
Roles of Two-Component Signal Transduction Systems in Shigella Virulence

 , ,
, ,  ,
,
Abstract
:1. Introduction
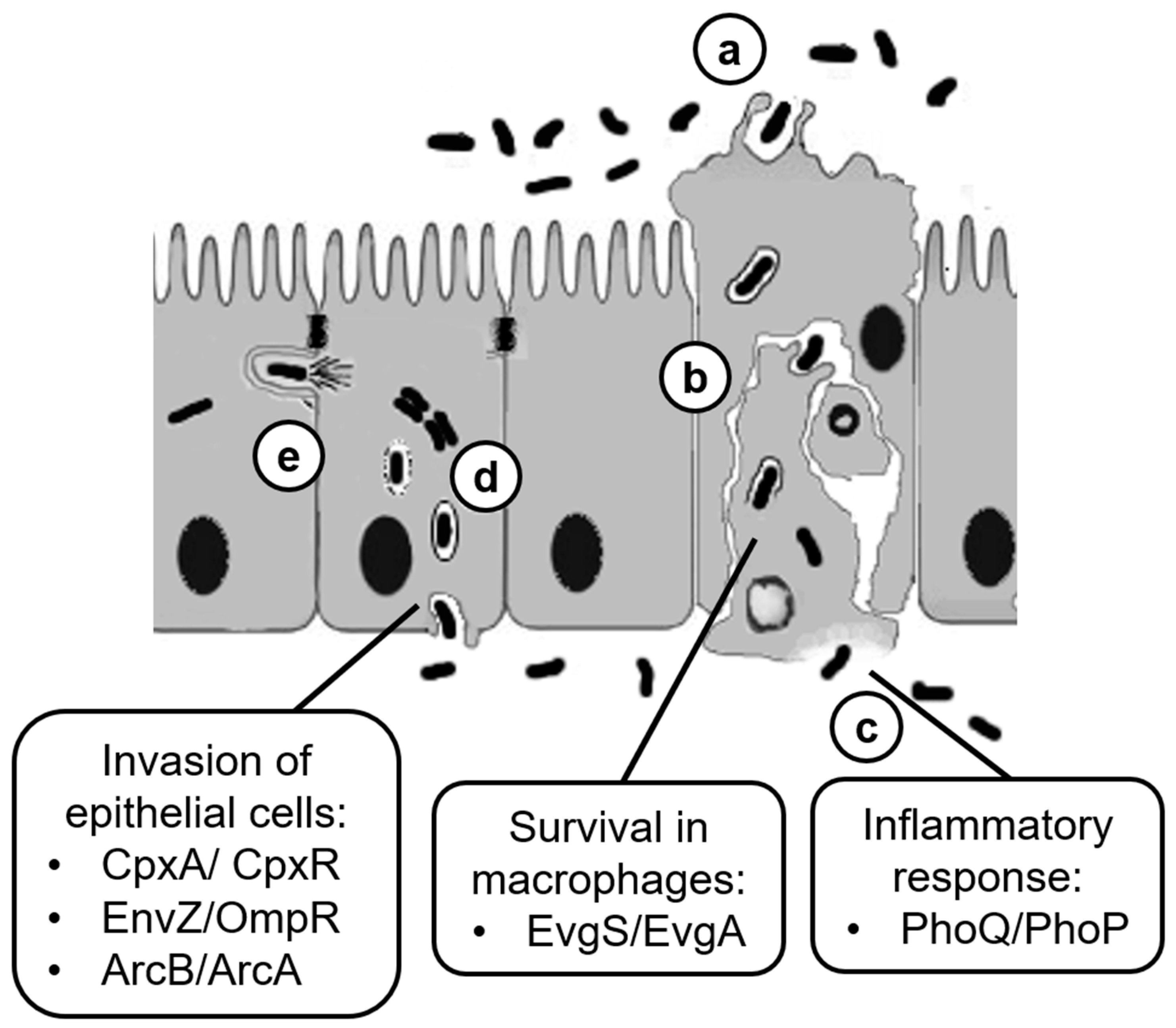
2. The Pathogenicity Process in Shigella
3. Regulation of Shigella Plasmid Virulence Genes
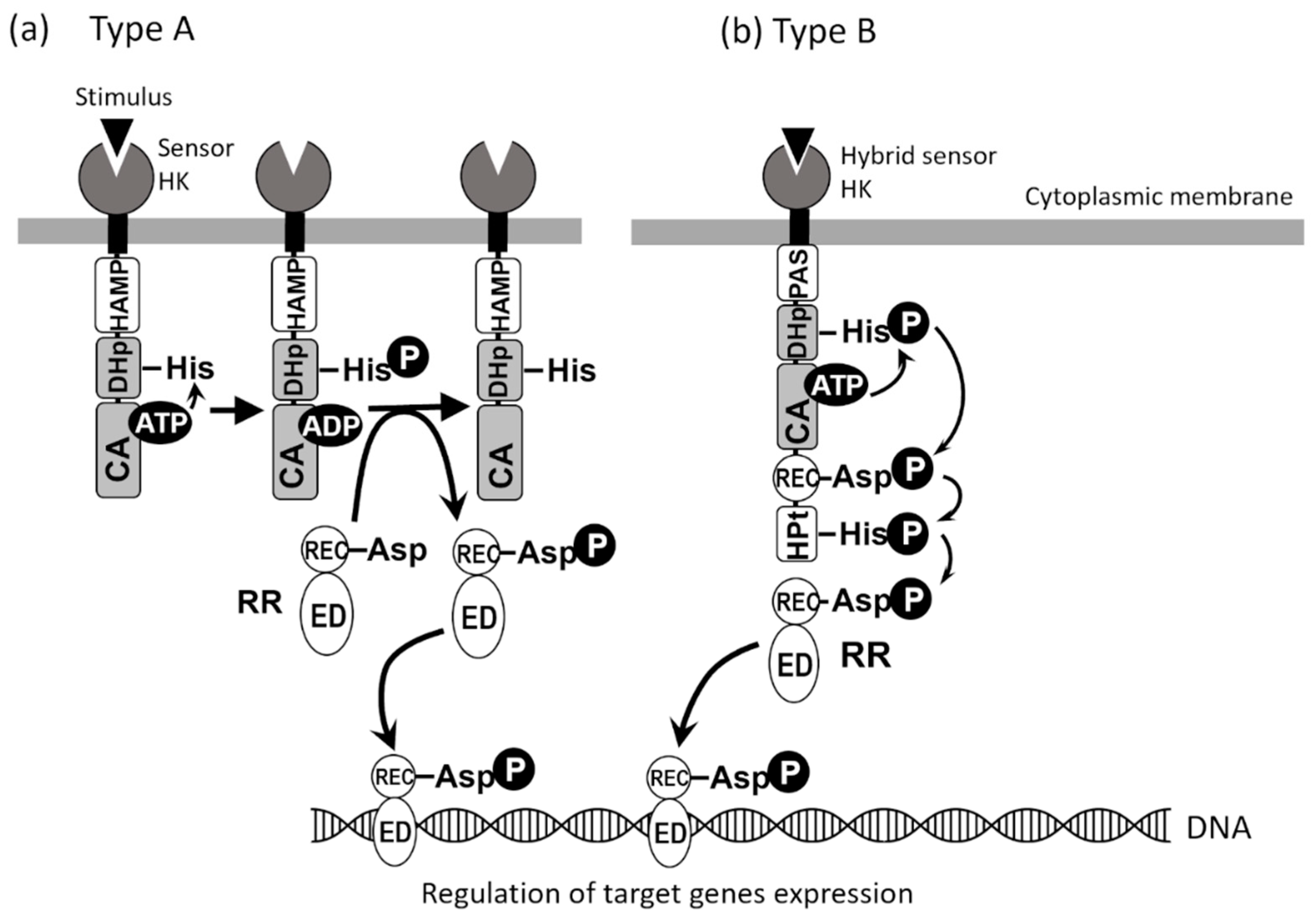
4. Two-Component Signal Transduction Systems of Shigella
5. EnvZ/OmpR and the Invasion of Epithelial Cells in Response to Osmolarity
6. CpxA/CpxR and the Regulation of vir Genes in Response to pH
7. ArcB/AcrA and the Activation of Iron Transport Systems
8. The Role of PhoQ/PhoP during Later Stage of Shigella Infection
9. Role of EvgS/EvgA during Intracellular Life of Shigella
10. Molecular Mechanism of TCS Network
11. Prospects: Drug Discovery Targeting TCS Network
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Beier, D.; Gross, R. Regulation of bacterial virulence by two-component systems. Curr. Opin. Microbiol. 2006, 9, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Calva, E.; Oropeza, R. Two-component signal transduction systems, environmental signals, and virulence. Microb. Ecol. 2006, 51, 166–176. [Google Scholar] [CrossRef]
- Jacob-Dubuisson, F.; Mechaly, A.; Betton, J.M.; Antoine, R. Structural insights into the signalling mechanisms of two-component systems. Nat. Rev. Microbiol. 2018, 16, 585–593. [Google Scholar] [CrossRef]
- Buschiazzo, A.; Trajtenberg, F. Two-Component Sensing and Regulation: How Do Histidine Kinases Talk with Response Regulators at the Molecular Level? Annu. Rev. Microbiol. 2019, 73, 507–528. [Google Scholar] [CrossRef]
- Anderson, M.; Sansonetti, P.J.; Marteyn, B.S. Shigella Diversity and Changing Landscape: Insights for the Twenty-First Century. Front. Cell. Infect. Microbiol. 2016, 6, 45. [Google Scholar] [CrossRef]
- The, H.C.; Thanh, D.P.; Holt, K.E.; Thomson, N.R.; Baker, S. The genomic signatures of Shigella evolution, adaptation and geographical spread. Nat. Rev. Microbiol. 2016, 14, 235–250. [Google Scholar] [CrossRef]
- Pupo, G.M.; Lan, R.; Reeves, P.R. Multiple independent origins of Shigella clones of Escherichia coli and convergent evolution of many of their characteristics. Proc. Natl. Acad. Sci. USA 2000, 97, 10567–10572. [Google Scholar] [CrossRef]
- Pasqua, M.; Michelacci, V.; Di Martino, M.L.; Tozzoli, R.; Grossi, M.; Colonna, B.; Morabito, S.; Prosseda, G. The intriguing evolutionary journey of enteroinvasive E. coli (EIEC) toward pathogenicity. Front. Microbiol. 2017, 8, 2390. [Google Scholar] [CrossRef] [PubMed]
- Sansonetti, P.J.; Kopecko, D.J.; Formal, S.B. Involvement of a plasmid in the invasive ability of Shigella flexneri. Infect. Immun. 1982, 35, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Bliven, K.A.; Maurelli, A.T. Antivirulence genes: Insights into pathogen evolution through gene loss. Infect. Immun. 2012, 80, 4061–4070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prosseda, G.; Di Martino, M.L.; Campilongo, R.; Fioravanti, R.; Micheli, G.; Casalino, M.; Colonna, B. Shedding of genes that interfere with the pathogenic lifestyle: The Shigella model. Res. Microbiol. 2012, 163, 399–406. [Google Scholar] [CrossRef]
- Leuzzi, A.; Grossi, M.; Di Martino, M.L.; Pasqua, M.; Micheli, G.; Colonna, B.; Prosseda, G. Role of the SRRz/Rz1 lambdoid lysis cassette in the pathoadaptive evolution of Shigella. Int. J. Med. Microbiol. 2017, 307, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Schnupf, P.; Sansonetti, P.J. Shigella Pathogenesis: New Insights through Advanced Methodologies. Microbiol. Spectr. 2019, 7, 7.2.28. [Google Scholar] [CrossRef] [PubMed]
- Ashida, H.; Suzuki, T.; Sasakawa, C. Shigella infection and host cell death: A double-edged sword for the host and pathogen survival. Curr. Opin. Microbiol. 2021, 59, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mattock, E.; Blocker, A.J. How Do the Virulence Factors of Shigella Work Together to Cause Disease? Front. Cell. Infect. Microbiol. 2017, 7, 64. [Google Scholar] [CrossRef] [PubMed]
- Parsot, C. Shigella spp. and enteroinvasive Escherichia coli pathogenicity factors. FEMS Microbiol. Lett. 2005, 252, 11–18. [Google Scholar] [CrossRef]
- Killackey, S.A.; Sorbara, M.T.; Girardin, S.E. Cellular Aspects of Shigella Pathogenesis: Focus on the Manipulation of Host Cell Processes. Front. Cell. Infect. Microbiol. 2016, 6, 38. [Google Scholar] [CrossRef]
- Buchrieser, C.; Glaser, P.; Rusniok, C.; Nedjari, H.; D’Hauteville, H.; Kunst, F.; Sansonetti, P.; Parsot, C. The virulence plasmid pWR100 and the repertoire of proteins secreted by the type III secretion apparatus of Shigella flexneri. Mol. Microbiol. 2000, 38, 760–771. [Google Scholar] [CrossRef]
- Di Martino, M.L.; Falconi, M.; Micheli, G.; Colonna, B.; Prosseda, G. The multifaceted activity of the VirF regulatory protein in the Shigella Lifestyle. Front. Mol. Biosci. 2016, 3, 61. [Google Scholar] [CrossRef]
- Falconi, M.; Colonna, B.; Prosseda, G.; Micheli, G.; Gualerzi, C.O. Thermoregulation of Shigella and Escherichia coli EIEC pathogenicity. A temperature-dependent structural transition of DNA modulates accessibility of virF promoter to transcriptional repressor H-NS. EMBO J. 1998, 17, 7033–7043. [Google Scholar] [CrossRef] [Green Version]
- Bernardini, M.L.; Mounier, J.; D’Hauteville, H.; Coquis-Rondon, M.; Sansonetti, P.J. Identification of icsA, a plasmid locus of Shigella flexneri that governs bacterial intra- and intercellular spread through interaction with F-actin. Proc. Natl. Acad. Sci. USA 1989, 86, 3867–3871. [Google Scholar] [CrossRef] [PubMed]
- Tobe, T.; Yoshikawa, M.; Mizuno, T.; Sasakawa, C. Transcriptional control of the invasion regulatory gene virB of Shigella flexneri: Activation by virF and repression by H-NS. J. Bacteriol. 1993, 175, 6142–6149. [Google Scholar] [CrossRef]
- Prosseda, G.; Falconi, M.; Giangrossi, M.; Gualerzi, C.O.; Micheli, G.; Colonna, B. The virF promoter in Shigella: More than just a curved DNA stretch. Mol. Microbiol. 2004, 51, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Tran, C.N.; Giangrossi, M.; Prosseda, G.; Brandi, A.; Di Martino, M.L.; Colonna, B.; Falconi, M. A multifactor regulatory circuit involving H-NS, VirF and an antisense RNA modulates transcription of the virulence gene icsA of Shigella flexneri. Nucleic Acids Res. 2011, 39, 8122–8134. [Google Scholar] [CrossRef]
- Falconi, M.; Prosseda, G.; Giangrossi, M.; Beghetto, E.; Colonna, B. Involvement of FIS in the H-NS-mediated regulation of virF gene of Shigella and enteroinvasive Escherichia coli. Mol. Microbiol. 2001, 42, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Porter, M.E.; Dorman, C.J. Positive regulation of Shigella flexneri virulence genes by integration host factor. J. Bacteriol. 1997, 179, 6537–6550. [Google Scholar] [CrossRef]
- Giangrossi, M.; Prosseda, G.; Tran, C.N.; Brandi, A.; Colonna, B.; Falconi, M. A novel antisense RNA regulates at transcriptional level the virulence gene icsA of Shigella flexneri. Nucleic Acids Res. 2010, 38, 3362–3375. [Google Scholar] [CrossRef]
- Broach, W.H.; Egan, N.; Wing, H.J.; Payne, S.M.; Murphy, E.R. VirF-Independent Regulation of Shigella virB Transcription is Mediated by the Small RNA RyhB. PLoS ONE 2012, 7, e38592. [Google Scholar] [CrossRef]
- Durand, J.M.B.; Dagberg, B.; Uhlin, B.E.; Björk, G.R. Transfer RNA modification, temperature and DNA superhelicity have a common target in the regulatory network of the virulence of Shigella flexneri: The expression of the virF gene. Mol. Microbiol. 2000, 35, 924–935. [Google Scholar] [CrossRef]
- Hurt, J.K.; Olgen, S.; Garcia, G.A. Site-specific modification of Shigella flexneri virF mRNA by tRNA-guanine transglycosylase in vitro. Nucleic Acids Res. 2007, 35, 4905–4913. [Google Scholar] [CrossRef]
- Di Martino, M.L.; Romilly, C.; Wagner, E.G.H.; Colonna, B.; Prosseda, G. One gene and two proteins: A leaderless mRNA supports the translation of a shorter form of the Shigella VirF regulator. mBio 2016, 7, e01860-16. [Google Scholar] [CrossRef]
- Skovajsová, E.; Colonna, B.; Prosseda, G.; Sellin, M.E.; Letizia, M.; Martino, D. The VirF21:VirF30 protein ratio is affected by temperature and impacts Shigella flexneri host cell invasion. FEMS Microbiol. Lett. 2022, 369, fnac043. [Google Scholar] [CrossRef] [PubMed]
- Tobe, T. The roles of two-component systems in virulence of pathogenic Escherichia coli and Shigella spp. Adv. Exp. Med. Biol. 2008, 631, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, Y.; Eguchi, Y.; Watanabe, T.; Okamoto, S.; Doi, A.; Utsumi, R. Two-component signal transduction as potential drug targets in pathogenic bacteria. Curr. Opin. Microbiol. 2010, 13, 232–239. [Google Scholar] [CrossRef]
- Wanner, B.L. Gene regulation by phosphate in enteric bacteria. J. Cell. Biochem. 1993, 51, 47–54. [Google Scholar] [CrossRef]
- Heermann, R.; Jung, K. Stimulus Perception and Signaling in Histidine Kinases. In Bacterial Signaling; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; pp. 135–161. [Google Scholar] [CrossRef]
- Ishii, E.; Eguchi, Y. Diversity in Sensing and Signaling of Bacterial Sensor Histidine Kinases. Biomolecules 2021, 11, 1524. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, Y.; Utsumi, R. Alkali metals in addition to acidic pH activate the EvgS histidine kinase sensor in Escherichia coli. J. Bacteriol. 2014, 196, 3140–3149. [Google Scholar] [CrossRef]
- Leblanc, S.K.D.; Oates, C.W.; Raivio, T.L. Characterization of the induction and cellular role of the BaeSR two-component envelope stress response of Escherichia coli. J. Bacteriol. 2011, 193, 3367–3375. [Google Scholar] [CrossRef]
- Chavez, R.G.; Alvarez, A.F.; Romeo, T.; Georgellis, D. The physiological stimulus for the BarA sensor kinase. J. Bacteriol. 2010, 192, 2009–2012. [Google Scholar] [CrossRef] [Green Version]
- Boon, N.; Kaur, M.; Aziz, A.; Bradnick, M.; Shibayama, K.; Eguchi, Y.; Lund, P.A. The Signaling Molecule Indole Inhibits Induction of the AR2 Acid Resistance System in Escherichia coli. Front. Microbiol. 2020, 11, 474. [Google Scholar] [CrossRef]
- Jiang, P.; Pioszak, A.; Atkinson, M.R.; Peliska, J.A.; Ninfa, A.J. Chapter 7—New Insights into the Mechanism of the Kinase and Phosphatase Activities of Escherichia coli NRII (NtrB) and Their Regulation by the PII Protein. In Histidine Kinases in Signal Transduction; Elsevier: Amsterdam, The Netherlands, 2003; pp. 143–164. [Google Scholar] [CrossRef]
- Behr, S.; Kristoficova, I.; Witting, M.; Breland, E.J.; Eberly, A.R.; Sachs, C.; Schmitt-Kopplin, P.; Hadjifrangiskou, M.; Jung, K. Identification of a High-Affinity Pyruvate Receptor in Escherichia coli. Sci. Rep. 2017, 7, 1388. [Google Scholar] [CrossRef] [PubMed]
- Fried, L.; Behr, S.; Jung, K. Identification of a target gene and activating stimulus for the YpdA/YpdB histidine kinase/response regulator system in Escherichia coli. J. Bacteriol. 2013, 195, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.H.; Ferrières, L.; Clarke, D.J. The role of the Rcs phosphorelay in Enterobacteriaceae. Res. Microbiol. 2006, 157, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Tomomori, C.; Kurokawa, H.; Ikura, M. Chapter 2—The Histidine Kinase Family: Structures of Essential Building Blocks. In Histidine Kinases in Signal Transduction; Elsevier: Amsterdam, The Netherlands, 2003; pp. 11–24. [Google Scholar] [CrossRef]
- Kinoshita-Kikuta, E.; Kinoshita, E.; Eguchi, Y.; Yanagihara, S.; Edahiro, K.; Inoue, Y.; Taniguchi, M.; Yoshida, M.; Yamamoto, K.; Takahashi, H.; et al. Functional Characterization of the Receiver Domain for Phosphorelay Control in Hybrid Sensor Kinases. PLoS ONE 2015, 10, e0132598. [Google Scholar] [CrossRef] [PubMed]
- Barakat, M.; Ortet, P.; Whitworth, D.E. P2CS: A database of prokaryotic two-component systems. Nucleic Acids Res. 2011, 39, D771–D776. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, S.I.; Watanabe, H. Involvement of cpxA, a sensor of a two-component regulatory system, in the pH-dependent regulation of expression of Shigella sonnei virF gene. J. Bacteriol. 1995, 177, 5062–5069. [Google Scholar] [CrossRef]
- Nakayama, S.I.; Watanabe, H. Identification of cpxR as a positive regulator essential for expression of the Shigella sonnei virF gene. J. Bacteriol. 1998, 180, 3522–3528. [Google Scholar] [CrossRef] [Green Version]
- Mitobe, J.; Arakawa, E.; Watanabe, H. A sensor of the two-component system CpxA affects expression of the type III secretion system through posttranscriptional processing of InvE. J. Bacteriol. 2005, 187, 107–113. [Google Scholar] [CrossRef]
- Bernardini, M.L.; Fontaine, A.; Sansonetti, P.J. The two-component regulatory system ompR-envZ controls the virulence of Shigella flexneri. J. Bacteriol. 1990, 172, 6274–6281. [Google Scholar] [CrossRef]
- Bernardini, M.L.; Sanna, M.G.; Fontaine, A.; Sansonetti, P.J. OmpC is involved in invasion of epithelial cells by Shigella flexneri. Infect. Immun. 1993, 61, 3625–3635. [Google Scholar] [CrossRef]
- Boulette, M.L.; Payne, S.M. Anaerobic regulation of Shigella flexneri virulence: ArcA regulates fur and iron acquisition genes. J. Bacteriol. 2007, 189, 6957–6967. [Google Scholar] [CrossRef] [PubMed]
- Pasqua, M.; Grossi, M.; Scinicariello, S.; Aussel, L.; Barras, F.; Colonna, B.; Prosseda, G. The MFS efflux pump EmrKY contributes to the survival of Shigella within macrophages. Sci. Rep. 2019, 9, 2906. [Google Scholar] [CrossRef]
- Monsieurs, P.; De Keersmaecker, S.; Navarre, W.W.; Bader, M.W.; De Smet, F.; McClelland, M.; Fang, F.C.; De Moor, B.; Vanderleyden, J.; Marchal, K. Comparison of the PhoPQ regulon in Escherichia coli and Salmonella typhimurium. J. Mol. Evol. 2005, 60, 462–474. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Cai, X.; Chen, M.; Ye, L.; Wu, Y.; Wang, X.; Lv, Z.; Shang, Y.; Qu, D. Virulence and stress responses of Shigella flexneri regulated by PhoP/PhoQ. Front. Microbiol. 2018, 8, 2689. [Google Scholar] [CrossRef] [PubMed]
- Slauch, J.M.; Silhavy, T.J. Genetic analysis of the switch that controls porin gene expression in Escherichia coli K-12. J. Mol. Biol. 1989, 210, 281–292. [Google Scholar] [CrossRef]
- Hall, M.N.; Silhavy, T.J. Genetic analysis of the ompB locus in Escherichia coli K-12. J. Mol. Biol. 1981, 151, 1–15. [Google Scholar] [CrossRef]
- Cai, S.J.; Inouye, M. EnvZ-OmpR Interaction and Osmoregulation in Escherichia coli. J. Biol. Chem. 2002, 277, 24155–24161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenney, L.J.; Anand, G.S. EnvZ/OmpR Two-Component Signaling: An Archetype System That Can Function Noncanonically. EcoSal Plus 2020, 9. [Google Scholar] [CrossRef]
- Perkins, T.T.; Davies, M.R.; Klemm, E.J.; Rowley, G.; Wileman, T.; James, K.; Keane, T.; Maskell, D.; Hinton, J.C.D.; Dougan, G.; et al. ChIP-seq and transcriptome analysis of the OmpR regulon of Salmonella enterica serovars Typhi and Typhimurium reveals accessory genes implicated in host colonization. Mol. Microbiol. 2013, 87, 526–538. [Google Scholar] [CrossRef]
- Gerken, H.; Vuong, P.; Soparkar, K.; Misra, R. Roles of the ENVZ/OMPR two-component system and porins in iron acquisition in Escherichia coli. mBio 2020, 11, e01192-20. [Google Scholar] [CrossRef]
- Quinn, H.J.; Cameron, A.D.S.; Dorman, C.J. Bacterial Regulon Evolution: Distinct Responses and Roles for the Identical OmpR Proteins of Salmonella typhimurium and Escherichia coli in the Acid Stress Response. PLoS Genet. 2014, 10, e1004215. [Google Scholar] [CrossRef]
- Raivio, T.L. Everything old is new again: An update on current research on the Cpx envelope stress response. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 1529–1541. [Google Scholar] [CrossRef] [PubMed]
- Isaac, D.D.; Pinkner, J.S.; Hultgren, S.J.; Silhavy, T.J. The extracytoplasmic adaptor protein CpxP is degraded with substrate by DegP. Proc. Natl. Acad. Sci. USA 2005, 102, 17775–17779. [Google Scholar] [CrossRef]
- Tschauner, K.; Hörnschemeyer, P.; Müller, V.S.; Hunke, S. Dynamic Interaction between the CpxA Sensor Kinase and the Periplasmic Accessory Protein CpxP Mediates Signal Recognition in E. coli. PLoS ONE 2014, 9, e107383. [Google Scholar] [CrossRef]
- Delhaye, A.; Laloux, G.; Collet, J.F. The lipoprotein NlpE is a cpx sensor that serves as a sentinel for protein sorting and folding defects in the Escherichia coli envelope. J. Bacteriol. 2019, 209, e00611-18. [Google Scholar] [CrossRef]
- Gubbins, M.J.; Lau, I.; Will, W.R.; Manchak, J.M.; Raivio, T.L.; Frost, L.S. The positive regulator, TraJ, of the Escherichia coli F plasmid is unstable in a cpxA* background. J. Bacteriol. 2002, 184, 5781–5788. [Google Scholar] [CrossRef]
- Vogt, S.L.; Raivio, T.L. Just scratching the surface: An expanding view of the Cpx envelope stress response. FEMS Microbiol. Lett. 2012, 326, 2–11. [Google Scholar] [CrossRef]
- Nakayama, S.I.; Kushiro, A.; Asahara, T.; Tanaka, R.I.; Hu, L.; Kopecko, D.J.; Watanabe, H. Activation of hilA expression at low pH requires the signal sensor CpxA, but not the cognate response regulator CpxR, in Salmonella enterica serovar Typhimurium. Microbiology 2003, 149, 2809–2817. [Google Scholar] [CrossRef] [PubMed]
- Hung, D.L.; Raivio, T.L.; Jones, C.H.; Silhavy, T.J.; Hultgren, S.J. Cpx signaling pathway monitors biogenesis and affects assembly and expression of P pili. EMBO J. 2001, 20, 1508–1518. [Google Scholar] [CrossRef] [PubMed]
- Tanner, J.R.; Li, L.; Faucher, S.P.; Brassinga, A.K.C. The CpxRA two-component system contributes to Legionella pneumophila virulence. Mol. Microbiol. 2016, 100, 1017–1038. [Google Scholar] [CrossRef]
- Nevesinjac, A.Z.; Raivio, T.L. The Cpx envelope stress response affects expression of the type IV bundle-forming pili of enteropathogenic Escherichia coli. J. Bacteriol. 2005, 187, 672–686. [Google Scholar] [CrossRef] [PubMed]
- Vogt, S.L.; Scholz, R.; Peng, Y.; Guest, R.L.; Scott, N.E.; Woodward, S.E.; Foster, L.J.; Raivio, T.L.; Finlay, B.B. Characterization of the Citrobacter rodentium Cpx regulon and its role in host infection. Mol. Microbiol. 2019, 111, 700–716. [Google Scholar] [CrossRef] [PubMed]
- Marteyn, B.; Gazi, A.; Sansonetti, P. Shigella: A model of virulence regulation in vivo. Gut Microbes. 2012, 3, 104–120. [Google Scholar] [CrossRef] [PubMed]
- Malpica, R.; Peña Sandoval, G.R.; Rodríguez, C.; Franco, B.; Georgellis, D. Signaling by the Arc Two-Component System Provides a Link Between the Redox State of the Quinone Pool and Gene Expression. Antioxid. Redox Signal. 2006, 8, 781–795. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.N.; Anderson, M.T.; Bachman, M.A.; Mobley, H.L.T. The ArcAB Two-Component System: Function in Metabolism, Redox Control, and Infection. Microbiol. Mol. Biol. Rev. 2022, 86, e00110-21. [Google Scholar] [CrossRef]
- Salmon, K.A.; Hung, S.P.; Steffen, N.R.; Krupp, R.; Baldi, P.; Hatfield, G.W.; Gunsalus, R.P. Global Gene Expression Profiling in Escherichia coli K12: EFFECTS OF OXYGEN AVAILABILITY AND ArcA. J. Biol. Chem. 2005, 280, 15084–15096. [Google Scholar] [CrossRef]
- Kwon, O.; Georgellis, D.; Lin, E.C.C. Phosphorelay as the sole physiological route of signal transmission by the arc two-component system of Escherichia coli. J. Bacteriol. 2000, 182, 3858–3862. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, A.F.; Georgellis, D. In Vitro and In Vivo Analysis of the ArcB/A Redox Signaling Pathway. Methods Enzymol. 2010, 471, 205–228. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Antonio, A.; Collado-Vides, J. Identifying global regulators in transcriptional regulatory networks in bacteria. Curr. Opin. Microbiol. 2003, 6, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Green, J.; Paget, M.S. Bacterial redox sensors. Nat. Rev. Microbiol. 2004, 2, 954–966. [Google Scholar] [CrossRef] [PubMed]
- Wyckoff, E.E.; Boulette, M.L.; Payne, S.M. Genetics and environmental regulation of Shigella iron transport systems. Biometals 2009, 22, 43–51. [Google Scholar] [CrossRef]
- Mey, A.R.; Gómez-Garzón, C.; Payne, S.M. Iron Transport and Metabolism in Escherichia, Shigella, and Salmonella. EcoSal Plus 2021, 9, eESP-0034-2020. [Google Scholar] [CrossRef]
- Wei, Y.; Murphy, E.R. Shigella Iron Acquisition Systems and their Regulation. Front. Cell. Infect. Microbiol. 2016, 6, 18. [Google Scholar] [CrossRef]
- Groisman, E.A.; Duprey, A.; Choi, J. How the PhoP/PhoQ System Controls Virulence and Mg 2+ Homeostasis: Lessons in Signal Transduction, Pathogenesis, Physiology, and Evolution. Microbiol. Mol. Biol. Rev. 2021, 85, e00176-20. [Google Scholar] [CrossRef]
- García Véscovi, E.; Soncini, F.C.; Groisman, E.A. Mg2+ as an Extracellular Signal: Environmental Regulation of Salmonella Virulence. Cell 1996, 84, 165–174. [Google Scholar] [CrossRef]
- Kato, A.; Tanabe, H.; Utsumi, R. Molecular characterization of the PhoP-PhoQ two-component system in Escherichia coli K-12: Identification of extracellular Mg2+-responsive promoters. J. Bacteriol. 1999, 181, 5516–5520. [Google Scholar] [CrossRef]
- Minagawa, S.; Ogasawara, H.; Kato, A.; Yamamoto, K.; Eguchi, Y.; Oshima, T.; Mori, H.; Ishihama, A.; Utsumi, R. Identification and molecular characterization of the Mg2+ stimulon of Escherichia coli. J. Bacteriol. 2003, 185, 3696–3702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prost, L.R.; Daley, M.E.; Le Sage, V.; Bader, M.W.; Le Moual, H.; Klevit, R.E.; Miller, S.I. Activation of the Bacterial Sensor Kinase PhoQ by Acidic pH. Mol. Cell 2007, 26, 165–174. [Google Scholar] [CrossRef]
- Bader, M.W.; Sanowar, S.; Daley, M.E.; Schneider, A.R.; Cho, U.; Xu, W.; Klevit, R.E.; Le Moual, H.; Miller, S.I. Recognition of Antimicrobial Peptides by a Bacterial Sensor Kinase. Cell 2005, 122, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Jin, F.; Glatter, T.; Sourjik, V. Osmosensing by the bacterial PhoQ/PhoP two-component system. Proc. Natl. Acad. Sci. USA 2017, 114, E10792–E10798. [Google Scholar] [CrossRef]
- Lippa, A.M.; Goulian, M. Perturbation of the oxidizing environment of the periplasm stimulates the PhoQ/PhoP system in Escherichia coli. J. Bacteriol. 2012, 194, 1457–1463. [Google Scholar] [CrossRef] [PubMed]
- Viarengo, G.; Sciara, M.I.; Salazar, M.O.; Kieffer, P.M.; Furlán, R.L.E.; Véscovi, E.G. Unsaturated Long Chain Free Fatty Acids Are Input Signals of the Salmonella enterica PhoP/PhoQ Regulatory System. J. Biol. Chem. 2013, 288, 22346–22358. [Google Scholar] [CrossRef] [PubMed]
- Fields, P.I.; Groisman, E.A.; Heffron, F. A Salmonella Locus that Controls Resistance to Microbicidal Proteins from Phagocytic Cells. Science 1989, 243, 1059–1062. [Google Scholar] [CrossRef] [PubMed]
- Prost, L.R.; Miller, S.I. The Salmonellae PhoQ sensor: Mechanisms of detection of phagosome signals. Cell. Microbiol. 2008, 10, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Moss, J.E.; Fisher, P.E.; Vick, B.; Groisman, E.A.; Zychlinsky, A. The regulatory protein PhoP controls susceptibility to the host inflammatory response in Shigella flexneri. Cell. Microbiol. 2000, 2, 443–452. [Google Scholar] [CrossRef]
- Huang, J.; Li, C.; Song, J.; Velkov, T.; Wang, L.; Zhu, Y.; Li, J. Regulating polymyxin resistance in Gram-negative bacteria: Roles of two-component systems PhoPQ and PmrAB. Future Microbiol. 2020, 15, 445–459. [Google Scholar] [CrossRef]
- Goldman, S.R.; Tu, Y.; Goldberg, M.B. Differential regulation by magnesium of the two MsbB paralogs of Shigella flexneri. J. Bacteriol. 2008, 190, 3526–3537. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.D.; Bell, J.; Clarke, K.; Chandler, R.; Pathak, P.; Xia, Y.; Marshall, R.L.; Weinstock, G.M.; Loman, N.J.; Winn, P.J.; et al. Characterization of mutations in the PAS domain of the EvgS sensor kinase selected by laboratory evolution for acid resistance in Escherichia coli. Mol. Microbiol. 2014, 93, 911–927. [Google Scholar] [CrossRef]
- Roggiani, M.; Yadavalli, S.S.; Goulian, M. Natural variation of a sensor kinase controlling a conserved stress response pathway in Escherichia coli. PLoS Genet. 2017, 13, e1007101. [Google Scholar] [CrossRef]
- Arico, B.; Miller, J.F.; Roy, C.; Stibitz, S.; Monack, D.; Falkow, S.; Gross, R.; Rappuoli, R. Sequences required for expression of Bordetella pertussis virulence factors share homology with prokaryotic signal transduction proteins. Proc. Natl. Acad. Sci. USA 1989, 86, 6671–6675. [Google Scholar] [CrossRef]
- Lai, Y.-C.; Lin, G.-T.; Yang, S.-L.; Chang, H.-Y.; Peng, H.-L. Identification and characterization of KvgAS, a two-component system in Klebsiella pneumoniae CG43. FEMS Microbiol. Lett. 2003, 218, 121–126. [Google Scholar] [CrossRef]
- Utsumi, R.; Katayama, S.; Taniguchi, M.; Horie, T.; Ikeda, M.; Igaki, S.; Nakagawa, H.; Miwa, A.; Tanabe, H.; Noda, M. Newly identified genes involved in the signal transduction of Escherichia coli K-12. Gene 1994, 140, 73–77. [Google Scholar] [CrossRef]
- Kato, A.; Ohnishi, H.; Yamamoto, K.; Furuta, E.; Tanabe, H.; Utsumi, R. Transcription of emrKY is Regulated by the EvgA-EvgS Two-Component System in Escherichia coli K-12. Biosci. Biotechnol. Biochem. 2000, 64, 1203–1209. [Google Scholar] [CrossRef] [PubMed]
- Nishino, K.; Yamaguchi, A. Analysis of a complete library of putative drug transporter genes in Escherichia coli. J. Bacteriol. 2001, 183, 5803–5812. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, Y.; Oshima, T.; Mori, H.; Aono, R.; Yamamoto, K.; Ishihama, A.; Utsumi, R. Transcriptional regulation of drug efflux genes by EvgAS, two-component system in Escherichia coli. Microbiology 2003, 149, 2819–2828. [Google Scholar] [CrossRef] [PubMed]
- Nishino, K.; Yamaguchi, A. EvgA of the two-component signal transduction system modulates production of the YhiUV multidrug transporter in Escherichia coli. J. Bacteriol. 2002, 184, 2319–2323. [Google Scholar] [CrossRef]
- Masuda, N.; Church, G.M. Escherichia coli gene expression responsive to levels of the response regulator EvgA. J. Bacteriol. 2002, 184, 6225–6234. [Google Scholar] [CrossRef] [Green Version]
- Masuda, N.; Church, G.M. Regulatory network of acid resistance genes in Escherichia coli. Mol. Microbiol. 2003, 48, 699–712. [Google Scholar] [CrossRef]
- Itou, J.; Eguchi, Y.; Utsumi, R. Molecular mechanism of transcriptional cascade initiated by the EvgS/EvgA system in Escherichia coli K-12. Biosci. Biotechnol. Biochem. 2009, 73, 870–878. [Google Scholar] [CrossRef]
- Ma, Z.; Gong, S.; Richard, H.; Tucker, D.L.; Conway, T.; Foster, J.W. GadE (YhiE) activates glutamate decarboxylase-dependent acid resistance in Escherichia coli K-12. Mol. Microbiol. 2003, 49, 1309–1320. [Google Scholar] [CrossRef]
- Inada, S.; Okajima, T.; Utsumi, R.; Eguchi, Y. Acid-Sensing Histidine Kinase With a Redox Switch. Front. Microbiol. 2021, 12, 652546. [Google Scholar] [CrossRef] [PubMed]
- Blanco, P.; Hernando-Amado, S.; Reales-Calderon, J.A.; Corona, F.; Lira, F.; Alcalde-Rico, M.; Bernardini, A.; Sanchez, M.B.; Martinez, J.L. Bacterial Multidrug Efflux Pumps: Much More Than Antibiotic Resistance Determinants. Microorganisms 2016, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Pasqua, M.; Grossi, M.; Zennaro, A.; Fanelli, G.; Micheli, G.; Barras, F.; Colonna, B.; Prosseda, G. The varied role of efflux pumps of the mfs family in the interplay of bacteria with animal and plant cells. Microorganisms 2019, 7, 285. [Google Scholar] [CrossRef] [PubMed]
- Pasqua, M.; Bonaccorsi di Patti, M.C.; Fanelli, G.; Utsumi, R.; Eguchi, Y.; Trirocco, R.; Prosseda, G.; Grossi, M.; Colonna, B. Host–Bacterial Pathogen Communication: The Wily Role of the Multidrug Efflux Pumps of the MFS Family. Front. Mol. Biosci. 2021, 8, 709. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Dorsey-Oresto, A.; Malik, M.; Wang, J.Y.; Drlica, K.; Zhao, X.; Lu, T. Escherichia coli genes that reduce the lethal effects of stress. BMC Microbiol. 2010, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Nagakubo, S.; Nishino, K.; Hirata, T.; Yamaguchi, A. The putative response regulator BaeR stimulates multidrug resistance of Escherichia coli via a novel multidrug exporter system, MdtABC. J. Bacteriol. 2002, 184, 4161–4167. [Google Scholar] [CrossRef]
- Demarre, G.L.; Prudent, V.; Schenk, H.; Rousseau, E.; Bringer, M.A.; Barnich, N.; Van Nhieu, G.T.; Rimsky, S.; De Monte, S.; Espéli, O. The Crohn’s disease-Associated Escherichia coli strain LF82 relies on SOS and stringent responses to survive, multiply and tolerate antibiotics within macrophages. PLoS Pathog. 2019, 15, e1008123. [Google Scholar] [CrossRef] [Green Version]
- Fanelli, G.; Pasqua, M.; Colonna, B.; Prosseda, G.; Grossi, M. Expression Profile of Multidrug Resistance Efflux Pumps during Intracellular Life of Adherent-Invasive Escherichia coli Strain LF82. Front. Microbiol. 2020, 11, 1935. [Google Scholar] [CrossRef]
- Sultan, M.; Arya, R.; Kim, K.K.; Sprenger, G.A.; Burkovski, A. Roles of Two-Component Systems in Pseudomonas aeruginosa Virulence. Int. J. Mol. Sci. 2021, 22, 12152. [Google Scholar] [CrossRef]
- Eguchi, Y.; Ishii, E.; Utsumi, R. Molecular Mechanism of Bacterial Two-component Signal Transduction Networks via Connectors. In Two-Component Systems in Bacteria; Gross, H., Beier, D., Eds.; Caister Academic Press: Norfolk, UK, 2012; pp. 149–162. [Google Scholar]
- Gerken, H.; Misra, R. MzrA-EnvZ interactions in the periplasm influence the EnvZ/OmpR two-component regulon. J. Bacteriol. 2010, 192, 6271–6278. [Google Scholar] [CrossRef]
- Motz, M.; Jung, K. The role of polyproline motifs in the histidine kinase EnvZ. PLoS ONE 2018, 13, e0199782. [Google Scholar] [CrossRef]
- Yamamoto, K.; Ishihama, A. Characterization of copper-inducible promoters regulated by CpxA/CpxR in Escherichia coli. Biosci. Biotechnol. Biochem. 2006, 70, 1688–1695. [Google Scholar] [CrossRef] [PubMed]
- Gerken, H.; Charlson, E.S.; Cicirelli, E.M.; Kenney, L.J.; Misra, R. MzrA: A novel modulator of the EnvZ/OmpR two-component regulon. Mol. Microbiol. 2009, 72, 1408–1422. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, Y.; Ishii, E.; Yamane, M.; Utsumi, R. The connector SafA interacts with the multi-sensing domain of PhoQ in Escherichia coli. Mol. Microbiol. 2012, 85, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Ishii, E.; Eguchi, Y.; Utsumi, R. Mechanism of activation of PhoQ/PhoP two-component signal transduction by SafA, an auxiliary protein of PhoQ histidine kinase in Escherichia coli. Biosci. Biotechnol. Biochem. 2013, 77, 814–819. [Google Scholar] [CrossRef]
- Bougdour, A.; Cunning, C.; Baptiste, P.J.; Elliott, T.; Gottesman, S. Multiple pathways for regulation of σS (RpoS) stability in Escherichia coli via the action of multiple anti-adaptors. Mol. Microbiol. 2008, 68, 298–313. [Google Scholar] [CrossRef]
- Villanueva, M.; García, B.; Valle, J.; Rapún, B.; Ruiz De Los Mozos, I.; Solano, C.; Martí, M.; Penadés, J.R.; Toledo-Arana, A.; Lasa, I. Sensory deprivation in Staphylococcus aureus. Nat. Commun. 2018, 9, 523. [Google Scholar] [CrossRef] [Green Version]
- Murret-Labarthe, C.; Kerhoas, M.; Dufresne, K.; Daigle, F. New Roles for Two-Component System Response Regulators of Salmonella enterica Serovar Typhi during Host Cell Interactions. Microorganisms 2020, 8, 722. [Google Scholar] [CrossRef]
- Wilke, K.E.; Carlson, E.E. All signals lost: Bacterial histidine kinase inhibitors that globally block bacterial signaling may offer a new offensive against antibiotic resistance. Sci. Transl. Med. 2013, 5, 203ps12. [Google Scholar] [CrossRef]
- Wilke, K.E.; Francis, S.; Carlson, E.E. Inactivation of multiple bacterial histidine kinases by targeting the ATP-binding domain. ACS Chem. Biol. 2015, 10, 328–335. [Google Scholar] [CrossRef]
- Fihn, C.A.; Carlson, E.E. Targeting a highly conserved domain in bacterial histidine kinases to generate inhibitors with broad spectrum activity. Curr. Opin. Microbiol. 2021, 61, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, Y.; Okajima, T.; Tochio, N.; Inukai, Y.; Shimizu, R.; Ueda, S.; Shinya, S.; Kigawa, T.; Fukamizo, T.; Igarashi, M.; et al. Angucycline antibiotic waldiomycin recognizes common structural motif conserved in bacterial histidine kinases. J. Antibiot. 2017, 70, 251–258. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Escherichia coli K-12 MG1655 | S.dysenteriae 1617 | S.flexneri 2a str.301 | S.boydii Sb227 | S.sonnei 53G | |||
|---|---|---|---|---|---|---|---|
| HK29 ** | RR32 | Stimulus | Ref | HK23/RR29 | HK25/RR30 | HK26/RR30 | HK30/RR32 |
| OmpR family | |||||||
| PhoR (a) | PhoB | Phosphate | [35] | HK/RR * | HK/RR | HK/RR | HK/RR |
| CusS (a) | CusR | Cu2+ | [36] | np | HK/RR | np | HK/RR |
| KdpD (a) | KdpE | PtsN. K+, ATP, Ionic strength | [36,37] | RR * | np * | HK/RR | HK/RR |
| TorS (b) | TorR | TorT, TMAO | [37] | np * | RR * | HK/RR | HK/RR |
| PhoQ (a) | PhoP | Ni2+, Mg2+, Ca2+, SafA Antimicrobial peptide | [36,37] | HK/RR | HK/RR | HK/RR | HK/RR |
| RstB (a) | RstA | Regulation by PhoQ/PhoP | [38] | HK/RR | HK/RR | HK/RR | HK/RR |
| YedV (a) | YedW | Unknown | HK/RR * | HK/RR | HK/RR | HK/RR | |
| BaeS (a) | BaeR | Myricetin, Na tungstate, Zinc | [39] | HK/RR | HK/RR | HK/RR | HK/RR |
| QseC (a) | QseB | Epinephrine, Norepinephrine, AI-3 | [36] | HK/RR | HK/RR | HK/RR | HK/RR |
| EnvZ (a) | OmpR | Osmolality, pH, CHAPS, MzrA | [37] | HK/RR | HK/RR | HK/RR | HK/RR |
| CpxA (a) | CpxR | CpxP, Misfolded envelope proteins (pH, osmotic stress) | [36] | HK/RR | HK/RR | HK/RR | HK/RR |
| BasS (a) | BasR | Indole, Fe2+/3+ | [36,37] | HK/RR * | HK/RR | HK/RR * | HK/RR |
| CreC (a) | CreB | Glycolytic carbon compounds | [36] | RR * | HK/RR | HK/RR | HK/RR |
| ArcB (b) | ArcA | Redox | [37] | HK/RR | HK/RR | HK/RR | HK/RR |
| NarL family | |||||||
| FimZ | np | HK/RR * | HK/RR | HK/RR | |||
| NarX (a) | NarL | Nitrate iron | [37] | HK/RR | HK/RR | HK/RR | HK/RR |
| BarA (b) | UvrY | Formate, Acetate | [40] | HK/RR | HK/RR | HK/RR | HK/RR |
| NarQ (a) | NarP | Nitrate iron | [37] | HK/RR | RR | HK/RR | HK/RR |
| RcsC /RcsD *** | RcsB | Undecaprenyl-(pyro) phosphate, outer membrane protein RcsF | [36] | HK/RR | HK/RR | HK/RR | HK/RR |
| EvgS (b) | EvgA | Mildly acetic pH, Monovalent cation (Na+, K+), Redox, Indole | [37,38,41] | RR * | HK/RR | HK/RR | HK/RR |
| UhpB (a) | UhpA | UhpC | [36] | RR * | HK/RR | HK/RR | HK/RR |
| NtrC family | |||||||
| AtoS (a) | AtoC | Acetoacetate | [36] | np | np | np | HK/RR |
| GlnL (a) (NtrB) | GlnG (NtrC) | 2-ketoglutarate, Glutamine | [42] | HK/RR | HK/RR | HK/RR | HK/RR |
| ZraS (a) | ZraR | Zn2+, Pb2+ | [36] | HK/RR | HK/RR | HK/RR | HK/RR |
| CitT family | |||||||
| CitA (a) | CitB | Citrate | [36] | np | np | RR | HK/RR |
| DcuS (a) | DcuR | Malate, Oxygen | [37] | HK/RR | HK/RR | HK/RR * | HK/RR |
| LytTR family | |||||||
| BtsS (a) | BtsR | Pyruvate | [43] | HK/RR | HK/RR | HK/RR | HK/RR |
| YpdA (a) | YpdB | Pyruvate | [44] | HK/RR | RR* | HK/RR | HK/RR |
| Others | |||||||
| CheA **** | CheB CheY | Metylation of methyl–accepting chemotaxis proteins | [36] | np | HK/RR | RR * | HK/RR |
| np | HK/RR | HK/RR | HK/RR | ||||
| RssB | Unknown | HK/RR | HK/RR | HK/RR | HK/RR | ||
| GlrK (a) | GlrR | Unknown | HK/RR | HK/RR * | HK/RR | HK/RR | |
| ModD(RR) | ModD | ||||||
| Adf1617_05396(RR) | |||||||
| Asd1617_05387(HK) | |||||||
| Effect on | TCS | Gene Regulated by TCS | Targets | Ref. |
|---|---|---|---|---|
| Invasion of epithelial cells | CpxA/CpxR | virF, virB | T3SS a and its effectors | [49,50,51] |
| EnvZ/OmpR | ompC | OmpC porin | [52,53] | |
| ArcB/ArcA | iuc, sit and feo b operons | Iron transport systems | [54] | |
| Survival within macrophages | EvgS/EvgA | emrKY | EmrKY efflux pump | [55] |
| Inflammatory response and resistance to CAMPs c | PhoQ/PhoP | virK, msbB2, membrane biosynthesis genes | Synthesis and modification of cell envelope | [56,57] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pasqua, M.; Coluccia, M.; Eguchi, Y.; Okajima, T.; Grossi, M.; Prosseda, G.; Utsumi, R.; Colonna, B. Roles of Two-Component Signal Transduction Systems in Shigella Virulence. Biomolecules 2022, 12, 1321. https://doi.org/10.3390/biom12091321
Pasqua M, Coluccia M, Eguchi Y, Okajima T, Grossi M, Prosseda G, Utsumi R, Colonna B. Roles of Two-Component Signal Transduction Systems in Shigella Virulence. Biomolecules. 2022; 12(9):1321. https://doi.org/10.3390/biom12091321
Chicago/Turabian StylePasqua, Martina, Marco Coluccia, Yoko Eguchi, Toshihide Okajima, Milena Grossi, Gianni Prosseda, Ryutaro Utsumi, and Bianca Colonna. 2022. "Roles of Two-Component Signal Transduction Systems in Shigella Virulence" Biomolecules 12, no. 9: 1321. https://doi.org/10.3390/biom12091321
APA StylePasqua, M., Coluccia, M., Eguchi, Y., Okajima, T., Grossi, M., Prosseda, G., Utsumi, R., & Colonna, B. (2022). Roles of Two-Component Signal Transduction Systems in Shigella Virulence. Biomolecules, 12(9), 1321. https://doi.org/10.3390/biom12091321