Myeloablative Bone Marrow Transplantation on the Immune System, Frailty, General Health, and Longevity of Aged Murine Recipients" Biomolecules 12, no. 4: 595. https://doi.org/10.3390/biom12040595
APA StyleJazbec, K., Jež, M., Švajger, U., Smrekar, B., Miceska, S., Rajčevič, U., Justin, M., Završnik, J., Malovrh, T., Švara, T., Gombač, M., Ramšak, Ž., & Rožman, P. (2022). The Influence of Heterochronic Non-Myeloablative Bone Marrow Transplantation on the Immune System, Frailty, General Health, and Longevity of Aged Murine Recipients. Biomolecules, 12(4), 595. https://doi.org/10.3390/biom12040595