
Effects of Risperidone and Prenatal Poly I:C Exposure on GABAA Receptors and AKT-GSK3β Pathway in the Ventral Tegmental Area of Female Juvenile Rats


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatment
2.2. Brain Dissection
2.3. RNA Isolation and Gene Expression Analysis by Real-Time qPCR
2.4. Statistical Analysis
3. Results
3.1. Effects on the GABAergic Markers
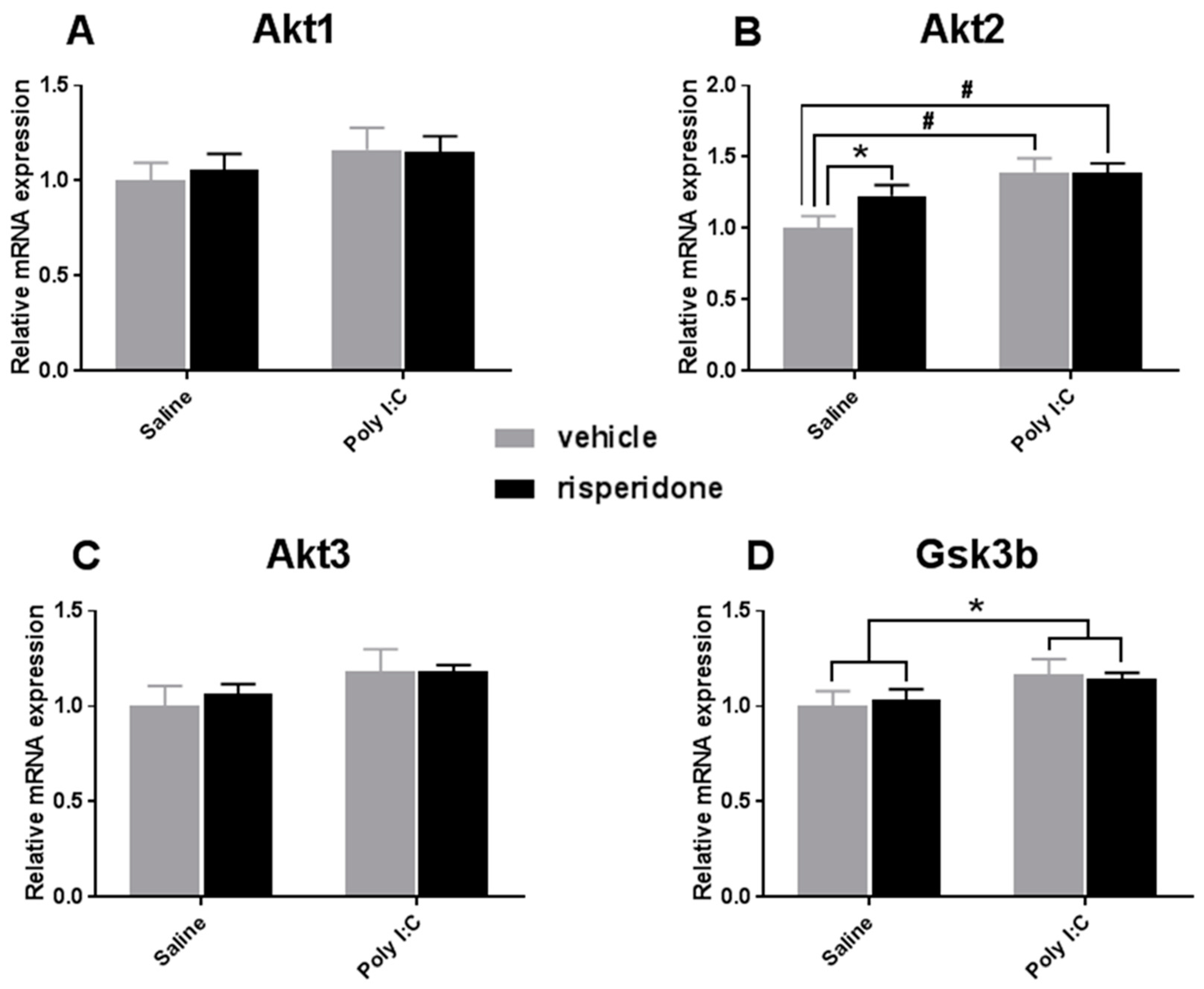
3.2. Effects on Akt-GSK3β Signaling Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, A.S.; Susser, E.S. In utero infection and adult schizophrenia. Ment. Retard. Dev. Disabil. Res. Rev. 2002, 8, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.S.; Derkits, E.J. Prenatal infection and schizophrenia: A review of epidemiologic and translational studies. Am. J. Psychiatry 2010, 167, 261–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buka, S.L.; Cannon, T.D.; Torrey, E.F.; Yolken, R.H. Maternal exposure to herpes simplex virus and risk of psychosis among adult offspring. Biol. Psychiatry 2008, 63, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Khandaker, G.M.; Zimbron, J.; Lewis, G.; Jones, P. Prenatal maternal infection, neurodevelopment and adult schizophrenia: A systematic review of population-based studies. Psychol. Med. 2013, 43, 239–257. [Google Scholar] [CrossRef] [Green Version]
- Haddad, F.L.; Patel, S.V.; Schmid, S. Maternal Immune Activation by Poly I:C as a preclinical Model for Neurodevelopmental Disorders: A focus on Autism and Schizophrenia. Neurosci. Biobehav. Rev. 2020, 113, 546–567. [Google Scholar] [CrossRef]
- Meyer, U. Prenatal poly(i:C) exposure and other developmental immune activation models in rodent systems. Biol. Psychiatry 2014, 75, 307–315. [Google Scholar] [CrossRef]
- Murray, K.N.; Edye, M.E.; Manca, M.; Vernon, A.C.; Oladipo, J.M.; Fasolino, V.; Harte, M.; Mason, V.; Grayson, B.; McHugh, P.C.; et al. Evolution of a maternal immune activation (mIA) model in rats: Early developmental effects. Brain Behav. Immun. 2018, 75, 48–59. [Google Scholar] [CrossRef] [Green Version]
- Reisinger, S.; Khan, D.; Kong, E.; Berger, A.; Pollak, A.; Pollak, D. The poly(I:C)-induced maternal immune activation model in preclinical neuropsychiatric drug discovery. Pharmacol. Ther. 2015, 149, 213–226. [Google Scholar] [CrossRef] [Green Version]
- Gogos, A.; Sbisa, A.; Witkamp, D.; Buuse, M.V.D. Sex differences in the effect of maternal immune activation on cognitive and psychosis-like behaviour in Long Evans rats. Eur. J. Neurosci. 2020, 52, 2614–2626. [Google Scholar] [CrossRef]
- Kokras, N.; Dalla, C. Sex differences in animal models of psychiatric disorders. Br. J. Pharmacol. 2014, 171, 4595–4619. [Google Scholar] [CrossRef]
- Prendergast, B.J.; Onishi, K.G.; Zucker, I. Female mice liberated for inclusion in neuroscience and biomedical research. Neurosci. Biobehav. Rev. 2014, 40, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Lian, J.; Hodgson, J.; Zhang, W.; Deng, C. Prenatal Poly I:C Challenge Affects Behaviors and Neurotransmission via Elevated Neuroinflammation Responses in Female Juvenile Rats. Int. J. Neuropsychopharmacol. 2022, 25, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Bergdolt, L.; Dunaevsky, A. Brain changes in a maternal immune activation model of neurodevelopmental brain disorders. Prog. Neurobiol. 2019, 175, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Osborne, A.L.; Solowij, N.; Babic, I.; Lum, J.S.; Newell, K.; Huang, X.-F.; Weston-Green, K. Effect of cannabidiol on endocannabinoid, glutamatergic and GABAergic signalling markers in male offspring of a maternal immune activation (poly I:C) model relevant to schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 95, 109666. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Pan, B.; Engel, M.; Huang, X.-F. Neuregulin-1 signalling and antipsychotic treatment. Psychopharmacology 2013, 226, 201–215. [Google Scholar] [CrossRef]
- Ginovart, N.; Kapur, S. Role of dopamine D(2) receptors for antipsychotic activity. Curr. Antipsychotics 2012, 212, 27–52. [Google Scholar] [CrossRef]
- Howes, O.; Kapur, S. The dopamine hypothesis of schizophrenia: Version III--the final common pathway. Schizophr. Bull. 2009, 35, 549–562. [Google Scholar] [CrossRef] [Green Version]
- Morikawa, H.; Paladini, C. Dynamic regulation of midbrain dopamine neuron activity: Intrinsic, synaptic, and plasticity mechanisms. Neuroscience 2011, 198, 95–111. [Google Scholar] [CrossRef] [Green Version]
- Pan, B.; Deng, C. Modulation by chronic antipsychotic administration of PKA- and GSK3β-mediated pathways and the NMDA receptor in rat ventral midbrain. Psychopharmacology 2019, 236, 2687–2697. [Google Scholar] [CrossRef]
- Benedetti, F.; Poletti, S.; Radaelli, D.; Bernasconi, A.; Cavallaro, R.; Falini, A.; Lorenzi, C.; Pirovano, A.; Dallaspezia, S.; Locatelli, C.; et al. Temporal lobe grey matter volume in schizophrenia is associated with a genetic polymorphism influencing glycogen synthase kinase 3-β activity. Genes Brain Behav. 2010, 9, 365–371. [Google Scholar] [CrossRef]
- Benedetti, F.; Bollettini, I.; Barberi, I.; Radaelli, D.; Poletti, S.; Locatelli, C.; Pirovano, A.; Lorenzi, C.; Falini, A.; Colombo, C.; et al. Lithium and GSK3-β promoter gene variants influence white matter microstructure in bipolar disorder. Neuropsychopharmacology 2013, 38, 313–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freyberg, Z.; Ferrando, S.J.; Javitch, J. Roles of the Akt/GSK-3 and Wnt signaling pathways in schizophrenia and antipsychotic drug action. Am. J. Psychiatry 2010, 167, 388–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, T.D.; Manji, H.K. Glycogen synthase kinase-3: A putative molecular target for lithium mimetic drugs. Neuropsychopharmacology 2005, 30, 1223–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karege, F.; Méary, A.; Perroud, N.; Jamain, S.; Leboyer, M.; Ballmann, E.; Fernandez, R.; Malafosse, A.; Schürhoff, F. Genetic overlap between schizophrenia and bipolar disorder: A study with AKT1 gene variants and clinical phenotypes. Schizophr. Res. 2012, 135, 8–14. [Google Scholar] [CrossRef]
- Alimohamad, H.; Sutton, L.; Mouyal, J.; Rajakumar, N.; Rushlow, W.J. The effects of antipsychotics on β-catenin, glycogen synthase kinase-3 and dishevelled in the ventral midbrain of rats. J. Neurochem. 2005, 95, 513–525. [Google Scholar] [CrossRef]
- Alimohamad, H.; Rajakumar, N.; Seah, Y.-H.; Rushlow, W. Antipsychotics alter the protein expression levels of β-catenin and GSK-3 in the rat medial prefrontal cortex and striatum. Biol. Psychiatry 2005, 57, 533–542. [Google Scholar] [CrossRef]
- Allen, J.A.; Yost, J.M.; Setola, V.; Chen, X.; Sassano, M.F.; Chen, M.; Peterson, S.; Yadav, P.N.; Huang, X.-P.; Feng, B.; et al. Discovery of β-Arrestin—Biased dopamine D2 ligands for probing signal transduction pathways essential for antipsychotic efficacy. Proc. Natl. Acad. Sci. USA 2011, 108, 18488–18493. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, J.-M.; Gainetdinov, R.R.; Caron, M.G. Akt/GSK3 signaling in the action of psychotropic drugs. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 327–347. [Google Scholar] [CrossRef] [Green Version]
- Emamian, E.S. AKT/GSK3 signaling pathway and schizophrenia. Front. Mol. Neurosci. 2012, 5, 33. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Rosborough, K.M.; Friedman, A.B.; Zhu, W.; Roth, K. Regulation of mouse brain glycogen synthase kinase-3 by atypical antipsychotics. Int. J. Neuropsychopharmacol. 2007, 10, 7–19. [Google Scholar] [CrossRef]
- Pan, B.; Huang, X.F.; Deng, C. Chronic administration of aripiprazole activates GSK3β-dependent signalling pathways and up-regulates GABAA receptor expression and CREB1 activity in rats. Sci. Rep. 2016, 6, 30040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, B.; Huang, X.-F.; Deng, C. Aripiprazole and Haloperidol Activate GSK3β-Dependent Signalling Pathway Differentially in Various Brain Regions of Rats. Int. J. Mol. Sci. 2016, 17, 459. [Google Scholar] [CrossRef] [PubMed]
- Sutton, L.P.; Rushlow, W.J. The dopamine D2 receptor regulates Akt and GSK-3 via Dvl-3. Int. J. Neuropsychopharmacol. 2012, 15, 965–979. [Google Scholar] [CrossRef] [Green Version]
- Karanges, E.A.; Stephenson, C.P.; McGregor, I.S. Longitudinal trends in the dispensing of psychotropic medications in Australia from 2009–2012: Focus on children, adolescents and prescriber specialty. Aust. N. Z. J. Psychiatry 2014, 48, 917–931. [Google Scholar] [CrossRef] [PubMed]
- Memarzia, J.; Tracy, D.; Giaroli, G. The use of antipsychotics in preschoolers: A veto or a sensible last option? J. Psychopharmacol. 2014, 28, 303–319. [Google Scholar] [CrossRef] [PubMed]
- Olfson, M.; Blanco, C.; Wang, S.; Laje, G.; Correll, C.U. National trends in the mental health care of children, adolescents, and adults by office-based physicians. JAMA Psychiatry 2014, 71, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Caccia, S. Safety and pharmacokinetics of atypical antipsychotics in children and adolescents. Paediatr. Drugs 2013, 15, 217–233. [Google Scholar] [CrossRef]
- Fraguas, D.; Correll, C.U.; Merchán-Naranjo, J.; Rapado-Castro, M.; Parellada, M.; Moreno, C.; Arango, C. Efficacy and safety of second-generation antipsychotics in children and adolescents with psychotic and bipolar spectrum disorders: Comprehensive review of prospective head-to-head and placebo-controlled comparisons. Eur. Neuropsychopharmacol. 2011, 21, 621–645. [Google Scholar] [CrossRef]
- Hoekstra, P.J. Risperidone for non-psychotic disorders in paediatric patients: Which child is to benefit? Dev. Med. Child Neurol. 2014, 56, 919–920. [Google Scholar] [CrossRef] [Green Version]
- Olfson, M.; Crystal, S.; Huang, C.; Gerhard, T. Trends in antipsychotic drug use by very young, privately insured children. J. Am. Acad. Child Adolesc. Psychiatry 2010, 49, 13–23. [Google Scholar] [CrossRef]
- Seida, J.C.; Schouten, J.R.; Boylan, K.; Newton, A.S.; Mousavi, S.S.; Beaith, A.; Vandermeer, B.; Dryden, D.M.; Carrey, N. Antipsychotics for children and young adults: A comparative effectiveness review. Pediatrics 2012, 129, e771–e784. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shaw, S.R. Efficacy of risperidone in managing maladaptive behaviors for children with autistic spectrum disorder: A meta-analysis. J. Pediatr. Health Care 2012, 26, 291–299. [Google Scholar] [CrossRef] [PubMed]
- De Santis, M.; Lian, J.; Huang, X.-F.; Deng, C. Early Antipsychotic Treatment in Juvenile Rats Elicits Long-Term Alterations to the Dopamine Neurotransmitter System. Int. J. Mol. Sci. 2016, 17, 1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, B.; Lian, J.; Deng, C. Chronic antipsychotic treatment differentially modulates protein kinase A- and glycogen synthase kinase 3 beta-dependent signaling pathways, N-methyl-D-aspartate receptor and γ-aminobutyric acid A receptors in nucleus accumbens of juvenile rats. J. Psychopharmacol. 2018, 32, 1252–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, B.; Chen, J.; Lian, J.; Huang, X.-F.; Deng, C. Unique Effects of Acute Aripiprazole Treatment on the Dopamine D2 Receptor Downstream cAMP-PKA and Akt-GSK3β Signalling Pathways in Rats. PLoS ONE 2015, 10, e0132722. [Google Scholar] [CrossRef] [Green Version]
- Paxinos, G.; Watson, C. The Rat Brain in Sterotaxic Coordinates, 6th ed.; Academic Press: New York, NY, USA, 2007. [Google Scholar]
- Richetto, J.; Calabrese, F.; Riva, M.A.; Meyer, U. Prenatal immune activation induces maturation-dependent alterations in the prefrontal GABAergic transcriptome. Schizophr. Bull. 2014, 40, 351–361. [Google Scholar] [CrossRef] [Green Version]
- De Jonge, J.C.; Vinkers, C.H.; Pol, H.E.H.; Marsman, A. GABAergic Mechanisms in Schizophrenia: Linking Postmortem and In Vivo Studies. Front. Psychiatry 2017, 8, 118. [Google Scholar] [CrossRef] [Green Version]
- Hoftman, G.D.; Volk, D.W.; Bazmi, H.H.; Li, S.; Sampson, A.R.; Lewis, D.A. Altered cortical expression of GABA-related genes in schizophrenia: Illness progression vs developmental disturbance. Schizophr. Bull. 2015, 41, 180–191. [Google Scholar] [CrossRef]
- Richetto, J.; Labouesse, M.; Poe, M.M.; Cook, J.M.; Grace, A.A.; Riva, M.A.; Meyer, U. Behavioral Effects of the Benzodiazepine-Positive Allosteric Modulator SH-053-2′F-S-CH3 in an Immune-Mediated Neurodevelopmental Disruption Model. Int. J. Neuropsychopharmacol. 2015, 18, pyu055. [Google Scholar] [CrossRef] [Green Version]
- Lazarus, M.S.; Krishnan, K.; Huang, Z.J. GAD67 deficiency in parvalbumin interneurons produces deficits in inhibitory transmission and network disinhibition in mouse prefrontal cortex. Cereb. Cortex 2015, 25, 1290–1296. [Google Scholar] [CrossRef] [Green Version]
- Luoni, A.; Richetto, J.; Longo, L.; Riva, M. Chronic lurasidone treatment normalizes GABAergic marker alterations in the dorsal hippocampus of mice exposed to prenatal immune activation. Eur. Neuropsychopharmacol. 2017, 27, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Grimm, J.W.; See, R.E. Unique activation of extracellular striato-pallidal neurotransmitters in rats following acute risperidone. Brain Res. 1998, 801, 182–189. [Google Scholar] [CrossRef]
- Xu, S.; Gullapalli, R.P.; Frost, D.O. Olanzapine antipsychotic treatment of adolescent rats causes long term changes in glutamate and GABA levels in the nucleus accumbens. Schizophr. Res. 2015, 161, 452–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, B.; Lian, J.; Huang, X.-F.; Deng, C. Aripiprazole Increases the PKA Signalling and Expression of the GABAA Receptor and CREB1 in the Nucleus Accumbens of Rats. J. Mol. Neurosci. 2016, 59, 36–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chadha, R.; Meador-Woodruff, J.H. Downregulated AKT-mTOR signaling pathway proteins in dorsolateral prefrontal cortex in Schizophrenia. Neuropsychopharmacology 2020, 45, 1059–1067. [Google Scholar] [CrossRef]
- Dummler, B.; Hemmings, B.A. Physiological roles of PKB/Akt isoforms in development and disease. Biochem. Soc. Trans. 2007, 35, 231–235. [Google Scholar] [CrossRef]
- Matsuda, S.; Ikeda, Y.; Murakami, M.; Nakagawa, Y.; Tsuji, A.; Kitagishi, Y. Roles of PI3K/AKT/GSK3 Pathway Involved in Psychiatric Illnesses. Diseases 2019, 7, 22. [Google Scholar] [CrossRef] [Green Version]
- Bitanihirwe, B.K.Y.; Weber, L.; Feldon, J.; Meyer, U. Cognitive impairment following prenatal immune challenge in mice correlates with prefrontal cortical AKT1 deficiency. Int. J. Neuropsychopharmacol. 2010, 13, 981–996. [Google Scholar] [CrossRef] [Green Version]
- Willi, R.; Harmeier, A.; Giovanoli, S.; Meyer, U. Altered GSK3β signaling in an infection-based mouse model of developmental neuropsychiatric disease. Neuropharmacology 2013, 73, 56–65. [Google Scholar] [CrossRef]
- Bitanihirwe, B.K.Y.; Peleg-Raibstein, D.; Mouttet, F.; Feldon, J.; Meyer, U. Late prenatal immune activation in mice leads to behavioral and neurochemical abnormalities relevant to the negative symptoms of schizophrenia. Neuropsychopharmacology 2010, 35, 2462–2478. [Google Scholar] [CrossRef] [Green Version]
- Baghel, M.S.; Singh, B.; Patro, N.; Khanna, V.K.; Patro, I.K.; Thakur, M.K. Poly (I:C) Exposure in Early Life Alters Methylation of DNA and Acetylation of Histone at Synaptic Plasticity Gene Promoter in Developing Rat Brain Leading to Memory Impairment. Ann. Neurosci. 2019, 26, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Liu, X.; Lian, J.; Deng, C. Epigenetic histone modulations of PPARγ and related pathways contribute to olanzapine-induced metabolic disorders. Pharmacol. Res. 2020, 155, 104703. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Jia, H.; Kast, R.J.; Thomas, E.A. Epigenetic changes at gene promoters in response to immune activation in utero. Brain Behav. Immun. 2013, 30, 168–175. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Lian, J.; Su, Y.; Deng, C. Effects of Risperidone and Prenatal Poly I:C Exposure on GABAA Receptors and AKT-GSK3β Pathway in the Ventral Tegmental Area of Female Juvenile Rats. Biomolecules 2022, 12, 732. https://doi.org/10.3390/biom12050732
Chen S, Lian J, Su Y, Deng C. Effects of Risperidone and Prenatal Poly I:C Exposure on GABAA Receptors and AKT-GSK3β Pathway in the Ventral Tegmental Area of Female Juvenile Rats. Biomolecules. 2022; 12(5):732. https://doi.org/10.3390/biom12050732
Chicago/Turabian StyleChen, Shiyan, Jiamei Lian, Yueqing Su, and Chao Deng. 2022. "Effects of Risperidone and Prenatal Poly I:C Exposure on GABAA Receptors and AKT-GSK3β Pathway in the Ventral Tegmental Area of Female Juvenile Rats" Biomolecules 12, no. 5: 732. https://doi.org/10.3390/biom12050732
APA StyleChen, S., Lian, J., Su, Y., & Deng, C. (2022). Effects of Risperidone and Prenatal Poly I:C Exposure on GABAA Receptors and AKT-GSK3β Pathway in the Ventral Tegmental Area of Female Juvenile Rats. Biomolecules, 12(5), 732. https://doi.org/10.3390/biom12050732