
NETosis and Nucleosome Biomarkers in Septic Shock and Critical COVID-19 Patients: An Observational Study

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Population and Clinical Outcome
2.2. Blood Sample Collection
2.3. Circulating Nucleosomes, Neutrophil Activation and Inflammatory Biomarkers
2.4. Statistical Analyses
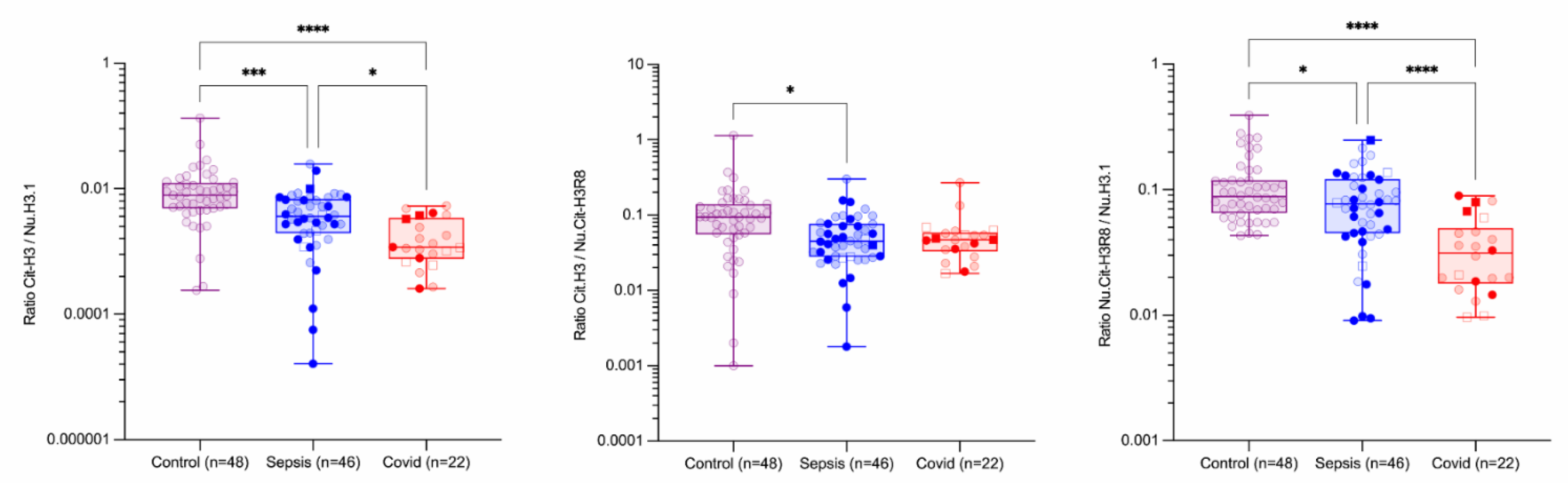
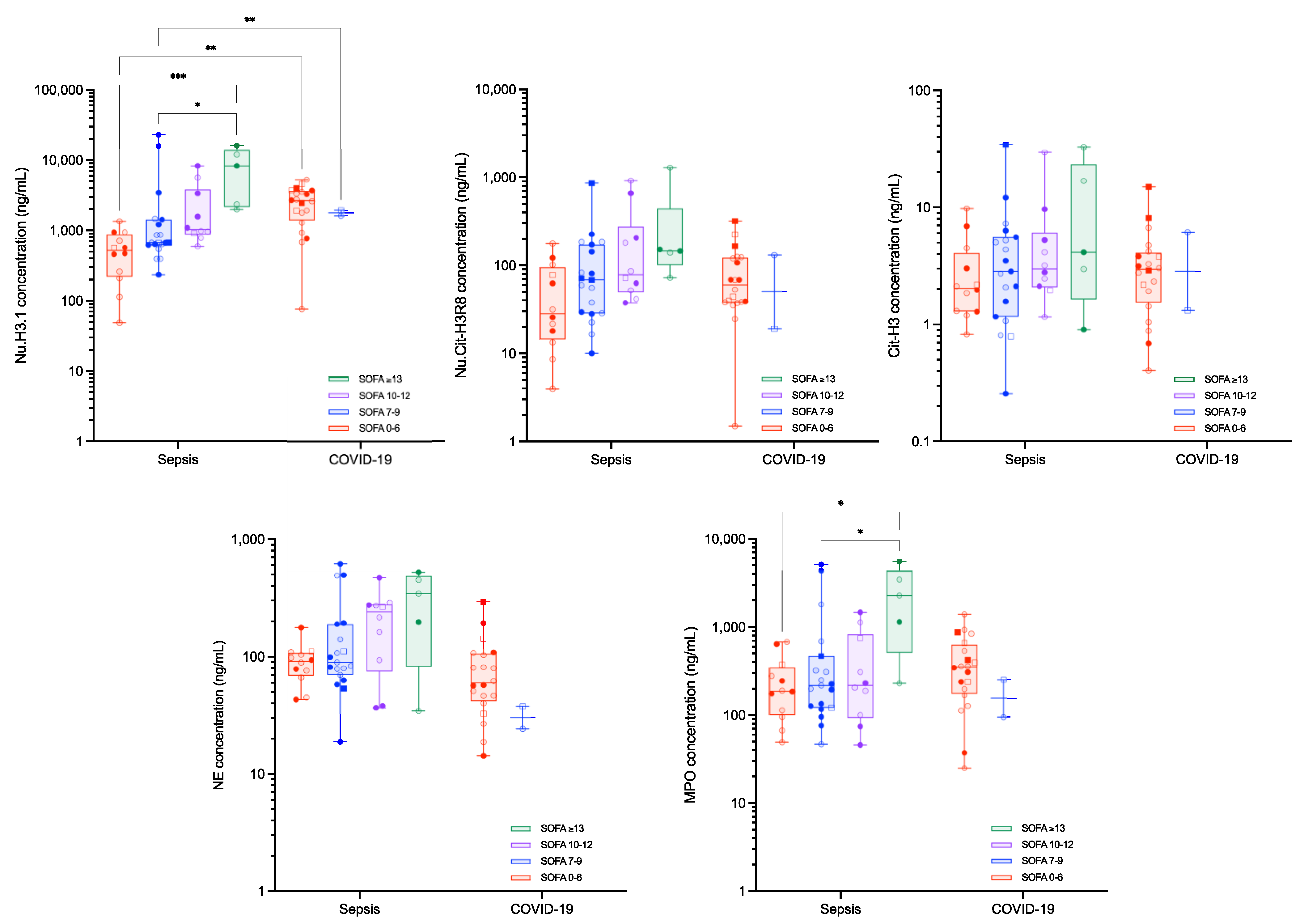
3. Results
Circulating Nucleosomes and Neutrophil Activation Biomarkers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References and Notes
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Urban, C.F.; Ermert, D.; Schmid, M.; Abu-Abed, U.; Goosmann, C.; Nacken, W.; Brinkmann, V.; Jungblut, P.R.; Zychlinsky, A. Neutrophil extracellular traps contain calprotectin, a cytosolic protein complex involved in host defense against Candida albicans. PLoS Pathog. 2009, 5, e1000639. [Google Scholar] [CrossRef] [Green Version]
- Petretto, A.; Bruschi, M.; Pratesi, F.; Croia, C.; Candiano, G.; Ghiggeri, G.; Migliorini, P. Neutrophil extracellular traps (NET) induced by different stimuli: A comparative proteomic analysis. PLoS ONE 2019, 14, e0218946. [Google Scholar] [CrossRef]
- Holdenrieder, S.; Stieber, P.; Bodenmuller, H.; Fertig, G.; Furst, H.; Schmeller, N.; Untch, M.; Seidel, D. Nucleosomes in serum as a marker for cell death. Clin. Chem. Lab. Med. 2001, 39, 596–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeerleder, S.; Zwart, B.; te Velthuis, H.; Stephan, F.; Manoe, R.; Rensink, I.; Aarden, L.A. Nucleosome-releasing factor: A new role for factor VII-activating protease (FSAP). FASEB J. 2008, 22, 4077–4084. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V.; Metzler, K.D.; Hakkim, A.; Zychlinsky, A. Neutrophil elastase and myeloperoxidase regulate the formation of neutrophil extracellular traps. J. Cell Biol. 2010, 191, 677–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wysocka, J.; Sayegh, J.; Lee, Y.H.; Perlin, J.R.; Leonelli, L.; Sonbuchner, L.S.; McDonald, C.H.; Cook, R.G.; Dou, Y.; et al. Human PAD4 regulates histone arginine methylation levels via demethylimination. Science 2004, 306, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Tanner, L.; Bhongir, R.K.V.; Karlsson, C.A.Q.; Le, S.; Ljungberg, J.K.; Andersson, P.; Andersson, C.; Malmstrom, J.; Egesten, A.; Single, A.B. Citrullination of extracellular histone H3.1 reduces antibacterial activity and exacerbates its proteolytic degradation. J. Cyst. Fibros. 2021, 20, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Yalavarthi, S.; Shi, H.; Gockman, K.; Zuo, M.; Madison, J.A.; Blair, C.; Weber, A.; Barnes, B.J.; Egeblad, M.; et al. Neutrophil extracellular traps in COVID-19. JCI Insight 2020, 5, e138999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, T.A.; Kremer Hovinga, J.A.; Schatzberg, D.; Wagner, D.D.; Lammle, B. Circulating DNA and myeloperoxidase indicate disease activity in patients with thrombotic microangiopathies. Blood 2012, 120, 1157–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalier, E.; Guiot, J.; Lechner, K.; Dutsch, A.; Eccleston, M.; Herzog, M.; Bygott, T.; Schomburg, A.; Kelly, T.; Holdenrieder, S. Circulating Nucleosomes as Potential Markers to Monitor COVID-19 Disease Progression. Front. Mol. Biosci 2021, 8, 600881. [Google Scholar] [CrossRef] [PubMed]
- Tsourouktsoglou, T.D.; Warnatsch, A.; Ioannou, M.; Hoving, D.; Wang, Q.; Papayannopoulos, V. Histones, DNA, and Citrullination Promote Neutrophil Extracellular Trap Inflammation by Regulating the Localization and Activation of TLR4. Cell Rep. 2020, 31, 107602. [Google Scholar] [CrossRef]
- Lin, G.L.; McGinley, J.P.; Drysdale, S.B.; Pollard, A.J. Epidemiology and Immune Pathogenesis of Viral Sepsis. Front. Immunol. 2018, 9, 2147. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Li, H.; Liu, L.; Zhang, D.; Xu, J.; Dai, H.; Tang, N.; Su, X.; Cao, B. SARS-CoV-2 and viral sepsis: Observations and hypotheses. Lancet 2020, 395, 1517–1520. [Google Scholar] [CrossRef]
- Lin, L.; Lu, L.; Cao, W.; Li, T. Hypothesis for potential pathogenesis of SARS-CoV-2 infection-a review of immune changes in patients with viral pneumonia. Emerg. Microbes Infect. 2020, 9, 727–732. [Google Scholar] [CrossRef] [Green Version]
- Dechamps, M.; De Poortere, J.; Martin, M.; Gatto, L.; Daumerie, A.; Bouzin, C.; Octave, M.; Ginion, A.; Robaux, V.; Pirotton, L.; et al. Inflammation-induced coagulopathy substantially differs between COVID-19 and septic shock: A prospective observational study. Front. Med. 2022, 8, 780750. [Google Scholar] [CrossRef]
- Gillot, C.; Favresse, J.; Mullier, F.; Lecompte, T.; Dogne, J.M.; Douxfils, J. NETosis and the Immune System in COVID-19: Mechanisms and Potential Treatments. Front. Pharmacol. 2021, 12, 708302. [Google Scholar] [CrossRef]
- Ma, A.C.; Kubes, P. Platelets, neutrophils, and neutrophil extracellular traps (NETs) in sepsis. J. Thromb. Haemost. 2008, 6, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Alhazzani, W.; Evans, L.; Alshamsi, F.; Moller, M.H.; Ostermann, M.; Prescott, H.C.; Arabi, Y.M.; Loeb, M.; Ng Gong, M.; Fan, E.; et al. Surviving Sepsis Campaign Guidelines on the Management of Adults With Coronavirus Disease 2019 (COVID-19) in the ICU: First Update. Crit. Care Med. 2021, 49, e219–e234. [Google Scholar] [CrossRef] [PubMed]
- Force, A.D.T.; Ranieri, V.M.; Rubenfeld, G.D.; Thompson, B.T.; Ferguson, N.D.; Caldwell, E.; Fan, E.; Camporota, L.; Slutsky, A.S. Acute respiratory distress syndrome: The Berlin Definition. JAMA 2012, 307, 2526–2533. [Google Scholar] [CrossRef]
- Jones, A.E.; Trzeciak, S.; Kline, J.A. The Sequential Organ Failure Assessment score for predicting outcome in patients with severe sepsis and evidence of hypoperfusion at the time of emergency department presentation. Crit. Care Med. 2009, 37, 1649–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iba, T.; Nisio, M.D.; Levy, J.H.; Kitamura, N.; Thachil, J. New criteria for sepsis-induced coagulopathy (SIC) following the revised sepsis definition: A retrospective analysis of a nationwide survey. BMJ Open 2017, 7, e017046. [Google Scholar] [CrossRef] [Green Version]
- Nu.Q H3.1 ELISA Assay—IFU Nu.QH3.1-01R—Version 3.
- Rothman, K.J. No adjustments are needed for multiple comparisons. Epidemiology 1990, 1, 43–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Zou, Z.Y.; Chen, Y.H.; Wang, C.L.; Feng, Y.W.; Liu, Z.F. Severe COVID-19-associated sepsis is different from classical sepsis induced by pulmonary infection with carbapenem-resistant klebsiella pneumonia (CrKP). Chin. J. Traumatol. 2022, 25, 17–24. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, X.; Ding, D.; Zhang, J.; Xu, L.; Hu, Z.; Xu, W.; Tao, Z. Comparative Study of Acute Lung Injury in COVID-19 and Non-COVID-19 Patients. Front. Med. 2021, 8, 666629. [Google Scholar] [CrossRef]
- Masso-Silva, J.A.; Moshensky, A.; Lam, M.T.Y.; Odish, M.; Patel, A.; Xu, L.; Hansen, E.; Trescott, S.; Nguyen, C.; Kim, R.; et al. Increased peripheral blood neutrophil activation phenotypes and NETosis in critically ill COVID-19 patients: A case series and review of the literature. Clin. Infect. Dis. 2021, 74, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Chow, O.A.; von Kockritz-Blickwede, M.; Bright, A.T.; Hensler, M.E.; Zinkernagel, A.S.; Cogen, A.L.; Gallo, R.L.; Monestier, M.; Wang, Y.; Glass, C.K.; et al. Statins enhance formation of phagocyte extracellular traps. Cell Host Microbe 2010, 8, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Von Kockritz-Blickwede, M.; Goldmann, O.; Thulin, P.; Heinemann, K.; Norrby-Teglund, A.; Rohde, M.; Medina, E. Phagocytosis-independent antimicrobial activity of mast cells by means of extracellular trap formation. Blood 2008, 111, 3070–3080. [Google Scholar] [CrossRef]
- Theoharides, T.C. Potential association of mast cells with coronavirus disease 2019. Ann. Allergy Asthma Immunol. 2021, 126, 217–218. [Google Scholar] [CrossRef] [PubMed]
- Knoll, R.; Schultze, J.L.; Schulte-Schrepping, J. Monocytes and Macrophages in COVID-19. Front. Immunol. 2021, 12, 720109. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; An, L.L.; Chaerkady, R.; Mittereder, N.; Clarke, L.; Cohen, T.S.; Chen, B.; Hess, S.; Sims, G.P.; Mustelin, T. Evidence for a direct link between PAD4-mediated citrullination and the oxidative burst in human neutrophils. Sci. Rep. 2018, 8, 15228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middleton, E.A.; He, X.Y.; Denorme, F.; Campbell, R.A.; Ng, D.; Salvatore, S.P.; Mostyka, M.; Baxter-Stoltzfus, A.; Borczuk, A.C.; Loda, M.; et al. Neutrophil extracellular traps contribute to immunothrombosis in COVID-19 acute respiratory distress syndrome. Blood 2020, 136, 1169–1179. [Google Scholar] [CrossRef]
- Fuentes Baldarrago, A.L.; Patel, A.; Masso Silva, J.A.; Moshensky, A.; Perera, S.; Ma, L.J.; Pham, J.; Lam, M.T.Y.; Odish, M.F.; Coufal, N.; et al. Neutrophil Extracellular Trap Formation (NETosis) Increases with Severity of Disease in COVID-19 Patients. In Proceedings of the American Thoracic Society 2021 International Conference, San Diego, CA, USA, 14–19 May 2021. [Google Scholar]
- Castanheira, F.V.S.; Kubes, P. Neutrophils and NETs in modulating acute and chronic inflammation. Blood 2019, 133, 2178–2185. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Ye, L.; Jin, Y.; Zhang, N.; Lou, T.; Qiu, Z.; Jin, Y.; Cheng, B.; Fang, X. Circulating nucleosomes as a predictor of sepsis and organ dysfunction in critically ill patients. Int. J. Infect. Dis. 2012, 16, e558–e564. [Google Scholar] [CrossRef] [Green Version]
- Shabrish, S.; Mittra, I. Cytokine Storm as a Cellular Response to dsDNA Breaks: A New Proposal. Front. Immunol. 2021, 12, 622738. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nu.H3.1 (ng/mL) | Nu.H3R8 (ng/mL) | Cit.H3 (ng/mL) | NE (ng/mL) | MPO (ng/mL) | ||
|---|---|---|---|---|---|---|
| All cohort | ||||||
| Survivors (n = 42) Median (10th–90th percentile) | 1081.4 [222.3–4295.4] | 54.3 [14.3–184.9] | 2.6 [0.8–8.9] | 86.7 [33.3–285.1] | 251.8 [74.5–1312.2] | |
| Non-survivors (n = 26) Median (10th–90th percentile) | 1778.3 [439.5–15,848.9] | 76.0 [23.1–396.3] | 3.1 [0.8–12.9] | 96.6 [30.1–504.7] | 235.0 [64.1–4581.4] | |
| p-value | 0.0982 | 0.1178 | 0.4818 | 0.3573 | 0.7197 | |
| Septic shock | ||||||
| Alive (n = 26) Median (10th–90th percentile) | 785.2 [173.4–3076.1] | 65.5 [11.7–299.2] | 2.6 [0.8–20.0] | 106.2 [59.4–374.1] | 239.9 [61.1–1932.0] | |
| Death (n = 20) Median (10th–90th percentile) | 901.6 [402.7–16,032.5] | 76.0 [18.7–592.9] | 2.9 [0.9–11.8] | 96.6 [37.0–523.6] | 210.4 [74.3–5046.6] | |
| p-value | 0.0664 | 0.4007 | 0.9598 | 0.8881 | 0.6946 | |
| COVID-19 | ||||||
| Alive (n = 16) Median (10th–90th percentile) | 1927.5 [353.2–4764.3] | 41.9 [8.9–154.2] | 2.5 [0.7–6.3] | 48.9 [22.5–117.5] | 301.3 [63.8–1052.0] | |
| Death (n = 6) Median (10th–90th percentile) | 2964.8 [765.6–3999.4] | 85.9 [39.1–318.4] | 3.5 [0.7–15.0] | 79.3 [14.3–292.4] | 326.6 [37.3–875.0] | |
| p-value | 0.6946 | 0.1442 | 0.2272 | 0.2349 | 0.9187 | |
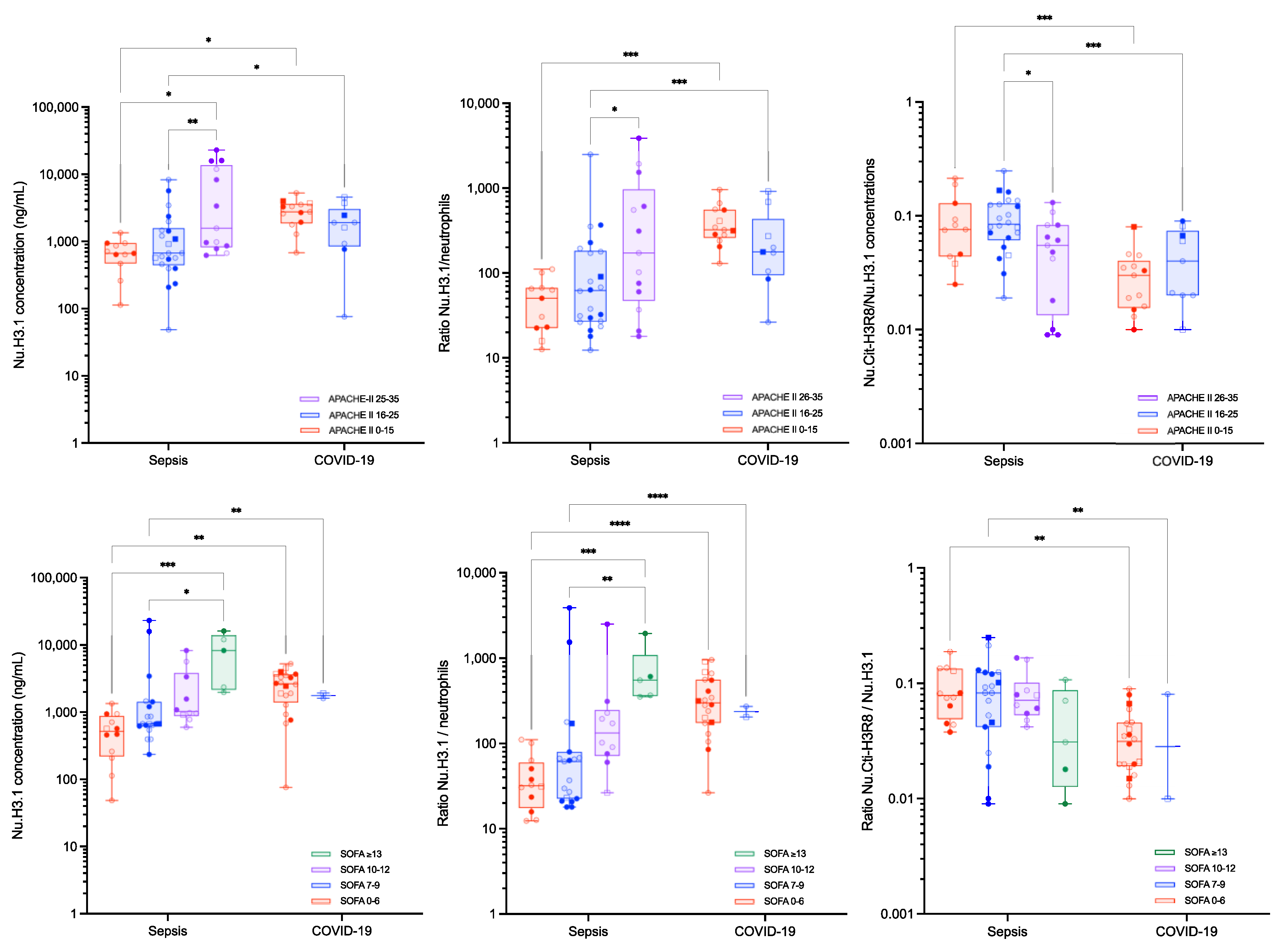
| APACHE-II 0–15 | APACHE-II 16–25 | APACHE-II 25–35 | SOFA 0–6 | SOFA 7–9 | SOFA 10–12 | SOFA ≥ 13 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nu.H3.1 (ng/mL) | ||||||||||||
| Septic shock | 666.4 (133.7–1257.8) | 670.0 (215.9–4898.9) | 1575.3 (641.4–19,955.7) | 517.9 (62.6–1213.9) | 673.0 (396.9–15,775.5) | 1032.8 (612.7–7993.8) | 8285.6 (1980.4–16,068.7) | |||||
| Critical COVID-19 | 2764.5 (877.8–4720.9) | 1904.0 (76.0–4555.3) | 2648.3 (689.2–4496.4) | 1769.1 (1615.1–1937.8) | ||||||||
| corrected p-value | 0.0321 | 0.0321 | 0.0025 | 0.025 | ||||||||
| Nu.H3.1/neutrophils | ||||||||||||
| Septic shock | 50.5 (13.2–108.9) | 62.3 (18.8–362.7) | 172.4 (19.0–2934.4) | 31.8 (12.4–107.9) | 61.4 (18.0–1533.1) | 132.7 (28.7–2025.4) | 553.7 (353.0–1933.7) | |||||
| Critical COVID-19 | 322.6 (155.5–826.3) | 177.4 (26.4–914.7) | 299.5 (86.9–888.9) | 236.0 (204.4–272.4) | ||||||||
| corrected p-value | 0.0005 | 0.0005 | <0.0001 | <0.0001 | ||||||||
| Nu.Cit-H3R8 (ng/mL) | ||||||||||||
| Septic shock | 31.6 (10.2–183.6) | 70.2 (11.0–798.3) | 86.2 (39.3–862.1) | 28.6 (5.4–161.1) | 68.5 (16.5–226.3) | 79.1 (38.0–893.3) | 145.7 (72.3–1286.0) | |||||
| Critical COVID-19 | 68.4 (21.9–271.5) | 41.1 (1.5–165.9) | 60.6 (25.6–218.6) | 75.3 (19.2–131.4) | ||||||||
| corrected p-value | >0.9999 | >0.9999 | 0.9538 | 0.9538 | ||||||||
| Cit-H3 (ng/mL) | ||||||||||||
| Septic shock | 2.12 (0.79–9.28) | 2.58 (1.1–25.8) | 3.18 (0.52–24.5) | 2.04 (0.94–8.92) | 2.84 (0.79–12.1) | 2.99 (1.24–27.6) | 4.14 (0.91–32.7) | |||||
| Critical COVID-19 | 3.04 (0.94–11.7) | 2.77 (0.40–2.77) | 2.96 (0.71–8.01) | 3.74 (1.32–6.15) | ||||||||
| corrected p-value | 0.9966 | 0.9966 | >0.9999 | >0.9999 | ||||||||
| Nu.Cit-H3R8/Nu.H3.1 | ||||||||||||
| Septic shock | 0.076 (0.027–0.209) | 0.084 (0.034–0.165) | 0.055 (0.009–0.121) | 0.079 (0.040–0.172) | 0.083 (0.010–0.214) | 0.072 (0.043–0.166) | 0.031 (0.009–0.108) | |||||
| Critical COVID-19 | 0.030 (0.011–0.064) | 0.040 (0.010–0.090) | 0.031 (0.013–0.079) | 0.028 (0.010–0.081) | ||||||||
| corrected p-value | 0.0002 | 0.0002 | 0.0038 | 0.038 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morimont, L.; Dechamps, M.; David, C.; Bouvy, C.; Gillot, C.; Haguet, H.; Favresse, J.; Ronvaux, L.; Candiracci, J.; Herzog, M.; et al. NETosis and Nucleosome Biomarkers in Septic Shock and Critical COVID-19 Patients: An Observational Study. Biomolecules 2022, 12, 1038. https://doi.org/10.3390/biom12081038
Morimont L, Dechamps M, David C, Bouvy C, Gillot C, Haguet H, Favresse J, Ronvaux L, Candiracci J, Herzog M, et al. NETosis and Nucleosome Biomarkers in Septic Shock and Critical COVID-19 Patients: An Observational Study. Biomolecules. 2022; 12(8):1038. https://doi.org/10.3390/biom12081038
Chicago/Turabian StyleMorimont, Laure, Mélanie Dechamps, Clara David, Céline Bouvy, Constant Gillot, Hélène Haguet, Julien Favresse, Lorian Ronvaux, Julie Candiracci, Marielle Herzog, and et al. 2022. "NETosis and Nucleosome Biomarkers in Septic Shock and Critical COVID-19 Patients: An Observational Study" Biomolecules 12, no. 8: 1038. https://doi.org/10.3390/biom12081038
APA StyleMorimont, L., Dechamps, M., David, C., Bouvy, C., Gillot, C., Haguet, H., Favresse, J., Ronvaux, L., Candiracci, J., Herzog, M., Laterre, P. -F., De Poortere, J., Horman, S., Beauloye, C., & Douxfils, J. (2022). NETosis and Nucleosome Biomarkers in Septic Shock and Critical COVID-19 Patients: An Observational Study. Biomolecules, 12(8), 1038. https://doi.org/10.3390/biom12081038