
Ginsenoside and Its Therapeutic Potential for Cognitive Impairment


Abstract
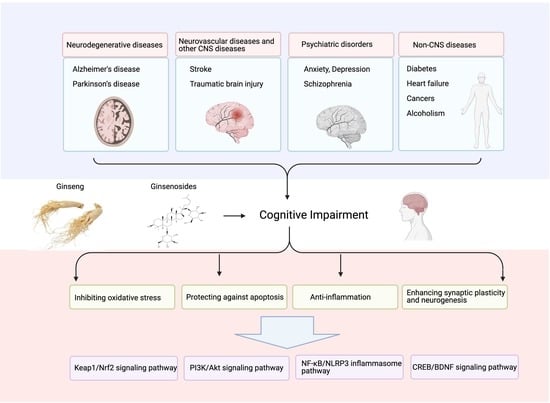
:
1. Introduction
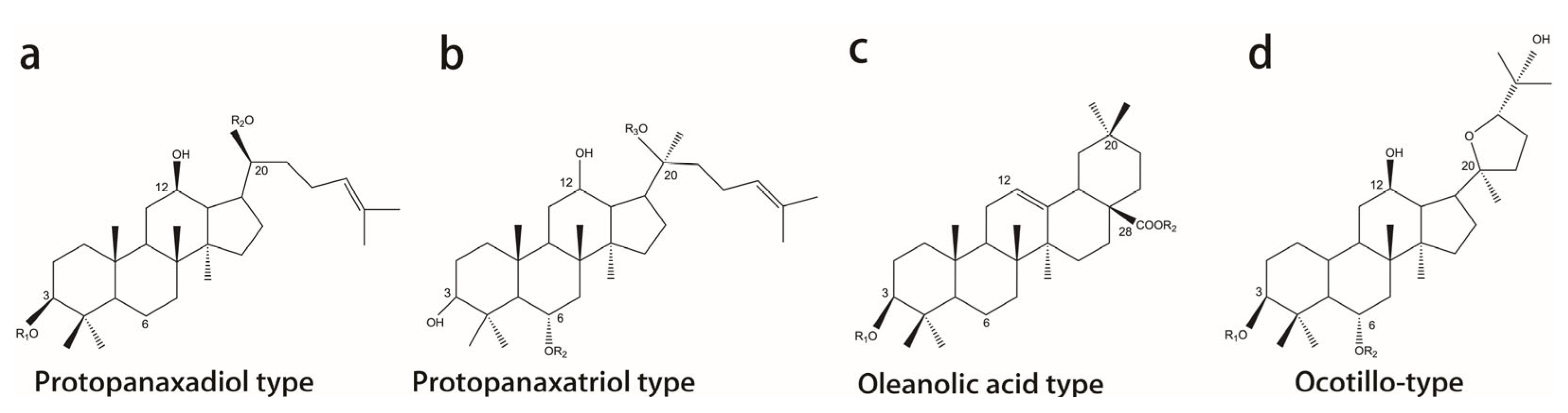
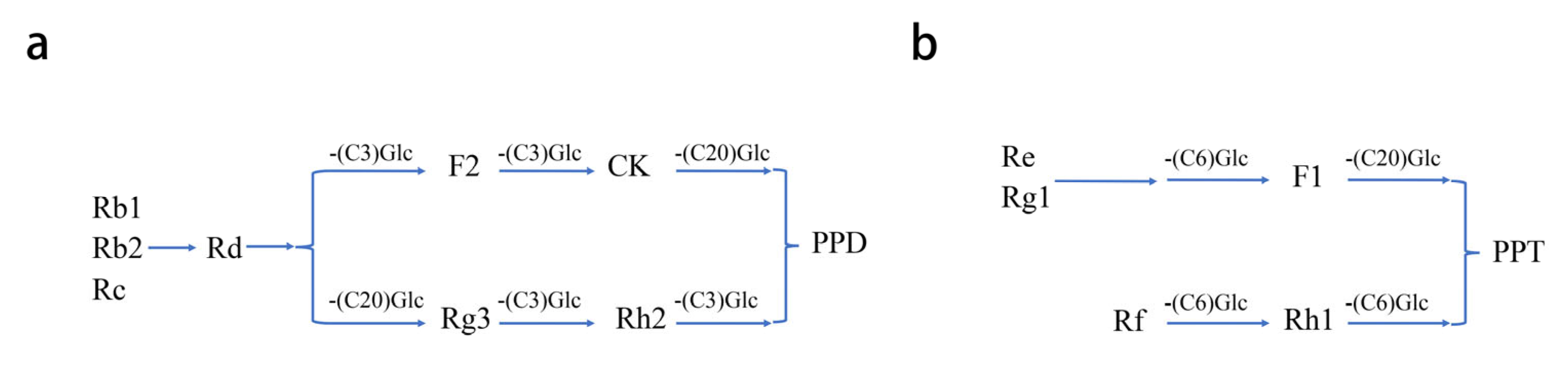
2. Classification, Chemical Structure, and Pharmacokinetics of Ginsenosides
3. Protective Effect of Ginsenosides on CI
3.1. Neurodegenerative Diseases
3.2. Neurovascular Diseases and Other CNS Diseases
3.3. Psychiatric Disorders
3.4. Non-CNS Diseases
4. Pharmacological Properties of Ginsenosides of CI
4.1. Regulating Cholinergic Transmission
4.2. Inhibiting Oxidative Stress
4.3. Protecting against Apoptosis
4.4. Inhibiting Inflammation
4.5. Enhancing Synaptic Plasticity and Neurogenesis
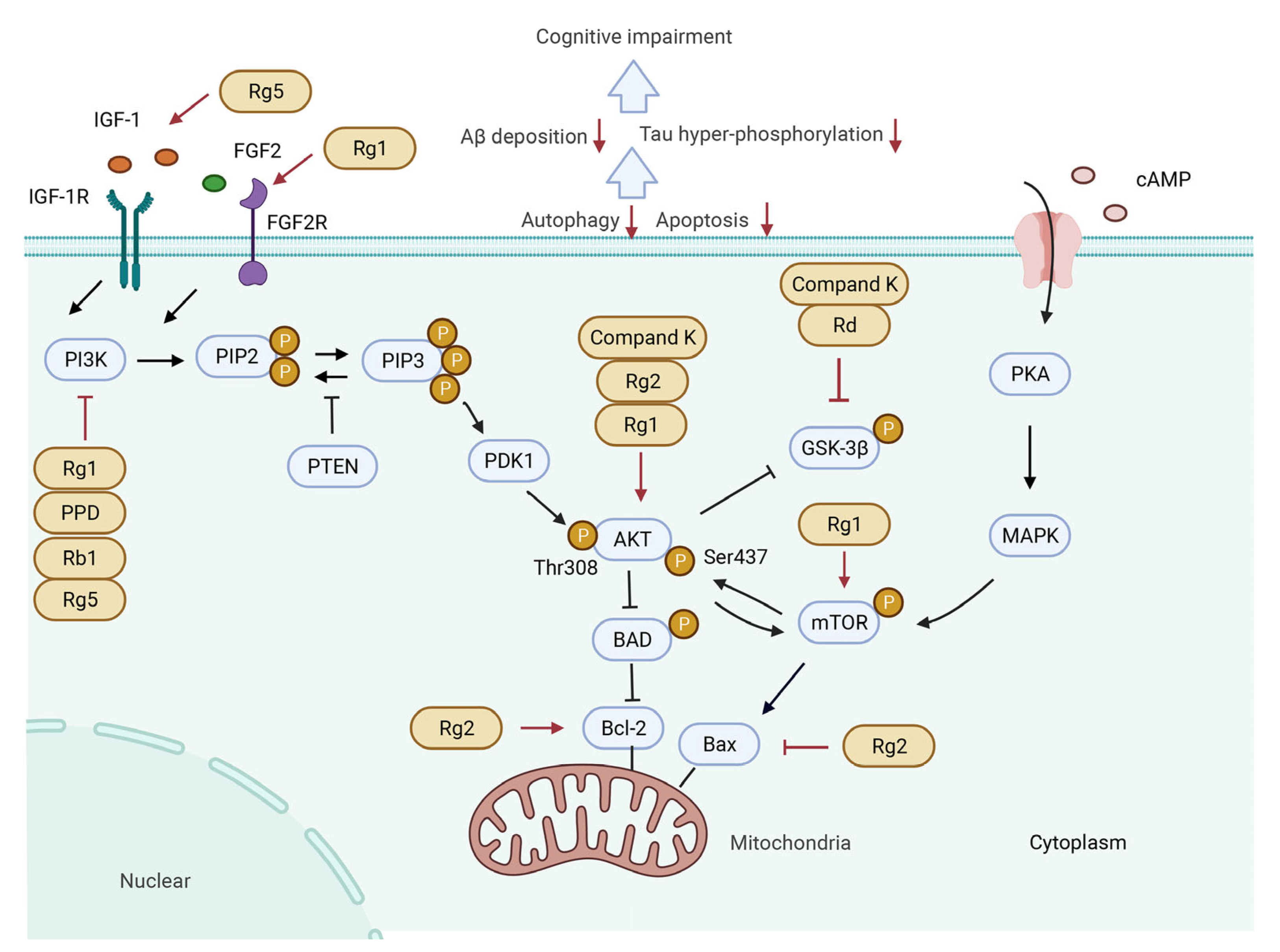
5. Signaling Pathways Involved in the Treatment of CI by Ginsenosides
5.1. PI3K/Akt Signaling Pathway
5.1.1. Activating PI3K/Akt Signaling Pathway by Upstream Cytokines
5.1.2. Regulating PI3K/Akt Signaling Pathway and Downstream Molecules
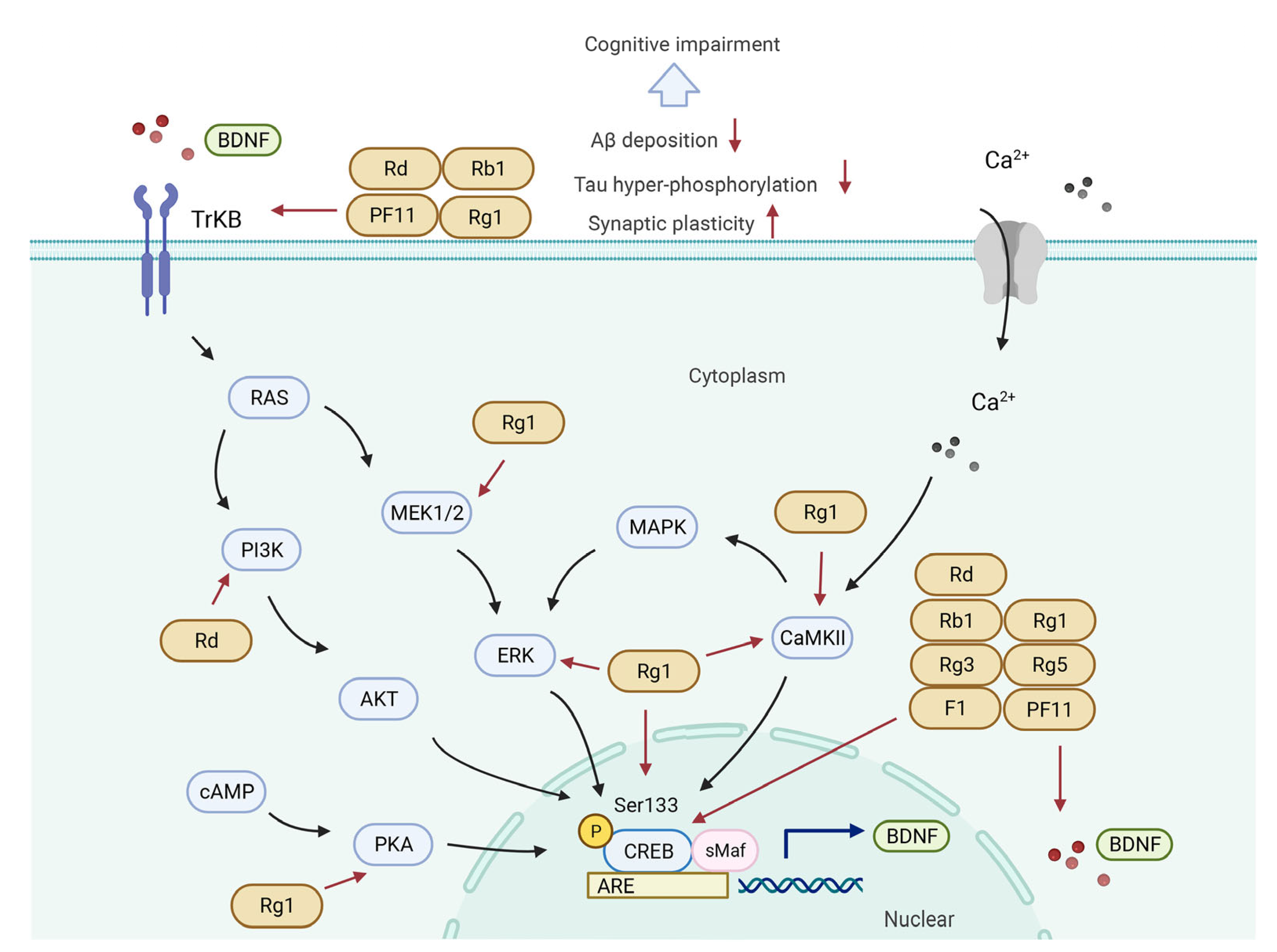
5.2. CREB/BDNF Signaling Pathway
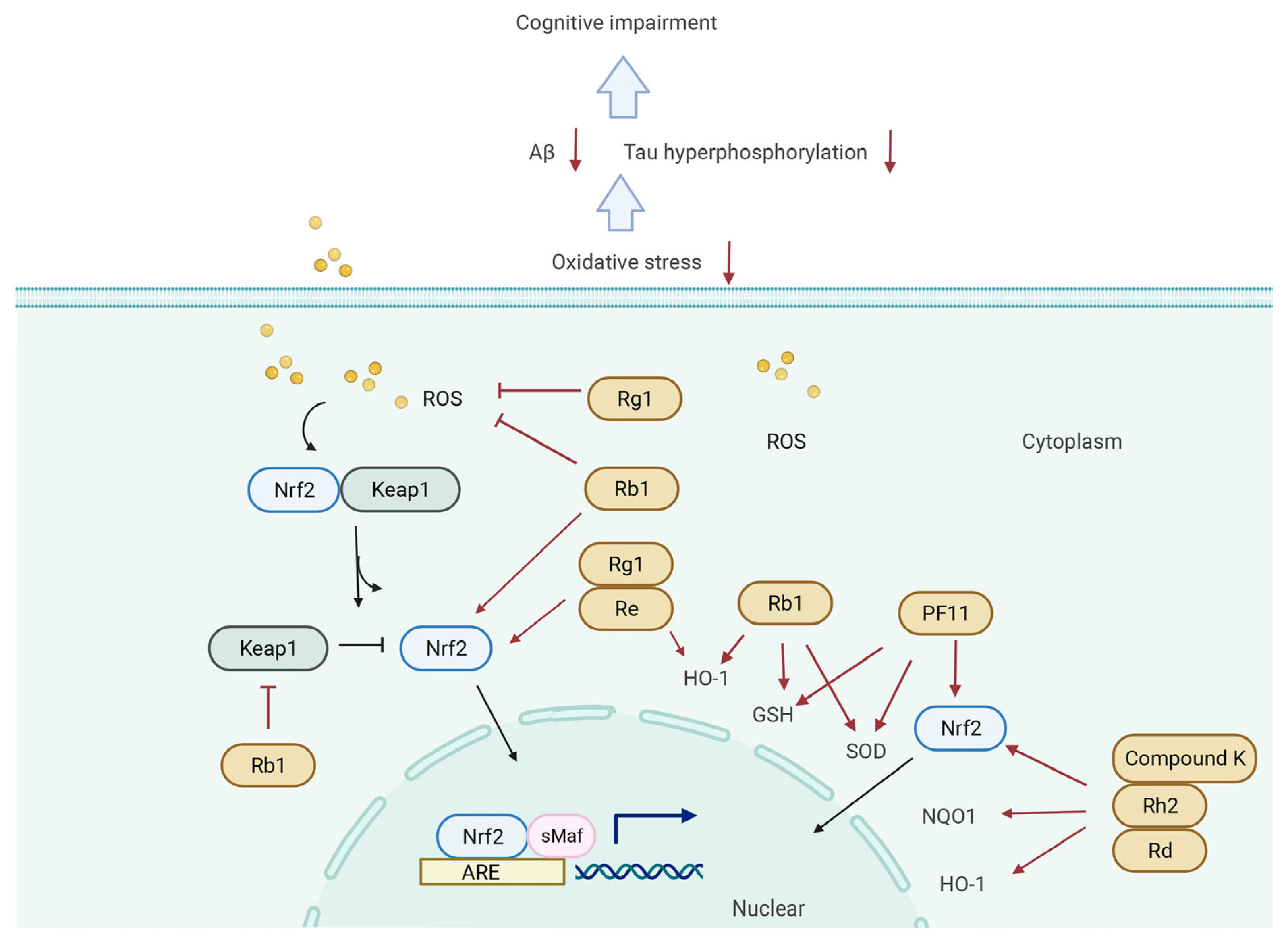
5.3. Keap1/Nrf2 Signaling Pathway
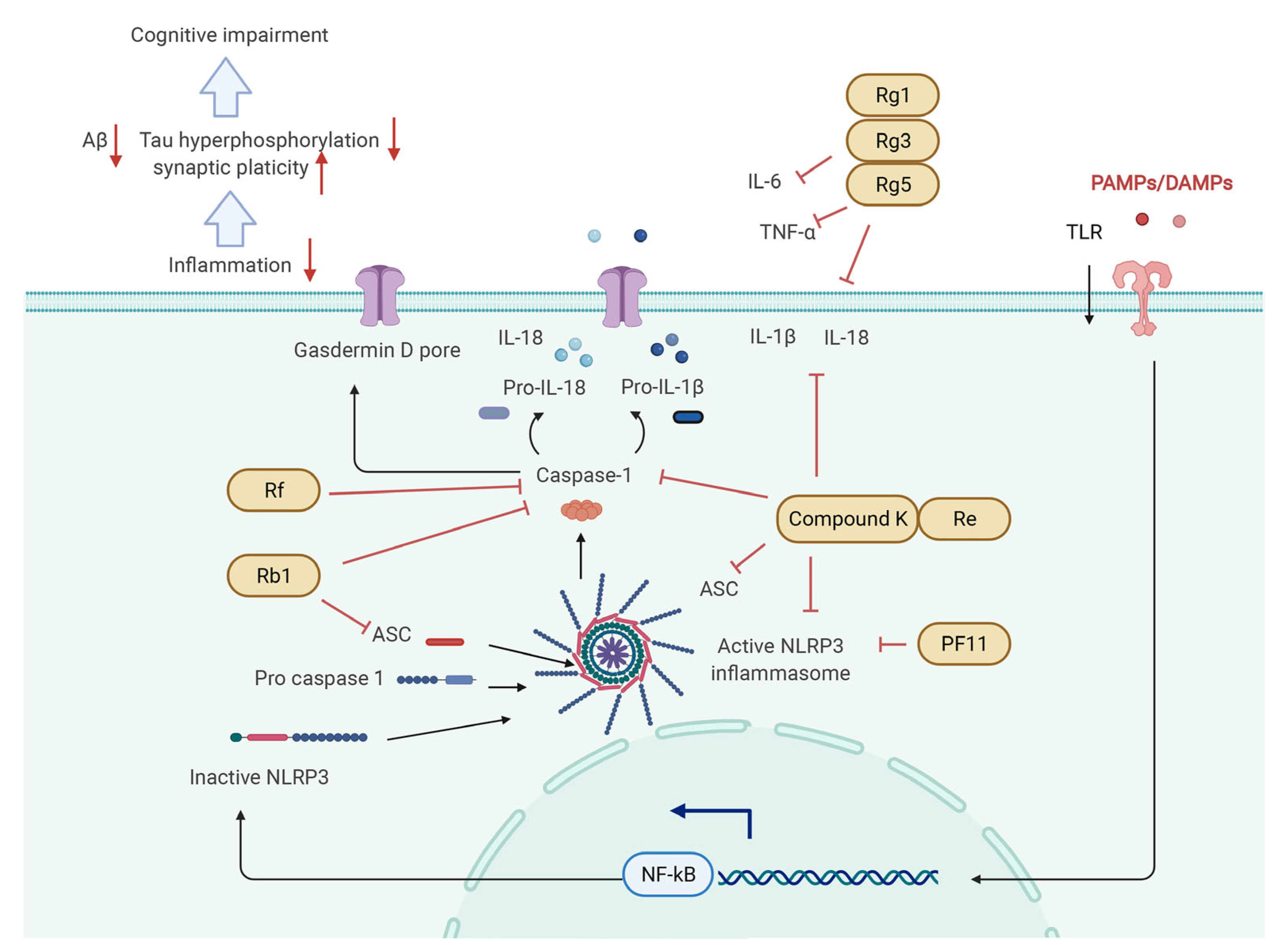
5.4. NF-κB/NLRP3 Inflammasome Pathway
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, N.D. State of the science on mild cognitive impairment (MCI). CNS Spectr. 2019, 24, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.P.; Feng, L.; Nyunt, M.S.; Feng, L.; Gao, Q.; Lim, M.L.; Collinson, S.L.; Chong, M.S.; Lim, W.S.; Lee, T.S.; et al. Metabolic Syndrome and the Risk of Mild Cognitive Impairment and Progression to Dementia: Follow-up of the Singapore Longitudinal Ageing Study Cohort. JAMA Neurol. 2016, 73, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Prince, M.; Comas-Herrera, A.; Knapp, M.; Guerchet, G.; Karagiannidou, M. World Alzheimer Report 2016: Improving Healthcare for People Living with Dementia: Coverage, Quality and Costs Now and in the Future 2016; Alzheimer’s Disease International (ADI): London, UK, 2018. [Google Scholar]
- Duan, J.; Lv, Y.B.; Gao, X.; Zhou, J.H.; Kraus, V.B.; Zeng, Y.; Hong, S.; Shi, X.M. Association of cognitive impairment and elderly mortality: Differences between two cohorts ascertained 6-years apart in China. BMC Geriatr. 2020, 20, 29. [Google Scholar] [CrossRef] [PubMed]
- Tangalos, E.G.; Petersen, R.C. Mild Cognitive Impairment in Geriatrics. Clin. Geriatr. Med. 2018, 34, 563–589. [Google Scholar] [CrossRef]
- Arvanitakis, Z.; Shah, R.C.; Bennett, D.A. Diagnosis and Management of Dementia: Review. Jama 2019, 322, 1589–1599. [Google Scholar] [CrossRef]
- Blanco-Silvente, L.; Castells, X.; Saez, M.; Barceló, M.A.; Garre-Olmo, J.; Vilalta-Franch, J.; Capella, D. Discontinuation, Efficacy, and Safety of Cholinesterase Inhibitors for Alzheime’s Disease: A Meta-Analysis and Meta-Regression of 43 Randomized Clinical Trials Enrolling 16 106 Patients. Int. J. Neuropsychopharmacol. 2017, 20, 519–528. [Google Scholar] [CrossRef]
- Campbell, N.L.; Perkins, A.J.; Gao, S.; Skaar, T.C.; Li, L.; Hendrie, H.C.; Fowler, N.; Callahan, M.C.; Boustani, M.A. Adherence and Tolerability of Alzheimer’s Disease Medications: A Pragmatic Randomized Trial. J. Am. Geriatr. Soc. 2017, 65, 1497–1504. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Huang, Y.; Yin, G.; Wang, J.; Wang, P.; Chen, Z.Y.; Wang, T.J.; Ren, G.X. Antimicrobial activities of Asian ginseng, American ginseng, and notoginseng. Phytother. Res. PTR 2020, 34, 1226–1236. [Google Scholar] [CrossRef]
- Jin, Y.; Cui, R.; Zhao, L.; Fan, J.; Li, B. Mechanisms of Panax ginseng action as an antidepressant. Cell Prolif. 2019, 52, e12696. [Google Scholar] [CrossRef]
- Heo, J.H.; Lee, S.T.; Chu, K.; Oh, M.J.; Park, H.J.; Shim, J.Y.; Kim, M. Heat-processed ginseng enhances the cognitive function in patients with moderately severe Alzheimer’s disease. Nutr. Neurosci. 2012, 15, 278–282. [Google Scholar] [CrossRef]
- Liu, M.; Yu, S.; Wang, J.; Qiao, J.; Liu, Y.; Wang, S.; Zhao, Y. Ginseng protein protects against mitochondrial dysfunction and neurodegeneration by inducing mitochondrial unfolded protein response in Drosophila melanogaster PINK1 model of Parkinson’s disease. J. Ethnopharmacol. 2020, 247, 112213. [Google Scholar] [CrossRef] [PubMed]
- Sood, A.; Mehrotra, A.; Dhawan, D.K.; Sandhir, R. Indian Ginseng (Withania somnifera) supplementation ameliorates oxidative stress and mitochondrial dysfunctions in experimental model of stroke. Metab. Brain Dis. 2018, 33, 1261–1274. [Google Scholar] [CrossRef] [PubMed]
- Christensen, L.P. Ginsenosides chemistry, biosynthesis, analysis, and potential health effects. Adv. Food Nutr. Res. 2009, 55, 1–99. [Google Scholar] [PubMed]
- Huang, X.; Li, N.; Pu, Y.; Zhang, T.; Wang, B. Neuroprotective Effects of Ginseng Phytochemicals: Recent Perspectives. Molecules 2019, 24, 2939. [Google Scholar] [CrossRef]
- Wong, A.S.; Che, C.M.; Leung, K.W. Recent advances in ginseng as cancer therapeutics: A functional and mechanistic overview. Nat. Prod. Rep. 2015, 32, 256–272. [Google Scholar] [CrossRef]
- Pan, W.; Xue, B.; Yang, C.; Miao, L.; Zhou, L.; Chen, Q.; Cai, Q.; Liu, Y.; Liu, D.; He, H.; et al. Biopharmaceutical characters and bioavailability improving strategies of ginsenosides. Fitoterapia 2018, 129, 272–282. [Google Scholar] [CrossRef]
- Piao, X.; Zhang, H.; Kang, J.P.; Yang, D.U.; Li, Y.; Pang, S.; Jin, Y.; Yang, D.C.; Wang, Y. Advances in Saponin Diversity of Panax ginseng. Molecules 2020, 25, 3452. [Google Scholar] [CrossRef]
- Shi, Z.Y.; Zeng, J.Z.; Wong, A.S.T. Chemical Structures and Pharmacological Profiles of Ginseng Saponins. Molecules 2019, 24, 2443. [Google Scholar] [CrossRef]
- Yang, W.Z.; Hu, Y.; Wu, W.Y.; Ye, M.; Guo, D.A. Saponins in the genus Panax L. (Araliaceae): A systematic review of their chemical diversity. Phytochemistry 2014, 106, 7–24. [Google Scholar] [CrossRef]
- Wang, Z.J.; Sun, L.; Peng, W.; Ma, S.; Zhu, C.; Fu, F.; Heinbockel, T. Ginseng derivative ocotillol enhances neuronal activity through increased glutamate release: A possible mechanism underlying increased spontaneous locomotor activity of mice. Neuroscience 2011, 195, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Luo, B.Y.; Jiang, J.L.; Fang, Y.F.; Yang, F.; Yin, M.D.; Zhang, B.C.; Zhao, R.R.; Shao, J.W. The effects of ginsenosides on platelet aggregation and vascular intima in the treatment of cardiovascular diseases: From molecular mechanisms to clinical applications. Pharmacol. Res. 2020, 159, 105031. [Google Scholar] [CrossRef] [PubMed]
- Mohanan, P.; Subramaniyam, S.; Mathiyalagan, R.; Yang, D.C. Molecular signaling of ginsenosides Rb1, Rg1, and Rg3 and their mode of actions. J. Ginseng Res. 2018, 42, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.N.; Ha, Y.W.; Shin, H.; Son, S.H.; Wu, S.J.; Kim, Y.S. Simultaneous quantification of 14 ginsenosides in Panax ginseng C.A. Meyer (Korean red ginseng) by HPLC-ELSD and its application to quality control. J. Pharm. Biomed. Anal. 2007, 45, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Balan, P.; Popovich, D.G. Comparison of Ginsenoside Components of Various Tissues of New Zealand Forest-Grown Asian Ginseng (Panax Ginseng) and American Ginseng (Panax Quinquefolium L.). Biomolecules 2020, 10, 372. [Google Scholar] [CrossRef]
- Qi, L.W.; Wang, C.Z.; Yuan, C.S. Ginsenosides from American ginseng: Chemical and pharmacological diversity. Phytochemistry 2011, 72, 689–699. [Google Scholar] [CrossRef]
- Chuang, W.C.; Wu, H.K.; Sheu, S.J.; Chiou, S.H.; Chang, H.C.; Chen, Y.P. A comparative study on commercial samples of ginseng radix. Planta Med. 1995, 61, 459–465. [Google Scholar] [CrossRef]
- Liu, H.; Lu, X.; Hu, Y.; Fan, X. Chemical constituents of Panax ginseng and Panax notoginseng explain why they differ in therapeutic efficacy. Pharmacol. Res. 2020, 161, 105263. [Google Scholar] [CrossRef]
- Liu, H.; Yang, J.; Du, F.; Gao, X.; Ma, X.; Huang, Y.; Xu, F.; Niu, W.; Wang, F.; Mao, Y.; et al. Absorption and disposition of ginsenosides after oral administration of Panax notoginseng extract to rats. Drug Metab. Dispos. Biol. Fate Chem. 2009, 37, 2290–2298. [Google Scholar] [CrossRef]
- Wang, H.Y.; Qi, L.W.; Wang, C.Z.; Li, P. Bioactivity enhancement of herbal supplements by intestinal microbiota focusing on ginsenosides. Am. J. Chin. Med. 2011, 39, 1103–1115. [Google Scholar] [CrossRef]
- Yang, L.; Zou, H.; Gao, Y.; Luo, J.; Xie, X.; Meng, W.; Zhou, H.; Tan, Z. Insights into gastrointestinal microbiota-generated ginsenoside metabolites and their bioactivities. Drug Metab. Rev. 2020, 52, 125–138. [Google Scholar] [CrossRef]
- Jin, S.; Jeon, J.H.; Lee, S.; Kang, W.Y.; Seong, S.J.; Yoon, Y.R.; Choi, M.K.; Song, I.S. Detection of 13 Ginsenosides (Rb1, Rb2, Rc, Rd, Re, Rf, Rg1, Rg3, Rh2, F1, Compound K, 20(S)-Protopanaxadiol, and 20(S)-Protopanaxatriol) in Human Plasma and Application of the Analytical Method to Human Pharmacokinetic Studies Following Two Week-Repeated Administration of Red Ginseng Extract. Molecules 2019, 24, 2618. [Google Scholar]
- Hou, M.; Wang, R.; Zhao, S.; Wang, Z. Ginsenosides in Panax genus and their biosynthesis. Acta Pharm. Sinica. B 2021, 11, 1813–1834. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Zhang, D.; Yang, D.C. Biosynthesis and biotechnological production of ginsenosides. Biotechnol. Adv. 2015, 33 Pt 1, 717–735. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, H.; Wang, R.; Lu, X.; Wang, Y.; Duan, M.; Li, H.; Fan, X.; Wang, S. Pharmacokinetics, tissue distribution and excretion of saponins after intravenous administration of ShenMai Injection in rats. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2019, 1128, 121777. [Google Scholar] [CrossRef]
- An, H.M.; Li, M.N.; Yang, H.; Pang, H.Q.; Qu, C.; Xu, Y.; Liu, R.Z.; Peng, C.; Li, P.; Gao, W. A validated UHPLC-MS/MS method for pharmacokinetic and brain distribution studies of twenty constituents in rat after oral administration of Jia-Wei-Qi-Fu-Yin. J. Pharm. Biomed. Anal. 2021, 202, 114140. [Google Scholar] [CrossRef]
- Wang, W.; Liao, Q.P.; Quan, L.H.; Liu, C.Y.; Chang, Q.; Liu, X.M.; Liao, Y.H. The effect of Acorus gramineus on the bioavailabilities and brain concentrations of ginsenosides Rg1, Re and Rb1 after oral administration of Kai-Xin-San preparations in rats. J. Ethnopharmacol. 2010, 131, 313–320. [Google Scholar] [CrossRef]
- Wei, W.; Li, Z.; Li, H.; An, Y.; Qu, H.; Yao, C.; Zhang, J.; Li, J.; Zhang, G.; Shi, Y.; et al. Exploration of tissue distribution of ginsenoside Rg1 by LC-MS/MS and nanospray desorption electrospray ionization mass spectrometry. J. Pharm. Biomed. Anal. 2021, 198, 113999. [Google Scholar] [CrossRef]
- Shi, J.; Xue, W.; Zhao, W.J.; Li, K.X. Pharmacokinetics and dopamine/acetylcholine releasing effects of ginsenoside Re in hippocampus and mPFC of freely moving rats. Acta Pharmacol. Sin. 2013, 34, 214–220. [Google Scholar] [CrossRef]
- Guo, Q.; Li, P.; Wang, Z.; Cheng, Y.; Wu, H.; Yang, B.; Du, S.; Lu, Y. Brain distribution pharmacokinetics and integrated pharmacokinetics of Panax Notoginsenoside R1, Ginsenosides Rg1, Rb1, Re and Rd in rats after intranasal administration of Panax Notoginseng Saponins assessed by UPLC/MS/MS. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2014, 969, 264–271. [Google Scholar] [CrossRef]
- Baker, D.J.; Petersen, R.C. Cellular senescence in brain aging and neurodegenerative diseases: Evidence and perspectives. J. Clin. Investig. 2018, 128, 1208–1216. [Google Scholar] [CrossRef]
- Dugger, B.N.; Dickson, D.W. Pathology of Neurodegenerative Diseases. Cold Spring Harb. Perspect. Biol. 2017, 9, a028035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondi, M.W.; Edmonds, E.C.; Salmon, D.P. Alzheimer’s Disease: Past, Present, and Future. J. Int. Neuropsychol. Soc. JINS 2017, 23, 818–831. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.H.; Lee, S.T.; Chu, K.; Oh, M.J.; Park, H.J.; Shim, J.Y.; Kim, M. An open-label trial of Korean red ginseng as an adjuvant treatment for cognitive impairment in patients with Alzheimer’s disease. Eur. J. Neurol. 2008, 15, 865–868. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.T.; Chu, K.; Sim, J.Y.; Heo, J.H.; Kim, M. Panax ginseng enhances cognitive performance in Alzheimer disease. Alzheimer Dis. Assoc. Disord. 2008, 22, 222–226. [Google Scholar] [CrossRef]
- Namgung, E.; Kim, J.; Jeong, H.; Hong, G.; Kim, M.; Kim, R.Y.; Kim, S.; Lyoo, I.K. Effects of Korean red ginseng on human gray matter volume and cognitive function: A voxel-based morphometry study. Hum. Psychopharmacol. 2021, 36, e2767. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Jung, S.W.; Kim, S.Y.; Cho, I.H.; Kim, H.C.; Rhim, H.; Kim, M.; Nah, S.Y. Panax ginseng as an adjuvant treatment for Alzheimer’ disease. J. Ginseng Res. 2018, 42, 401–411. [Google Scholar] [CrossRef]
- Chen, E.Y.; Hui, C.L. HT1001, a proprietary North American ginseng extract, improves working memory in schizophrenia: A double-blind, placebo-controlled study. Phytother. Res. PTR 2012, 26, 1166–1172. [Google Scholar] [CrossRef]
- Baek, J.H.; Heo, J.Y.; Fava, M.; Mischoulon, D.; Choi, K.W.; Na, E.J.; Cho, H.; Jeon, H.J. Effect of Korean Red Ginseng in individuals exposed to high stress levels: A 6-week, double-blind, randomized, placebo-controlled trial. J. Ginseng Res. 2019, 43, 402–407. [Google Scholar] [CrossRef]
- Scholey, A.; Ossoukhova, A.; Owen, L.; Ibarra, A.; Pipingas, A.; He, K.; Roller, M.; Stough, C. Effects of American ginseng (Panax quinquefolius) on neurocognitive function: An acute, randomised, double-blind, placebo-controlled, crossover study. Psychopharmacology 2010, 212, 345–356. [Google Scholar] [CrossRef]
- Reay, J.L.; Kennedy, D.O.; Scholey, A.B. Effects of Panax ginseng, consumed with and without glucose, on blood glucose levels and cognitive performance during sustained ‘mentally demanding’ tasks. J. Psychopharmacol. 2006, 20, 771–781. [Google Scholar] [CrossRef]
- Sheng, C.; Peng, W.; Xia, Z.A.; Wang, Y.; Chen, Z.; Su, N.; Wang, Z. The impact of ginsenosides on cognitive deficits in experimental animal studies of Alzheimer’s disease: A systematic review. BMC Complementary Altern. Med. 2015, 15, 386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Li, S.; Huang, H.; Lv, J.; Chen, S.; Pires Dias, A.C.; Li, Y.; Liu, X.; Wang, Q. Comparison of the Protective Effects of Ginsenosides Rb1 and Rg1 on Improving Cognitive Deficits in SAMP8 Mice Based on Anti-Neuroinflammation Mechanism. Front. Pharmacol. 2020, 11, 834. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.Q.; Huang, T.W.; Chen, L.M.; Pan, X.D.; Zhang, J.; Zhu, Y.G.; Chen, X.C. Ginsenoside Rg1 attenuates amyloid-beta content, regulates PKA/CREB activity, and improves cognitive performance in SAMP8 mice. J. Alzheimer’s Dis. JAD 2010, 19, 977–989. [Google Scholar] [CrossRef]
- Shi, R.; Zhang, S.; Cheng, G.; Yang, X.; Zhao, N.; Chen, C. Ginsenoside Rg1 and Acori Graminei Rhizoma Attenuates Neuron Cell Apoptosis by Promoting the Expression of miR-873-5p in Alzheimer’s Disease. Neurochem. Res. 2018, 43, 1529–1538. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.M.; Lin, Z.Y.; Zhu, Y.G.; Lin, N.; Zhang, J.; Pan, X.D.; Chen, X.C. Ginsenoside Rg1 attenuates β-amyloid generation via suppressing PPARγ-regulated BACE1 activity in N2a-APP695 cells. Eur. J. Pharmacol. 2012, 675, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Song, X.Y.; Hu, J.F.; Chu, S.F.; Zhang, Z.; Xu, S.; Yuan, Y.H.; Han, N.; Liu, Y.; Niu, F.; He, X.; et al. Ginsenoside Rg1 attenuates okadaic acid induced spatial memory impairment by the GSK3β/tau signaling pathway and the Aβ formation prevention in rats. Eur. J. Pharmacol. 2013, 710, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, M.; Ye, R.; Wang, W.; Liu, X.; Zhang, G.; Han, J.; Zhang, Y.; Wang, B.; Zhao, J.; et al. Ginsenoside Rd attenuates tau protein phosphorylation via the PI3K/AKT/GSK-3β pathway after transient forebrain ischemia. Neurochem. Res. 2014, 39, 1363–1373. [Google Scholar] [CrossRef]
- Zhang, H.; Su, Y.; Sun, Z.; Chen, M.; Han, Y.; Li, Y.; Dong, X.; Ding, S.; Fang, Z.; Li, W.; et al. Ginsenoside Rg1 alleviates Aβ deposition by inhibiting NADPH oxidase 2 activation in APP/PS1 mice. J. Ginseng Res. 2021, 45, 665–675. [Google Scholar] [CrossRef]
- Wang, L.; Lu, J.; Zeng, Y.; Guo, Y.; Wu, C.; Zhao, H.; Zheng, H.; Jiao, J. Improving Alzheimer’s disease by altering gut microbiota in tree shrews with ginsenoside Rg1. FEMS Microbiol. Lett. 2020, 367, fnaa011. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, L.; Lu, J.; Jiao, J.; Yang, Y.; Zhao, H.; Liang, Z.; Zheng, H. Ginsenoside Rg1 improves cognitive capability and affects the microbiota of large intestine of tree shrew model for Alzheimer’s disease. Mol. Med. Rep. 2021, 23, 291. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Yang, W.; Gao, S.; Lin, J.; Wang, T.; Zhou, K.; Hu, H. Ginsenoside Rb1 inhibit apoptosis in rat model of Alzheimer’s disease induced by Aβ(1-40). Am. J. Transl. Res. 2018, 10, 796–805. [Google Scholar] [PubMed]
- Lin, J.; Gao, S.; Wang, T.; Shen, Y.; Yang, W.; Li, Y.; Hu, H. Ginsenoside Rb1 improves learning and memory ability through its anti-inflammatory effect in Aβ(1-40) induced Alzheimer’s disease of rats. Am. J. Transl. Res. 2019, 11, 2955–2968. [Google Scholar]
- Zhao, H.H.; Di, J.; Liu, W.S.; Liu, H.L.; Lai, H.; Lü, Y.L. Involvement of GSK3 and PP2A in ginsenoside Rb1’s attenuation of aluminum-induced tau hyperphosphorylation. Behav. Brain Res. 2013, 241, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.Y.; Ju, S.H.; Oh, J.; Lee, S.K.; Kim, J.S. Neuroprotective and Cognition-Enhancing Effects of Compound K Isolated from Red Ginseng. J. Agric. Food Chem. 2016, 64, 2855–2864. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zong, X.; Huang, J.; Guan, Y.; Li, Y.; Du, T.; Liu, K.; Kang, X.; Dou, C.; Sun, X.; et al. Ginsenoside Rb1 regulates prefrontal cortical GABAergic transmission in MPTP-treated mice. Aging 2019, 11, 5008–5034. [Google Scholar] [CrossRef] [PubMed]
- Rudakewich, M.; Ba, F.; Benishin, C.G. Neurotrophic and neuroprotective actions of ginsenosides Rb(1) and Rg(1). Planta Med. 2001, 67, 533–537. [Google Scholar] [CrossRef]
- Yan, Y.T.; Li, S.D.; Li, C.; Xiong, Y.X.; Lu, X.H.; Zhou, X.F.; Yang, L.Q.; Pu, L.J.; Luo, H.Y. Panax notoginsenoside saponins Rb1 regulates the expressions of Akt/ mTOR/PTEN signals in the hippocampus after focal cerebral ischemia in rats. Behav. Brain Res. 2018, 345, 83–92. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, H.; Gu, W.; Liu, Y.; Zhang, M. Ginsenoside Rb1 protects hippocampal neurons from high glucose-induced neurotoxicity by inhibiting GSK3β-mediated CHOP induction. Mol. Med. Rep. 2014, 9, 1434–1438. [Google Scholar] [CrossRef]
- Yang, R.; Jiang, X.; He, X.; Liang, D.; Sun, S.; Zhou, G. Ginsenoside Rb1 Improves Cognitive Impairment Induced by Insulin Resistance through Cdk5/p35-NMDAR-IDE Pathway. BioMed Res. Int. 2020, 2020, 3905719. [Google Scholar] [CrossRef]
- Huang, L.; Peng, Z.; Lu, C.; Chen, Y.; Lv, J.W.; Qin, M.; Liao, D.F.; Liu, X.M.; Shi, Z. Ginsenoside Rg1 alleviates repeated alcohol exposure-induced psychomotor and cognitive deficits. Chin. Med. 2020, 15, 44. [Google Scholar] [CrossRef]
- Zhu, J.; Mu, X.; Zeng, J.; Xu, C.; Liu, J.; Zhang, M.; Li, C.; Chen, J.; Li, T.; Wang, Y. Ginsenoside Rg1 prevents cognitive impairment and hippocampus senescence in a rat model of D-galactose-induced aging. PLoS ONE 2014, 9, e101291. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.J.; Wang, L.; Gu, R.Z.; Zhang, W.H.; Lan, R.; Qin, X.Y. Ginsenoside Rg1 ameliorates the cognitive deficits in D-galactose and AlCl(3)-induced aging mice by restoring FGF2-Akt and BDNF-TrkB signaling axis to inhibit apoptosis. Int. J. Med. Sci. 2020, 17, 1048–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Q.; Ma, X.; Zhang, Z.J.; Sun, T.; Xia, F.; Zhao, G.; Wu, Y.M. Ginsenoside Reduces Cognitive Impairment During Chronic Cerebral Hypoperfusion Through Brain-Derived Neurotrophic Factor Regulated by Epigenetic Modulation. Mol. Neurobiol. 2017, 54, 2889–2900. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jiang, N.; Lv, J.; Huang, H.; Liu, X. Ginsenoside Rd reverses cognitive deficits by modulating BDNF-dependent CREB pathway in chronic restraint stress mice. Life Sci. 2020, 258, 118107. [Google Scholar] [CrossRef]
- Choi, S.Y.; Kim, K.J.; Song, J.H.; Lee, B.Y. Ginsenoside Rg5 prevents apoptosis by modulating heme-oxygenase-1/nuclear factor E2-related factor 2 signaling and alters the expression of cognitive impairment-associated genes in thermal stress-exposed HT22 cells. J. Ginseng Res. 2018, 42, 225–228. [Google Scholar] [CrossRef]
- Liu, Y.W.; Zhu, X.; Li, W.; Lu, Q.; Wang, J.Y.; Wei, Y.Q.; Yin, X.X. Ginsenoside Re attenuates diabetes-associated cognitive deficits in rats. Pharmacol. Biochem. Behav. 2012, 101, 93–98. [Google Scholar] [CrossRef]
- Hu, X.; De Silva, T.M.; Chen, J.; Faraci, F.M. Cerebral Vascular Disease and Neurovascular Injury in Ischemic Stroke. Circ. Res. 2017, 120, 449–471. [Google Scholar] [CrossRef]
- Shi, D.D.; Huang, Y.H.; Lai, C.S.W.; Dong, C.M.; Ho, L.C.; Li, X.Y.; Wu, E.X.; Li, Q.; Wang, X.M.; Chen, Y.J.; et al. Ginsenoside Rg1 Prevents Chemotherapy-Induced Cognitive Impairment: Associations with Microglia-Mediated Cytokines, Neuroinflammation, and Neuroplasticity. Mol. Neurobiol. 2019, 56, 5626–5642. [Google Scholar] [CrossRef]
- Kumar, A.; Rinwa, P.; Dhar, H. Microglial inhibitory effect of ginseng ameliorates cognitive deficits and neuroinflammation following traumatic head injury in rats. Inflammopharmacology 2014, 22, 155–167. [Google Scholar] [CrossRef]
- Rinwa, P.; Kumar, A. Modulation of nitrergic signalling pathway by American ginseng attenuates chronic unpredictable stress-induced cognitive impairment, neuroinflammation, and biochemical alterations. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2014, 387, 129–141. [Google Scholar] [CrossRef]
- Wang, Q.; Sun, L.H.; Jia, W.; Liu, X.M.; Dang, H.X.; Mai, W.L.; Wang, N.; Steinmetz, A.; Wang, Y.Q.; Xu, C.J. Comparison of ginsenosides Rg1 and Rb1 for their effects on improving scopolamine-induced learning and memory impairment in mice. Phytother. Res. PTR 2010, 24, 1748–1754. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Liu, Y.; Li, W.; Zhou, L.; Li, Q.; Wang, X.; He, P. A UPLC/MS-based metabolomics investigation of the protective effect of ginsenosides Rg1 and Rg2 in mice with Alzheimer’s disease. J. Ginseng Res. 2016, 40, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Yang, J.; Liu, C.; Xie, J.; Qiu, S.; Yang, X.; Wu, C. Pseudoginsenoside-F11 alleviates cognitive deficits and Alzheimer’s disease-type pathologies in SAMP8 mice. Pharmacol. Res. 2019, 139, 512–523. [Google Scholar] [CrossRef]
- Oh, J.; Kim, J.S. Compound K derived from ginseng: Neuroprotection and cognitive improvement. Food Funct. 2016, 7, 4506–4515. [Google Scholar] [CrossRef]
- Grodzicki, W.; Dziendzikowska, K. The Role of Selected Bioactive Compounds in the Prevention of Alzheimer’s Disease. Antioxidants 2020, 9, 229. [Google Scholar] [CrossRef] [PubMed]
- Coon, J.T.; Ernst, E. Panax ginseng: A systematic review of adverse effects and drug interactions. Drug Safety 2002, 25, 323–344. [Google Scholar] [CrossRef]
- Siegel, R.K. Ginseng abuse syndrome. Problems with the panacea. JAMA 1979, 241, 1614–1615. [Google Scholar] [CrossRef]
- Moss, J.; Yuan, C.S. Herbal medicines and perioperative care. Anesthesiology 2006, 105, 441–442. [Google Scholar] [CrossRef]
- Jalili, J.; Askeroglu, U.; Alleyne, B.; Guyuron, B. Herbal products that may contribute to hypertension. Plast. Reconstr. Surg. 2013, 131, 168–173. [Google Scholar] [CrossRef]
- Greenspan, E.M. Ginseng and vaginal bleeding. JAMA 1983, 249, 2018. [Google Scholar] [CrossRef]
- Oh, K.J.; Chae, M.J.; Lee, H.S.; Hong, H.D.; Park, K. Effects of Korean red ginseng on sexual arousal in menopausal women: Placebo-controlled, double-blind crossover clinical study. J. Sex. Med. 2010, 4 Pt 1, 1469–1477. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Yang, D.; Cheng, X.Y.; Yang, H.; Yang, X.H.; Liu, H.T.; Wang, R.; Zheng, P.; Yao, Y.; Li, J. Astragaloside IV Ameliorates Cognitive Impairment and Neuroinflammation in an Oligomeric Aβ Induced Alzheimer’s Disease Mouse Model via Inhibition of Microglial Activation and NADPH Oxidase Expression. Biol. Pharm. Bull. 2021, 44, 1688–1696. [Google Scholar] [CrossRef] [PubMed]
- Mazza, M.; Capuano, A.; Bria, P.; Mazza, S. Ginkgo biloba and donepezil: A comparison in the treatment of Alzheimer’s dementia in a randomized placebo-controlled double-blind study. Eur. J. Neurol. 2006, 13, 981–985. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; An, S.; Shao, T.; Xu, H.; Chen, H.; Ning, J.; Zhou, Y.; Chai, X. Active compounds of herbs ameliorate impaired cognition in APP/PS1 mouse model of Alzheimer’s disease. Aging 2019, 11, 11186–11201. [Google Scholar] [CrossRef]
- Tian, X.; Zhang, L.; Wang, J.; Dai, J.; Shen, S.; Yang, L.; Huang, P. The protective effect of hyperbaric oxygen and Ginkgo biloba extract on Aβ25-35-induced oxidative stress and neuronal apoptosis in rats. Behav. Brain Res. 2013, 242, 1–8. [Google Scholar] [CrossRef]
- Lee, Y.K.; Choi, I.S.; Ban, J.O.; Lee, H.J.; Lee, U.S.; Han, S.B.; Jung, J.K.; Kim, Y.H.; Kim, K.H.; Oh, K.W.; et al. 4-O-methylhonokiol attenuated β-amyloid-induced memory impairment through reduction of oxidative damages via inactivation of p38 MAP kinase. J. Nutr. Biochem. 2011, 22, 476–486. [Google Scholar] [CrossRef]
- Wang, C.; Xiao, Y.; Yang, B.; Wang, Z.; Wu, L.; Su, X.; Brantner, A.; Kuang, H.; Wang, Q. Isolation and screened neuroprotective active constituents from the roots and rhizomes of Valeriana amurensis. Fitoterapia 2014, 96, 48–55. [Google Scholar] [CrossRef]
- Jeong, H.Y.; Kim, J.Y.; Lee, H.K.; Ha do, T.; Song, K.S.; Bae, K.; Seong, Y.H. Leaf and stem of Vitis amurensis and its active components protect against amyloid β protein (25-35)-induced neurotoxicity. Arch. Pharmacal Res. 2010, 33, 1655–1664. [Google Scholar] [CrossRef]
- Li, D.D.; Fan, H.X.; Yang, R.; Li, Y.Y.; Zhang, F.; Shi, J.S. Dendrobium Nobile Lindl. Alkaloid Suppresses NLRP3-Mediated Pyroptosis to Alleviate LPS-Induced Neurotoxicity. Front. Pharmacol. 2022, 13, 846541. [Google Scholar]
- Ko, S.Y.; Lee, H.E.; Park, S.J.; Jeon, S.J.; Kim, B.; Gao, Q.; Jang, D.S.; Ryu, J.H. Spinosin, a C-Glucosylflavone, from Zizyphus jujuba var. spinosa Ameliorates Aβ1-42 Oligomer-Induced Memory Impairment in Mice. Biomol. Ther. 2015, 23, 156–164. [Google Scholar] [CrossRef]
- Hampel, H.; Mesulam, M.M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; et al. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain A J. Neurol. 2018, 141, 1917–1933. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.J.; Lin, C.H.; Lane, H.Y. Involvement of Cholinergic, Adrenergic, and Glutamatergic Network Modulation with Cognitive Dysfunction in Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 2283. [Google Scholar] [CrossRef] [PubMed]
- Van der Zee, S.; Müller, M.; Kanel, P.; van Laar, T.; Bohnen, N.I. Cholinergic Denervation Patterns Across Cognitive Domains in Parkinson’s Disease. Mov. Disord. Off. J. Mov. Disord. Soc. 2021, 36, 642–650. [Google Scholar]
- Choi, R.J.; Roy, A.; Jung, H.J.; Ali, M.Y.; Min, B.S.; Park, C.H.; Yokozawa, T.; Fan, T.P.; Choi, J.S.; Jung, H.A. BACE1 molecular docking and anti-Alzheimer’s disease activities of ginsenosides. J. Ethnopharmacol. 2016, 190, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liang, X.; Jin, P.; Li, N.; Zhang, Q.; Yan, W.; Zhang, H.; Sun, J. Screening and determination for potential acetylcholinesterase inhibitory constituents from ginseng stem-leaf saponins using ultrafiltration (UF)-LC-ESI-MS(2). Phytochem. Anal. PCA 2019, 30, 26–33. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, Y.; Li, K.X.; Xue, W. Pharmacokinetics and acetylcholine releasing effects of ginsenoside Rg1 in hippocampus of beta-amyloid model rats. J. Asian Nat. Prod. Res. 2019, 21, 772–781. [Google Scholar] [CrossRef]
- Kim, E.J.; Jung, I.H.; Van Le, T.K.; Jeong, J.J.; Kim, N.J.; Kim, D.H. Ginsenosides Rg5 and Rh3 protect scopolamine-induced memory deficits in mice. J. Ethnopharmacol. 2013, 146, 294–929. [Google Scholar] [CrossRef]
- An, K.S.; Choi, Y.O.; Lee, S.M.; Ryu, H.Y.; Kang, S.J.; Yeon, Y.; Kim, Y.R.; Lee, J.G.; Kim, C.J.; Lee, Y.J.; et al. Ginsenosides Rg5 and Rk1 Enriched Cultured Wild Ginseng Root Extract Bioconversion of Pediococcus pentosaceus HLJG0702: Effect on Scopolamine-Induced Memory Dysfunction in Mice. Nutrients 2019, 11, 1120. [Google Scholar] [CrossRef]
- Lu, C.; Lv, J.; Dong, L.; Jiang, N.; Wang, Y.; Wang, Q.; Li, Y.; Chen, S.; Fan, B.; Wang, F.; et al. Neuroprotective effects of 20(S)-protopanaxatriol (PPT) on scopolamine-induced cognitive deficits in mice. Phytother. Res. PTR 2018, 32, 1056–1063. [Google Scholar] [CrossRef]
- Kim, M.S.; Yu, J.M.; Kim, H.J.; Kim, H.B.; Kim, S.T.; Jang, S.K.; Choi, Y.W.; Lee, D.I.; Joo, S.S. Ginsenoside Re and Rd enhance the expression of cholinergic markers and neuronal differentiation in Neuro-2a cells. Biol. Pharm. Bull. 2014, 37, 826–833. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, H.; Xu, H.; Zheng, Y.; Wu, T.; Lian, Y. Ginsenoside Rb1 ameliorates cisplatin-induced learning and memory impairments. J. Ginseng Res. 2019, 43, 499–507. [Google Scholar] [CrossRef]
- Kim, J.M.; Park, C.H.; Park, S.K.; Seung, T.W.; Kang, J.Y.; Ha, J.S.; Lee, D.S.; Lee, U.; Kim, D.O.; Heo, H.J. Ginsenoside Re Ameliorates Brain Insulin Resistance and Cognitive Dysfunction in High Fat Diet-Induced C57BL/6 Mice. J. Agric. Food Chem. 2017, 65, 2719–2729. [Google Scholar] [CrossRef]
- Jin, Y.; Peng, J.; Wang, X.; Zhang, D.; Wang, T. Ameliorative Effect of Ginsenoside Rg1 on Lipopolysaccharide-Induced Cognitive Impairment: Role of Cholinergic System. Neurochem. Res. 2017, 42, 1299–1307. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Halliwell, B. Oxidative stress, dysfunctional glucose metabolism and Alzheimer disease. Nat. Rev. Neurosci. 2019, 20, 148–160. [Google Scholar] [CrossRef]
- Chen, L.; Yao, H.; Chen, X.; Wang, Z.; Xiang, Y.; Xia, J.; Liu, Y.; Wang, Y. Ginsenoside Rg1 Decreases Oxidative Stress and Down-Regulates Akt/mTOR Signalling to Attenuate Cognitive Impairment in Mice and Senescence of Neural Stem Cells Induced by D-Galactose. Neurochem. Res. 2018, 43, 430–440. [Google Scholar] [CrossRef]
- Miao, H.H.; Zhang, Y.; Ding, G.N.; Hong, F.X.; Dong, P.; Tian, M. Ginsenoside Rb1 Attenuates Isoflurane/surgery-induced Cognitive Dysfunction via Inhibiting Neuroinflammation and Oxidative Stress. Biomed. Environ. Sci. BES 2017, 30, 363–372. [Google Scholar]
- Zhang, Z.; Yang, H.; Yang, J.; Xie, J.; Xu, J.; Liu, C.; Wu, C. Pseudoginsenoside-F11 attenuates cognitive impairment by ameliorating oxidative stress and neuroinflammation in D-galactose-treated mice. Int. Immunopharmacol. 2019, 67, 78–86. [Google Scholar] [CrossRef]
- Nan, F.; Sun, G.; Xie, W.; Ye, T.; Sun, X.; Zhou, P.; Dong, X.; Sun, J.; Sun, X.; Zhang, M. Ginsenoside Rb1 mitigates oxidative stress and apoptosis induced by methylglyoxal in SH-SY5Y cells via the PI3K/Akt pathway. Mol. Cell. Probes 2019, 48, 101469. [Google Scholar] [CrossRef]
- Lu, C.; Wang, Y.; Lv, J.; Jiang, N.; Fan, B.; Qu, L.; Li, Y.; Chen, S.; Wang, F.; Liu, X. Ginsenoside Rh2 reverses sleep deprivation-induced cognitive deficit in mice. Behav. Brain Res. 2018, 349, 109–115. [Google Scholar] [CrossRef]
- Fernández-Moriano, C.; González-Burgos, E.; Iglesias, I.; Lozano, R.; Gómez-Serranillos, M.P. Evaluation of the adaptogenic potential exerted by ginsenosides Rb1 and Rg1 against oxidative stress-mediated neurotoxicity in an in vitro neuronal model. PLoS ONE 2017, 12, e0182933. [Google Scholar] [CrossRef]
- Liu, J.; Yan, X.; Li, L.; Zhu, Y.; Qin, K.; Zhou, L.; Sun, D.; Zhang, X.; Ye, R.; Zhao, G. Ginsennoside rd attenuates cognitive dysfunction in a rat model of Alzheimer’s disease. Neurochem. Res. 2012, 37, 2738–2747. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.M.; Liu, M.Y.; Wang, F.; Wei, M.J.; Wang, S.; Wu, C.F.; Yang, J.Y. Anti-amnesic effect of pseudoginsenoside-F11 in two mouse models of Alzheimer’s disease. Pharmacol. Biochem. Behav. 2013, 106, 57–67. [Google Scholar] [CrossRef]
- Cheon, S.Y.; Cho, K.J. Pathological role of apoptosis signal-regulating kinase 1 in human diseases and its potential as a therapeutic target for cognitive disorders. J. Mol. Med. 2019, 97, 153–161. [Google Scholar] [CrossRef]
- Li, Y.; Wang, F.; Luo, Y. Ginsenoside Rg1 protects against sepsis-associated encephalopathy through beclin 1-independent autophagy in mice. J. Surg. Res. 2017, 207, 181–189. [Google Scholar] [CrossRef]
- Leigh, S.J.; Morris, M.J. Diet, inflammation and the gut microbiome: Mechanisms for obesity-associated cognitive impairment. Biochim. Et Biophys. Acta. Mol. Basis Dis. 2020, 1866, 165767. [Google Scholar] [CrossRef]
- Den, H.; Dong, X.; Chen, M.; Zou, Z. Efficacy of probiotics on cognition, and biomarkers of inflammation and oxidative stress in adults with Alzheimer’s disease or mild cognitive impairment–A meta-analysis of randomized controlled trials. Aging 2020, 12, 4010–4039. [Google Scholar] [CrossRef]
- Holmes, C. Review: Systemic inflammation and Alzheimer’s disease. Neuropathol. Appl. Neurobiol. 2013, 39, 51–68. [Google Scholar] [CrossRef]
- Pugazhenthi, S.; Qin, L.; Reddy, P.H. Common neurodegenerative pathways in obesity, diabetes, and Alzheimer’s disease. Biochim. Biophys. Acta. Mol. Basis Dis. 2017, 1863, 1037–1045. [Google Scholar] [CrossRef]
- Xu, K.; Zhang, Y.; Wang, Y.; Ling, P.; Xie, X.; Jiang, C.; Zhang, Z.; Lian, X.Y. Ginseng Rb fraction protects glia, neurons and cognitive function in a rat model of neurodegeneration. PLoS ONE 2014, 9, e101077. [Google Scholar] [CrossRef]
- Du, Y.; Fu, M.; Wang, Y.T.; Dong, Z. Neuroprotective Effects of Ginsenoside Rf on Amyloid-β-Induced Neurotoxicity in vitro and in vivo. J. Alzheimer’s Dis. JAD 2018, 64, 309–322. [Google Scholar] [CrossRef]
- Chu, S.; Gu, J.; Feng, L.; Liu, J.; Zhang, M.; Jia, X.; Liu, M.; Yao, D. Ginsenoside Rg5 improves cognitive dysfunction and beta-amyloid deposition in STZ-induced memory impaired rats via attenuating neuroinflammatory responses. Int. Immunopharmacol. 2014, 19, 317–326. [Google Scholar] [CrossRef]
- Li, C.W.; Deng, M.Z.; Gao, Z.J.; Dang, Y.Y.; Zheng, G.D.; Yang, X.J.; Chao, Y.X.; Cai, Y.F.; Wu, X.L. Effects of compound K, a metabolite of ginsenosides, on memory and cognitive dysfunction in db/db mice involve the inhibition of ER stress and the NLRP3 inflammasome pathway. Food Funct. 2020, 11, 4416–4427. [Google Scholar] [CrossRef]
- Price, R.B.; Duman, R. Neuroplasticity in cognitive and psychological mechanisms of depression: An integrative model. Mol. Psychiatry 2020, 25, 530–543. [Google Scholar] [CrossRef]
- Li, S.; Selkoe, D.J. A mechanistic hypothesis for the impairment of synaptic plasticity by soluble Aβ oligomers from Alzheimer’s brain. J. Neurochem. 2020, 154, 583–597. [Google Scholar] [CrossRef]
- Li, F.; Wu, X.; Li, J.; Niu, Q. Ginsenoside Rg1 ameliorates hippocampal long-term potentiation and memory in an Alzheimer’s disease model. Mol. Med. Rep. 2016, 13, 4904–4910. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Y.; Zhang, J.T. Effects of ginsenoside Rg1 on synaptic plasticity of freely moving rats and its mechanism of action. Acta Pharmacol. Sin. 2001, 22, 657–662. [Google Scholar]
- Yang, L.; Zhang, J.; Zheng, K.; Shen, H.; Chen, X. Long-term ginsenoside Rg1 supplementation improves age-related cognitive decline by promoting synaptic plasticity associated protein expression in C57BL/6J mice. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2014, 69, 282–294. [Google Scholar] [CrossRef]
- Zhu, G.; Wang, Y.; Li, J.; Wang, J. Chronic treatment with ginsenoside Rg1 promotes memory and hippocampal long-term potentiation in middle-aged mice. Neuroscience 2015, 292, 81–89. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, W.; Zhang, B.; Yin, Y.; Zhang, J.; Huang, D.; Huang, R.; Li, W.; Li, W. Ginsenoside Rg1 protects against neuronal degeneration induced by chronic dexamethasone treatment by inhibiting NLRP-1 inflammasomes in mice. Int. J. Mol. Med. 2017, 40, 1134–1142. [Google Scholar] [CrossRef]
- Qi, D.; Zhu, Y.; Wen, L.; Liu, Q.; Qiao, H. Ginsenoside Rg1 restores the impairment of learning induced by chronic morphine administration in rats. J. Psychopharmacol. 2009, 23, 74–83. [Google Scholar] [CrossRef]
- Mook-Jung, I.; Hong, H.S.; Boo, J.H.; Lee, K.H.; Yun, S.H.; Cheong, M.Y.; Joo, I.; Huh, K.; Jung, M.W. Ginsenoside Rb1 and Rg1 improve spatial learning and increase hippocampal synaptophysin level in mice. J. Neurosci. Res. 2001, 63, 509–515. [Google Scholar] [CrossRef]
- Liu, Z.J.; Zhao, M.; Zhang, Y.; Xue, J.F.; Chen, N.H. Ginsenoside Rg1 promotes glutamate release via a calcium/calmodulin-dependent protein kinase II-dependent signaling pathway. Brain Res. 2010, 1333, 1–8. [Google Scholar] [CrossRef]
- Fan, C.; Zhu, X.; Song, Q.; Wang, P.; Liu, Z.; Yu, S.Y. MiR-134 modulates chronic stress-induced structural plasticity and depression-like behaviors via downregulation of Limk1/cofilin signaling in rats. Neuropharmacology 2018, 131, 364–376. [Google Scholar] [CrossRef]
- Qu, S.; Meng, X.; Liu, Y.; Zhang, X.; Zhang, Y. Ginsenoside Rb1 prevents MPTP-induced changes in hippocampal memory via regulation of the α-synuclein/PSD-95 pathway. Aging 2019, 11, 1934–1964. [Google Scholar] [CrossRef]
- Jiang, N.; Wang, K.; Zhang, Y.; Huang, H.; Lv, J.W.; Wang, Q.; Wang, H.X.; Xia, T.J.; Liu, X.M. Protective effect of ginsenoside Rb1 against chronic restraint stress (CRS)-induced memory impairments in rats. Behav. Brain Res. 2021, 405, 113146. [Google Scholar] [CrossRef]
- Liu, L.; Hoang-Gia, T.; Wu, H.; Lee, M.R.; Gu, L.; Wang, C.; Yun, B.S.; Wang, Q.; Ye, S.; Sung, C.K. Ginsenoside Rb1 improves spatial learning and memory by regulation of cell genesis in the hippocampal subregions of rats. Brain Res. 2011, 1382, 147–154. [Google Scholar] [CrossRef]
- Abe, K.; Cho, S.I.; Kitagawa, I.; Nishiyama, N.; Saito, H. Differential effects of ginsenoside Rb1 and malonylginsenoside Rb1 on long-term potentiation in the dentate gyrus of rats. Brain Res. 1994, 649, 7–11. [Google Scholar] [CrossRef]
- Martini, M.; De Santis, M.C.; Braccini, L.; Gulluni, F.; Hirsch, E. PI3K/AKT signaling pathway and cancer: An updated review. Ann. Med. 2014, 46, 372–383. [Google Scholar] [CrossRef]
- Borrie, S.C.; Brems, H.; Legius, E.; Bagni, C. Cognitive Dysfunctions in Intellectual Disabilities: The Contributions of the Ras-MAPK and PI3K-AKT-mTOR Pathways. Annu. Rev. Genom. Hum. Genet. 2017, 18, 115–142. [Google Scholar] [CrossRef]
- Stitt, T.N.; Drujan, D.; Clarke, B.A.; Panaro, F.; Timofeyva, Y.; Kline, W.O.; Gonzalez, M.; Yancopoulos, G.D.; Glass, D.J. The IGF-1/PI3K/Akt pathway prevents expression of muscle atrophy-induced ubiquitin ligases by inhibiting FOXO transcription factors. Mol. Cell 2004, 14, 395–403. [Google Scholar] [CrossRef]
- Dudka, A.A.; Sweet, S.M.; Heath, J.K. Signal transducers and activators of transcription-3 binding to the fibroblast growth factor receptor is activated by receptor amplification. Cancer Res. 2010, 70, 3391–3401. [Google Scholar] [CrossRef]
- Rai, S.N.; Dilnashin, H.; Birla, H.; Singh, S.S.; Zahra, W.; Rathore, A.S.; Singh, B.K.; Singh, S.P. The Role of PI3K/Akt and ERK in Neurodegenerative Disorders. Neurotox. Res. 2019, 35, 775–795. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Z.; Wang, H.; Cai, N.; Zhou, S.; Zhao, Y.; Chen, X.; Zheng, S.; Si, Q.; Zhang, W. Neuroprotective effect of ginsenoside Rg1 prevents cognitive impairment induced by isoflurane anesthesia in aged rats via antioxidant, anti-inflammatory and anti-apoptotic effects mediated by the PI3K/AKT/GSK-3β pathway. Mol. Med. Rep. 2016, 14, 2778–2784. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Shan, R.; Cao, Y.; Zhou, Y.; Liu, C.; Fan, Y. Protective effects of ginsenoside Rg2 against memory impairment and neuronal death induced by Aβ25-35 in rats. J. Ethnopharmacol. 2021, 266, 113466. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Hu, G.; Yan, W.; Chen, T.; Yang, F.; Zhang, X.; Zhao, G.; Liu, J. Ginsenoside Rd promotes non-amyloidogenic pathway of amyloid precursor protein processing by regulating phosphorylation of estrogen receptor alpha. Life Sci. 2017, 168, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Zong, W.; Zeng, X.; Chen, S.; Chen, L.; Zhou, L.; Wang, X.; Gao, Q.; Zeng, G.; Hu, K.; Ouyang, D. Ginsenoside compound K attenuates cognitive deficits in vascular dementia rats by reducing the Aβ deposition. J. Pharmacol. Sci. 2019, 139, 223–230. [Google Scholar] [CrossRef]
- Wang, F.; Wang, L.; Wang, Y.; Li, D.; Hu, T.; Sun, M.; Lei, P. Exogenous IGF-1 improves cognitive function in rats with high-fat diet consumption. J. Mol. Endocrinol. 2020, 64, 115–123. [Google Scholar] [CrossRef]
- Wang, Z.; Ye, Z.; Huang, G.; Wang, N.; Wang, E.; Guo, Q. Sevoflurane Post-conditioning Enhanced Hippocampal Neuron Resistance to Global Cerebral Ischemia Induced by Cardiac Arrest in Rats through PI3K/Akt Survival Pathway. Front. Cell. Neurosci. 2016, 10, 271. [Google Scholar] [CrossRef]
- Xie, L.; Fang, Q.; Wei, X.; Zhou, L.; Wang, S. Exogenous insulin-like growth factor 1 attenuates sevoflurane anesthesia-induced cognitive dysfunction in aged rats. J. Neurophysiol. 2021, 125, 2117–2124. [Google Scholar] [CrossRef]
- Ma, J.; Jiang, Q.; Xu, J.; Sun, Q.; Qiao, Y.; Chen, W.; Wu, Y.; Wang, Y.; Xiao, Q.; Liu, J.; et al. Plasma insulin-like growth factor 1 is associated with cognitive impairment in Parkinson′s disease. Dement. Geriatr. Cogn. Disord. 2015, 39, 251–256. [Google Scholar] [CrossRef]
- Westwood, A.J.; Beiser, A.; Decarli, C.; Harris, T.B.; Chen, T.C.; He, X.M.; Roubenoff, R.; Pikula, A.; Au, R.; Braverman, L.E.; et al. Insulin-like growth factor-1 and risk of Alzheimer dementia and brain atrophy. Neurology 2014, 82, 1613–1619. [Google Scholar] [CrossRef]
- Kiyota, T.; Ingraham, K.L.; Jacobsen, M.T.; Xiong, H.; Ikezu, T. FGF2 gene transfer restores hippocampal functions in mouse models of Alzheimer’s disease and has therapeutic implications for neurocognitive disorders. Proc. Natl. Acad. Sci. USA 2011, 108, E1339–E1348. [Google Scholar] [CrossRef] [PubMed]
- Lang, X.Y.; Hu, Y.; Bai, J.P.; Wang, J.; Qin, X.Y.; Lan, R. Coeloglossum viride Var. Bracteatum Extract Attenuates MPTP-Induced Neurotoxicity in vivo by Restoring BDNF-TrkB and FGF2-Akt Signaling Axis and Inhibiting RIP1-Driven Inflammation. Front. Pharmacol. 2022, 13, 903235. [Google Scholar] [CrossRef] [PubMed]
- Lixing, X.; Zhouye, J.; Liting, G.; Ruyi, Z.; Rong, Q.; Shiping, M. Saikosaponin-d-mediated downregulation of neurogenesis results in cognitive dysfunction by inhibiting Akt/Foxg-1 pathway in mice. Toxicol. Lett. 2018, 284, 79–85. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Park, J.S.; Jung, J.S.; Kim, D.H.; Kim, H.S. Anti-inflammatory effect of ginsenoside Rg5 in lipopolysaccharide-stimulated BV2 microglial cells. Int. J. Mol. Sci. 2013, 14, 9820–9833. [Google Scholar] [CrossRef]
- Hooper, C.; Killick, R.; Lovestone, S. The GSK3 hypothesis of Alzheimer’s disease. J. Neurochem. 2008, 104, 1433–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, J.J.; Hernández, F.; Gómez-Ramos, P.; Morán, M.A.; Hen, R.; Avila, J. Decreased nuclear beta-catenin, tau hyperphosphorylation and neurodegeneration in GSK-3beta conditional transgenic mice. EMBO J. 2001, 20, 27–39. [Google Scholar] [CrossRef]
- Hernández, F.; Borrell, J.; Guaza, C.; Avila, J.; Lucas, J.J. Spatial learning deficit in transgenic mice that conditionally over-express GSK-3beta in the brain but do not form tau filaments. J. Neurochem. 2002, 83, 1529–1533. [Google Scholar] [CrossRef]
- Wang, S.; He, B.; Hang, W.; Wu, N.; Xia, L.; Wang, X.; Zhang, Q.; Zhou, X.; Feng, Z.; Chen, Q.; et al. Berberine Alleviates Tau Hyperphosphorylation and Axonopathy-Associated with Diabetic Encephalopathy via Restoring PI3K/Akt/GSK3β Pathway. J. Alzheimer’s Dis. JAD 2018, 65, 1385–1400. [Google Scholar] [CrossRef]
- Stratmann, G.; Sall, J.W.; May, L.D.; Loepke, A.W.; Lee, M.T. Beyond anesthetic properties: The effects of isoflurane on brain cell death, neurogenesis, and long-term neurocognitive function. Anesth. Analg. 2010, 110, 431–437. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, L.; Zhang, C.; Guo, Y.; Li, J.; Wu, C.; Jiao, J.; Zheng, H. Ginsenoside Rg1 improves Alzheimer’s disease by regulating oxidative stress, apoptosis, and neuroinflammation through Wnt/GSK-3β/β-catenin signaling pathway. Chem. Biol. Drug Des. 2022, 99, 884–896. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Driving neural regeneration through the mammalian target of rapamycin. Neural Regen. Res. 2014, 9, 1413–1417. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.P.; Ding, H.; Yang, X.Q.; Li, J.X.; Tang, B.; Liu, X.D.; Tang, Y.H.; Deng, C.Q. Synergism and mechanism of Astragaloside IV combined with Ginsenoside Rg1 against autophagic injury of PC12 cells induced by oxygen glucose deprivation/reoxygenation. Biomed. Pharmacother. 2017, 89, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Lv, J.; Lu, J.; Fan, L.; Huang, X.; Hu, L.; Wang, J.; Shen, X. Protopanaxadiol derivative DDPU improves behavior and cognitive deficit in AD mice involving regulation of both ER stress and autophagy. Neuropharmacology 2018, 130, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, D.; Liu, J.; Diao, L.; Ling, S.; Li, Y.; Gao, J.; Fan, Q.; Sun, W.; Li, Q.; et al. Dammarane Sapogenins Ameliorates Neurocognitive Functional Impairment Induced by Simulated Long-Duration Spaceflight. Front. Pharmacol. 2017, 8, 315. [Google Scholar] [CrossRef]
- Yu, F.; Sugawara, T.; Maier, C.M.; Hsieh, L.B.; Chan, P.H. Akt/Bad signaling and motor neuron survival after spinal cord injury. Neurobiol. Dis. 2005, 20, 491–499. [Google Scholar] [CrossRef]
- Cui, J.; Wang, J.; Zheng, M.; Gou, D.; Liu, C.; Zhou, Y. Ginsenoside Rg2 protects PC12 cells against β-amyloid(25-35)-induced apoptosis via the phosphoinositide 3-kinase/Akt pathway. Chem. Biol. Interact. 2017, 275, 152–161. [Google Scholar] [CrossRef]
- Amidfar, M.; de Oliveira, J.; Kucharska, E.; Budni, J.; Kim, Y.K. The role of CREB and BDNF in neurobiology and treatment of Alzheimer’s disease. Life Sci. 2020, 257, 118020. [Google Scholar] [CrossRef]
- Tardito, D.; Perez, J.; Tiraboschi, E.; Musazzi, L.; Racagni, G.; Popoli, M. Signaling pathways regulating gene expression, neuroplasticity, and neurotrophic mechanisms in the action of antidepressants: A critical overview. Pharmacol. Rev. 2006, 58, 115–134. [Google Scholar] [CrossRef]
- Rosa, E.; Fahnestock, M. CREB expression mediates amyloid β-induced basal BDNF downregulation. Neurobiol. Aging 2015, 36, 2406–2413. [Google Scholar] [CrossRef]
- Leal, G.; Bramham, C.R.; Duarte, C.B. BDNF and Hippocampal Synaptic Plasticity. Vitam. Horm. 2017, 104, 153–195. [Google Scholar] [PubMed]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in neuronal development and function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [PubMed]
- Bekinschtein, P.; Cammarota, M.; Medina, J.H. BDNF and memory processing. Neuropharmacology 2014, 76 Pt C, 677–683. [Google Scholar] [CrossRef]
- Pugazhenthi, S.; Wang, M.; Pham, S.; Sze, C.I.; Eckman, C.B. Downregulation of CREB expression in Alzheimer’s brain and in Aβ-treated rat hippocampal neurons. Mol. Neurodegener. 2011, 6, 60. [Google Scholar] [CrossRef]
- Kezhu, W.; Pan, X.; Cong, L.; Liming, D.; Beiyue, Z.; Jingwei, L.; Yanyan, Y.; Xinmin, L. Effects of Ginsenoside Rg1 on Learning and Memory in a Reward-directed Instrumental Conditioning Task in Chronic Restraint Stressed Rats. Phytother. Res. PTR 2017, 31, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, M.; Li, Y.; Quan, Q.; Wang, J. Cellular and molecular mechanisms underlying the action of ginsenoside Rg1 against Alzheimer’s disease. Neural Regen. Res. 2012, 7, 2860–2866. [Google Scholar]
- Han, J.; Oh, J.P.; Yoo, M.; Cui, C.H.; Jeon, B.M.; Kim, S.C.; Han, J.H. Minor ginsenoside F1 improves memory in APP/PS1 mice. Mol. Brain 2019, 12, 77. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Xue, J.; Lee, M.; Yu, J.; Sung, C. Long-term administration of ginsenoside Rh1 enhances learning and memory by promoting cell survival in the mouse hippocampus. Int. J. Mol. Med. 2014, 33, 234–240. [Google Scholar] [CrossRef]
- Klein, R.; Nanduri, V.; Jing, S.A.; Lamballe, F.; Tapley, P.; Bryant, S.; Cordon-Cardo, C.; Jones, K.R.; Reichardt, L.F.; Barbacid, M. The trkB tyrosine protein kinase is a receptor for brain-derived neurotrophic factor and neurotrophin-3. Cell 1991, 66, 395–403. [Google Scholar] [CrossRef]
- Guo, W.; Nagappan, G.; Lu, B. Differential effects of transient and sustained activation of BDNF-TrkB signaling. Dev. Neurobiol. 2018, 78, 647–659. [Google Scholar] [CrossRef]
- Chou, W.; Liu, Y.F.; Lin, C.H.; Lin, M.T.; Chen, C.C.; Liu, W.P.; Chang, C.P.; Chio, C.C. Exercise Rehabilitation Attenuates Cognitive Deficits in Rats with Traumatic Brain Injury by Stimulating the Cerebral HSP20/BDNF/TrkB Signalling Axis. Mol. Neurobiol. 2018, 55, 8602–8611. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Xiang, Y.; Qu, X.; Liu, H.; Liu, C.; Li, G.; Han, L.; Qin, X. Apelin-13 Suppresses Neuroinflammation Against Cognitive Deficit in a Streptozotocin-Induced Rat Model of Alzheimer’s Disease Through Activation of BDNF-TrkB Signaling Pathway. Front. Pharmacol. 2019, 10, 395. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Sun, S.; Pan, X.; Zheng, L.; Li, Y.; Yang, J.; Wu, C. Pseudoginsenoside-F11 improves long-term neurological function and promotes neurogenesis after transient cerebral ischemia in mice. Neurochem. Int. 2020, 133, 104586. [Google Scholar] [CrossRef]
- Walton, M.R.; Dragunow, I. Is CREB a key to neuronal survival? Trends Neurosci. 2000, 23, 48–53. [Google Scholar] [CrossRef]
- Landeira, B.S.; Santana, T.; Araújo, J.A.M.; Tabet, E.I.; Tannous, B.A.; Schroeder, T.; Costa, M.R. Activity-Independent Effects of CREB on Neuronal Survival and Differentiation during Mouse Cerebral Cortex Development. Cerebral Cortex 2018, 28, 538–548. [Google Scholar] [CrossRef]
- De Cesare, D.; Fimia, G.M.; Sassone-Corsi, P. Signaling routes to CREM and CREB: Plasticity in transcriptional activation. Trends Biochem. Sci. 1999, 24, 281–285. [Google Scholar] [CrossRef]
- Hu, J.F.; Xue, W.; Ning, N.; Yuan, Y.H.; Zhang, J.T.; Chen, N.H. Ginsenoside Rg1 activated CaMKIIalpha mediated extracellular signal-regulated kinase/mitogen activated protein kinase signaling pathway. Acta Pharmacol. Sin. 2008, 29, 1119–1126. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Li, Q.; Pei, X.; Zhang, Z.; Yang, R.; Wang, J.; Li, Y. Long-term ginsenoside administration prevents memory impairment in aged C57BL/6J mice by up-regulating the synaptic plasticity-related proteins in hippocampus. Behav. Brain Res. 2009, 201, 311–317. [Google Scholar] [CrossRef]
- Zhao, H.; Li, Q.; Zhang, Z.; Pei, X.; Wang, J.; Li, Y. Long-term ginsenoside consumption prevents memory loss in aged SAMP8 mice by decreasing oxidative stress and up-regulating the plasticity-related proteins in hippocampus. Brain Res. 2009, 1256, 111–122. [Google Scholar] [CrossRef]
- Silva-Islas, C.A.; Maldonado, P.D. Canonical and non-canonical mechanisms of Nrf2 activation. Pharmacol. Res. 2018, 134, 92–99. [Google Scholar] [CrossRef]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; Igarashi, K.; Engel, J.D.; Yamamoto, M. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes Dev. 1999, 13, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. CMLS 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, C.P.; Glass, C.A.; Montgomery, M.B.; Lindl, K.A.; Ritson, G.P.; Chia, L.A.; Hamilton, R.L.; Chu, C.T.; Jordan-Sciutto, K.L. Expression of Nrf2 in neurodegenerative diseases. J. Neuropathol. Exp. Neurol. 2007, 66, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Tapias, V.; Jainuddin, S.; Ahuja, M.; Stack, C.; Elipenahli, C.; Vignisse, J.; Gerges, M.; Starkova, N.; Xu, H.; Starkov, A.A.; et al. Benfotiamine treatment activates the Nrf2/ARE pathway and is neuroprotective in a transgenic mouse model of tauopathy. Hum. Mol. Genet. 2018, 27, 2874–2892. [Google Scholar] [CrossRef]
- Joshi, G.; Gan, K.A.; Johnson, D.A.; Johnson, J.A. Increased Alzheimer’s disease-like pathology in the APP/ PS1ΔE9 mouse model lacking Nrf2 through modulation of autophagy. Neurobiol. Aging 2015, 36, 664–679. [Google Scholar] [CrossRef]
- Wang, C.Y.; Zhang, Q.; Xun, Z.; Yuan, L.; Li, R.; Li, X.; Tian, S.Y.; Xin, N.; Xu, Y. Increases of iASPP-Keap1 interaction mediated by syringin enhance synaptic plasticity and rescue cognitive impairments via stabilizing Nrf2 in Alzheimer’s models. Redox Biol. 2020, 36, 101672. [Google Scholar] [CrossRef]
- Shi, Y.; Miao, W.; Teng, J.; Zhang, L. Ginsenoside Rb1 Protects the Brain from Damage Induced by Epileptic Seizure via Nrf2/ARE Signaling. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 45, 212–225. [Google Scholar] [CrossRef]
- Wang, H.; Lv, J.; Jiang, N.; Huang, H.; Wang, Q.; Liu, X. Ginsenoside Re protects against chronic restraint stress-induced cognitive deficits through regulation of NLRP3 and Nrf2 pathways in mice. Phytother. Res. PTR 2021, 35, 2523–2535. [Google Scholar] [CrossRef]
- Ju, S.; Seo, J.Y.; Lee, S.K.; Oh, J.; Kim, J.S. Oral administration of hydrolyzed red ginseng extract improves learning and memory capability of scopolamine-treated C57BL/6J mice via upregulation of Nrf2-mediated antioxidant mechanism. J. Ginseng Res. 2021, 45, 108–118. [Google Scholar] [CrossRef]
- Wang, C.; Fan, L.; Khawaja, R.R.; Liu, B.; Zhan, L.; Kodama, L.; Chin, M.; Li, Y.; Le, D.; Zhou, Y.; et al. Microglial NF-κB drives tau spreading and toxicity in a mouse model of tauopathy. Nat. Commun. 2022, 13, 1969. [Google Scholar] [CrossRef]
- Jiao, H.; Jia, J. Ginsenoside compound K acts via LRP1 to alleviate Amyloid β(42)-induced neuroinflammation in microglia by suppressing NF-κB. Biochem. Biophys. Res. Commun. 2022, 590, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Mayor, A.; Tschopp, J. The inflammasomes: Guardians of the body. Annu. Rev. Immunol. 2009, 27, 229–265. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, H.; Kouadir, M.; Song, H.; Shi, F. Recent advances in the mechanisms of NLRP3 inflammasome activation and its inhibitors. Cell Death Dis. 2019, 10, 128. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.Y.; Tang, L.Q. Roles of the NLRP3 inflammasome in the pathogenesis of diabetic nephropathy. Pharmacol. Res. 2016, 114, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, Y.; Zhang, J.; Yang, G. Mechanisms of NLRP3 Inflammasome Activation: Its Role in the Treatment of Alzheimer’s Disease. Neurochem. Res. 2020, 45, 2560–2572. [Google Scholar] [CrossRef]
- Qi, Y.; Klyubin, I.; Cuello, A.C.; Rowan, M.J. NLRP3-dependent synaptic plasticity deficit in an Alzheimer’s disease amyloidosis model in vivo. Neurobiol. Dis. 2018, 114, 24–30. [Google Scholar] [CrossRef]
- Ising, C.; Venegas, C.; Zhang, S.; Scheiblich, H.; Schmidt, S.V.; Vieira-Saecker, A.; Schwartz, S.; Albasset, S.; McManus, R.M.; Tejera, D.; et al. NLRP3 inflammasome activation drives tau pathology. Nature 2019, 575, 669–673. [Google Scholar] [CrossRef]
- Navarro-González, J.F.; Mora-Fernández, C.; Muros de Fuentes, M.; García-Pérez, J. Inflammatory molecules and pathways in the pathogenesis of diabetic nephropathy. Nat. Rev. Nephrol. 2011, 7, 327–340. [Google Scholar] [CrossRef]
- Chen, J.; Xuan, J.; Gu, Y.T.; Shi, K.S.; Xie, J.J.; Chen, J.X.; Zheng, Z.M.; Chen, Y.; Chen, X.B.; Wu, Y.S. Celastrol reduces IL-1β induced matrix catabolism, oxidative stress and inflammation in human nucleus pulposus cells and attenuates rat intervertebral disc degeneration in vivo. Biomed. Pharmacother. 2017, 91, 208–219. [Google Scholar] [CrossRef]
- Lee, B.; Sur, B.; Park, J.; Kim, S.H.; Kwon, S.; Yeom, M.; Shim, I.; Lee, H.; Hahm, D.H. Ginsenoside rg3 alleviates lipopolysaccharide-induced learning and memory impairments by anti-inflammatory activity in rats. Biomol. Ther. 2013, 21, 381–390. [Google Scholar] [CrossRef]
- Gu, Y.; Wang, G.J.; Sun, J.G.; Jia, Y.W.; Wang, W.; Xu, M.J.; Lv, T.; Zheng, Y.T.; Sai, Y. Pharmacokinetic characterization of ginsenoside Rh2, an anticancer nutrient from ginseng, in rats and dogs. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2009, 47, 2257–2268. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, H. Proof of the mysterious efficacy of ginseng: Basic and clinical trials: Metabolic activation of ginsenoside: Deglycosylation by intestinal bacteria and esterification with fatty acid. J. Pharmacol. Sci. 2004, 95, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, J.H.; Kim, J.E.; Kim, Y.S.; Ryu, C.H.; Lee, H.J.; Kim, H.M.; Jeon, H.; Won, H.J.; Lee, J.Y. Micro-/nano-sized delivery systems of ginsenosides for improved systemic bioavailability. J. Ginseng Res. 2018, 42, 361–369. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Saponins | Formula | R1(C3) | R2(C6) |
|---|---|---|---|---|
| Protopanaxadiol type | ||||
| (PPD-type) | Rb1 | C54H92O23 | Glc-Glc | H |
| Rb2 | C54H90O22 | Glc-Glc | H | |
| Rc | C53H90O22 | Glc-Glc | H | |
| Rd | C48H82O18 | Glc-Glc | H | |
| Rg3 | C42H72O13 | Glc-Glc | H | |
| Rh2 | C36H62O8 | Glc | H | |
| Compound K | C36H62O8 | H | H | |
| Protopanaxatriol type | ||||
| (PPT-type) | Rg1 | C42H72O14 | H | Glc |
| Rg2 | C42H72O13 | H | Glc-Rha | |
| Rh1 | C36H62O9 | H | Glc | |
| Re | C30H42O4 | H | Glc-Rha | |
| Rf | C42H72O14 | H | Glc-Glc | |
| F1 | C36H62O9 | H | H | |
| Oleanolic acid type | ||||
| Ro | C48H76O19 | GlcUA-Glc | Glc | |
| Ocotillol-type | ||||
| P-F11 | C42H72O14 | Glc-Rha | None |
| Treatment | Study Design | Diseases | Sample Size | Treatment Dosage and Rote | Outcomes | References |
|---|---|---|---|---|---|---|
| Sun ginseng-135 (ginsenoside complex) | Randomized, open-label trial | AD | 40 | From 1.5 g/day up to 4.5 g/day; oral administration | MMSE scores and ADAS scores were improved | Heo et al. [11] |
| Korean red ginseng (KRG) (containing 8.54% of ginsenosides) | Randomized, open-label trial | AD | 31 | 4.5 g/day or 9 g/day; oral administration | CDR and ADAS scores were significantly improved | Heo et al. [44] |
| Panax ginseng powder (contains total 8.19% of ginsenosides) | Randomized controlled trial | AD | 97 | 4.5 g/day or 9 g/day; oral administration | ADAS and the MMSE score show improvements during ginseng treatment | Lee et al. [45] |
| KRG | Randomized, double-blind, placebo-controlled trail | Healthy individuals | 51 | 1000 mg/d (500 mg/capsule × 2 capsules). oral administration | Gray matter volume in the left parahippocampal gyrus and the composite score of combined cognitive function were significantly increased | Namgung et al. [46] |
| HT1001 (consists of a mixture of important ginsenosides) | Randomized controlled trial | Schizophrenia | 64 | 100 mg/day; oral administration | Visual working memory was significantly improved, extrapyramidal symptoms were significantly reduced | Chen et al. [48] |
| KRG (contained major ginsenosides) | Double-blind, randomized, placebo-controlled trial | Participants with high-stress occupations | 63 | 500 mg of KRG powder per capsule; oral administration | Triglyceride levels were significantly increased, epinephrine level was decreased | Beak et al. [49] |
| Cereboost™ (10.65% ginsenosides ) | Randomized, double-blind, placebo-controlled crossover | Healthy young adults | 32 | 100, 200, or 400 mg/day; oral administration | Working memory was improved | Scholey et al. [50] |
| G115 (ginsenoside complex) | Double-blind, placebo-controlled, balanced crossover | Healthy young adults | 27 | 200 mg/day; oral administration | Enhancing cognitive performance | Reay et al. [51] |
| Compound | Diseases Model | Species | Administration Method and Duration | Treatment Dosage | Results | References |
|---|---|---|---|---|---|---|
| Rb1 | AD | SAMP8 mice | Intragastric administration (8 weeks) | 30 and 60 µmol/kg | Repaired neuronal cells loss and inhibited the activation of astrocyte and microglia in hippocampus | Yang et al. [53] |
| AD | Aβ1–40-induced rat | Intraperitoneal injection (2 weeks) | 12.5, 25, and 50 mg/kg | Inhibited the levels of pro-apoptosis mediators and improved the levels of anti-apoptosis mediators | Wang et al. [62] | |
| AD | Aβ1–40-induced rat | Intragastric administration (2 weeks) | 12.5 mg/kg/d, 25.0 mg/kg/ d, and 50.0 mg/kg/d) | Altered the amyloidogenic process of APP into non-amyloidogenic process | Lin et al. [63] | |
| AD | ICR mice | Intragastric administration (4 months) | 20 mg/kg/day | Protected against Al-induced toxicity | Zhao et al. [64] | |
| PD | C57BL/6 mice | Intraperitoneal injection | 10 mg/kg | Enhanced GABA release | Liu et al. [66] | |
| PD | PC12 cells | - | 50 or 150 μM | Reduced the cytotoxicity of MPTP | Rudakewich et al. [67] | |
| Focal cerebral ischemia | Sprague–Dawley (SD) rats | Intraperitoneal injection | 100 mg/kg, 50 mg/kg, 25 mg/kg | Increased the expressions of P-Akt, P-mTOR, and reducedP-PTEN and caspase-3 | Yan et al. [68] | |
| Diabetes | Primary hippocampal neuronal cells | - | 1 µM | inhibited GSK3β-mediated CHOP induction | Liu et al. [69] | |
| Diabetes | C57BL/6N male mice | Intragastric administration (4 weeks) | 30 mg/kg | Relieved glucose intolerance, inhibited Cdk5/p35 | Yang et al. [70] | |
| Rg1 | AD | SAMP8 mice | Intragastric administration (8 weeks) | 30 and 60 µmol/kg | Repaired neuronal cells loss and inhibited the activation of astrocyte and microglia in hippocampus | Yang et al. [53] |
| AD | SAMP8 mice | Intragastric administration (3 months) | 2.5, 5.0, and 10 mg/kg | Attenuated hippocampal Aβ content | Shi et al. [54] | |
| AD | SAMP8 mice | Intragastric administration | 7.5 mg/kg/day | Suppressed neuron cell apoptosis | Shi et al. [55] | |
| AD | N2a-APP695 cells | - | 2.5 μM | Decreased the levels of Aβ1–40 and Aβ1–42 | Chen et al. [56] | |
| AD | SD rats | Intracerebroventricular injection (25 days) | 20 mg/kg | Attenuated Aβ formation | Song et al [57] | |
| AD | APP/PS1 mice | Intragastric administration (6 to 9 months) | 5 mg/kg | Decreased the p-Tau level, amyloid precursor protein (APP) expression, and Aβ generation | Zhang et al. [59] | |
| AD | Male conventional tree shrews | Intraperitoneally intragastric administration (8 weeks) | 7.5, 15, and 30 mg/kg | Changed the abundance of gut microbiota | Wang et al. [60] | |
| AD | Tree shrews | Intracerebroventricular injection (6 weeks) | 30 mg/kg/day | Altered the microbiota abundance, affected the expression of apoptosis proteins | Guo et al. [61] | |
| PD | PC12 cells | - | 50 or 150 μM | Reduced the cytotoxicity of MPTP | Rudakewich et al. [67] | |
| Alcohol-induced psychomotor and cognitive deficits | ICR mice | Intragastric administration (14 days) | 3 mg/kg, 6 mg/kg, and 12 mg/kg | Meliorated repeated alcohol-induced cognitive deficits | Huang et al. [71] | |
| Aging | SD rats | Subcutaneous injection (42 days) | 20 mg/kg·d | Improved cognitive ability, protected NSCs/NPCs, and promoted neurogenesis | Zhu et al. [72] | |
| Aging | Kunming mice | Intraperitoneal injection (42 days) | 10, 20 mg·kg−1 | Inhibited apoptosis | Zhong et al. [73] | |
| Rd | Transient forebrain ischemia | Primary neurons of SD rats | - | 10 μM | Attenuated Tau protein phosphorylation | Zhang et al. [58] |
| Chronic cerebral hypoperfusion | C57BL/6J mice | Intraperitoneal injection (21 days) | 10 or 30 mg/kg | Upregulated BDNF and increased neuron survival | Wan et al. [74] | |
| Chronic restraint stress | C57BL/6J mice | Intragastric administration (7 days) | 10, 20, or 40 mg/kg | Mitigated oxidative stress and inflammation | Wang et al. [75] | |
| Compound K | AD | Mouse hippocampal HT22 cells/male C57BL/6J mice | Intragastric administration (2 weeks) | 1, 5, 10 mg/kg | Reduced reactive oxygen species- | Seo et al. [65] |
| Rg5 | Thermal stress | HT22 cells | - | 20 mg/ mL and 40 mg/mL | Prevented apoptosis | Choi et al. [76] |
| Re | Diabetes | SD rats | Intragastric administration (8 weeks) | 40 mg/kg | Attenuated diabetes-associated cognitive decline | Liu et al. [77] |
| Compounds | Herbal Plants | Diseases | Main Findings | References |
|---|---|---|---|---|
| Astragaloside IV | Astmgali Radix. | AD | Inhibited of microglial activation | Chen et al. [93] |
| EGb 761 | Ginkgo biloba | AD | Neuroprotective effect | Mazza et al. [94] |
| Puerarin | Pueraria lobata | AD | Reduced impairment of iron metabolism | Yu et al. [95] |
| EGB761 and HBO | Ginkgo biloba | AD | Reduced cell toxicity and oxidative stress | Tian et al. [96] |
| 4-O-methylhonokiol | Magnolia officinalis | AD | Attenuated β-amyloid-induced memory impairment | Lee et al. [97] |
| xiecaoside A–C and xiecaoline A, xiecaoside D, xiecaoside E | Valeriana amurensis | AD | Protected against Aβ-induced toxicity | Wang et al. [98] |
| Amurensin G, r-2-viniferin and trans-ε-viniferin | Vitis amurensis | AD | Protected against amyloid β protein (25–35)-induced neurotoxicity | Jeong et al. [99] |
| Dendrobium nobile Lindl. alkaloid | Dendrobium nobile Lindl | AD | Suppressed NLRP3-mediated pyroptosis | Li et al. [100] |
| Spinosin | Zizyphus jujuba var. spinosa | AD | Ameliorated Aβ1–42 oligomer-induced memory impairment | Ko et al. [101] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, H.; Xue, M.; Deng, H.; Cheng, S.; Hu, Y.; Zhou, C. Ginsenoside and Its Therapeutic Potential for Cognitive Impairment. Biomolecules 2022, 12, 1310. https://doi.org/10.3390/biom12091310
Feng H, Xue M, Deng H, Cheng S, Hu Y, Zhou C. Ginsenoside and Its Therapeutic Potential for Cognitive Impairment. Biomolecules. 2022; 12(9):1310. https://doi.org/10.3390/biom12091310
Chicago/Turabian StyleFeng, Hui, Mei Xue, Hao Deng, Shiqi Cheng, Yue Hu, and Chunxiang Zhou. 2022. "Ginsenoside and Its Therapeutic Potential for Cognitive Impairment" Biomolecules 12, no. 9: 1310. https://doi.org/10.3390/biom12091310
APA StyleFeng, H., Xue, M., Deng, H., Cheng, S., Hu, Y., & Zhou, C. (2022). Ginsenoside and Its Therapeutic Potential for Cognitive Impairment. Biomolecules, 12(9), 1310. https://doi.org/10.3390/biom12091310