
The Human Mercaptopyruvate Sulfurtransferase TUM1 Is Involved in Moco Biosynthesis, Cytosolic tRNA Thiolation and Cellular Bioenergetics in Human Embryonic Kidney Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cultivation of Mammalian Cell Lines
2.3. Generating TUM1 Knockout Cells with CRISPR/Cas9
2.4. Immunoblotting
2.5. MTT Assay
2.6. Aconitase Activity Assay
2.7. Sulfite Oxidase Activity Assay
2.8. tRNA Extraction and Analysis
2.9. Quantification of Moco and cPMP in HEK293T Cells
2.10. Measurement of Free H2S via Methylene Blue
2.11. Reactive Oxygen Species Quantification
2.12. Measurement of Cellular Bioenergetics
3. Results
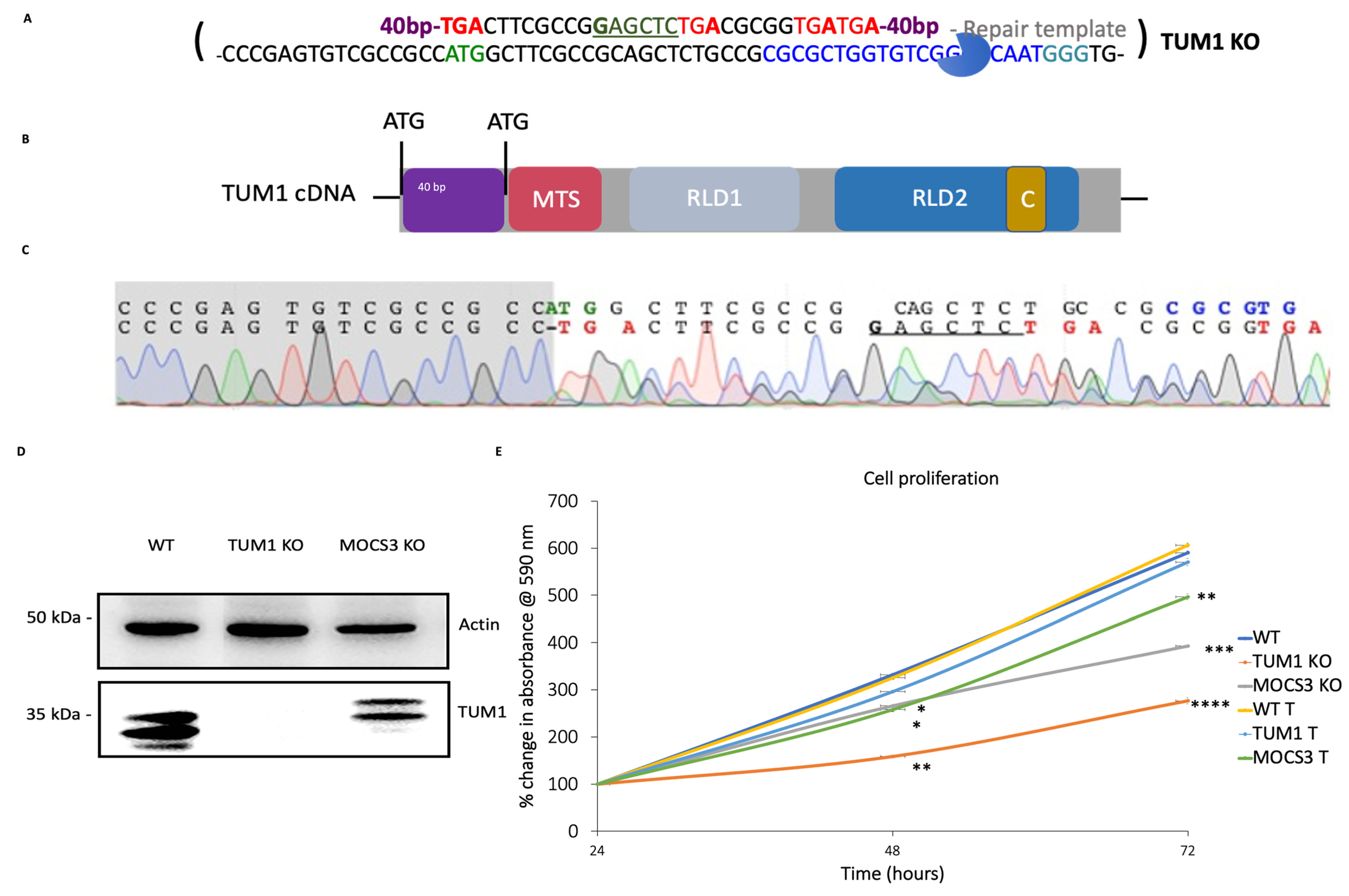
3.1. Generation of HEK293T TUM1 KO Cell Lines with CRISPR/Cas9 System
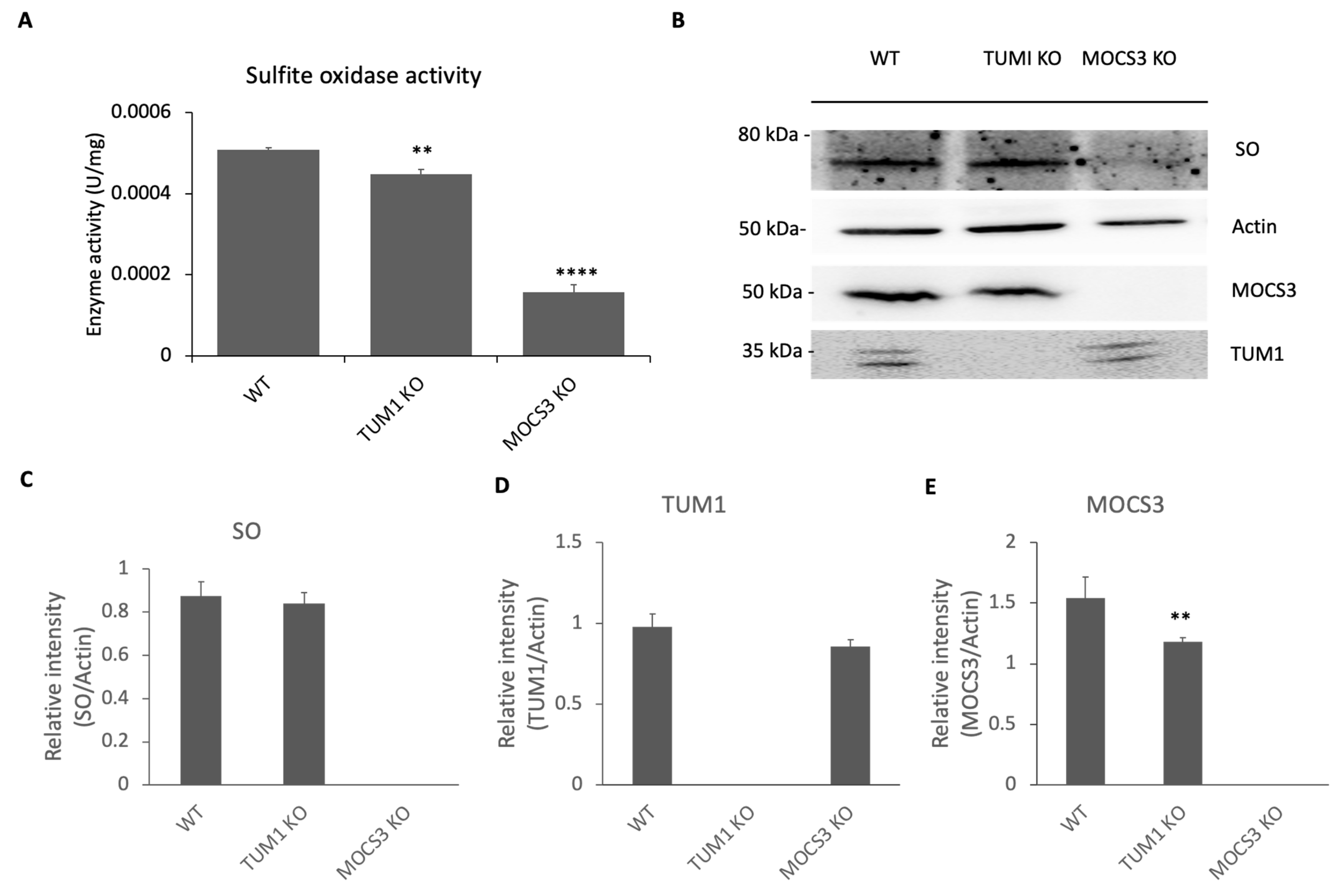
3.2. Effect of TUM1 KO on Sulfite Oxidase Activity and Moco Biosynthesis
3.3. Effect of TUM1 KO on Moco and cPMP Levels
3.4. Repair of Sulfite Oxidase Activity in TUM1 KO Cells with NaHS
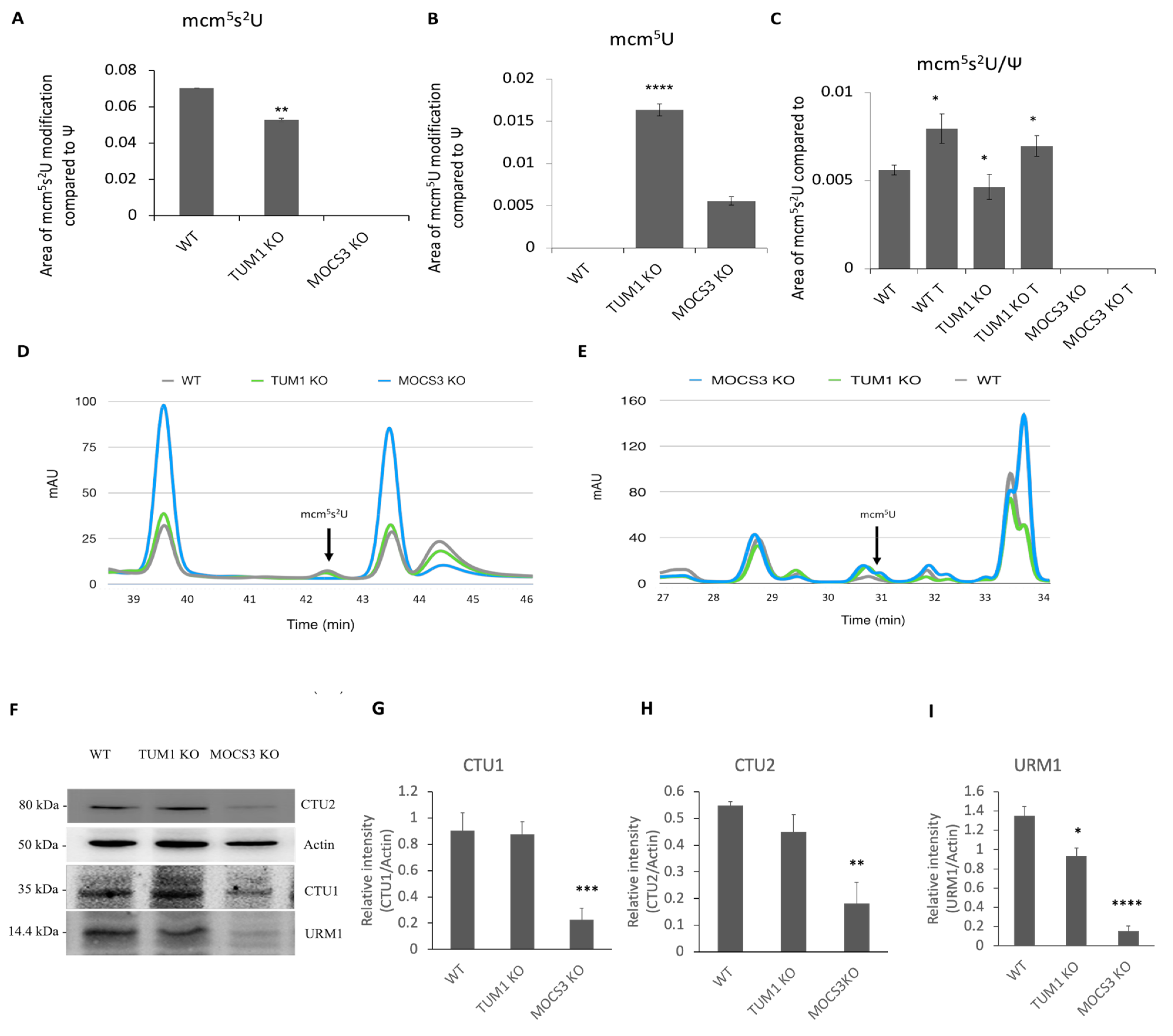
3.5. Effect of TUM1 KO on tRNA Thiolation
3.6. Effect of TUM1 on H2S Biosynthesis
3.7. Effect of TUM1 on Cellular Bioenergetics
3.8. Effect of TUM1 KO on TCA Cycle
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bordo, D.; Bork, P. The rhodanese/Cdc25 phosphatase superfamily. EMBO Rep. 2002, 3, 741–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibuya, N.; Tanaka, M.; Yoshida, M.; Ogasawara, Y.; Togawa, T.; Ishii, K.; Kimura, H. 3-Mercaptopyruvate Sulfurtransferase Produces Hydrogen Sulfide and Bound Sulfane Sulfur in the Brain. Antioxid. Redox Signal. 2009, 11, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.K.; Yamada, K.; Chiku, T.; Koutmos, M.; Banerjee, R. Structure and Kinetic Analysis of H2S Production by Human Mercaptopyruvate Sulfurtransferase. J. Biol. Chem. 2013, 288, 20002–20013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palenchar, P.M.; Buck, C.J.; Cheng, H.; Larson, T.J.; Mueller, E.G. Evidence That ThiI, an Enzyme Shared between Thiamin and 4-Thiouridine Biosynthesis, May Be a Sulfurtransferase That Proceeds through a Persulfide Intermediate. J. Biol. Chem. 2000, 275, 8283–8286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ubuka, T.; Ohta, J.; Yao, W.-B.; Abe, T.; Teraoka, T.; Kurozumi, Y. l-Cysteine metabolism via 3-mercaptopyruvate pathway and sulfate formation in rat liver mitochondria. Amino Acids 1992, 2, 143–155. [Google Scholar] [CrossRef]
- Westley, J.; Adler, H.; Westley, L.; Nishida, C. The sulfurtransferases. Fundam. Appl. Toxicol. 1983, 3, 377–382. [Google Scholar] [CrossRef]
- Fräsdorf, B.; Radon, C.; Leimkühler, S. Characterization and Interaction Studies of Two Isoforms of the Dual Localized 3-Mercaptopyruvate Sulfurtransferase TUM1 from Humans. J. Biol. Chem. 2014, 289, 34543–34556. [Google Scholar] [CrossRef] [Green Version]
- Noma, A.; Sakaguchi, Y.; Suzuki, T. Mechanistic characterization of the sulfur-relay system for eukaryotic 2-thiouridine biogenesis at tRNA wobble positions. Nucleic Acids Res. 2009, 37, 1335–1352. [Google Scholar] [CrossRef] [Green Version]
- Pedre, B.; Dick, T.P. 3-Mercaptopyruvate sulfurtransferase: An enzyme at the crossroads of sulfane sulfur trafficking. Biol. Chem. 2020, 402, 223–237. [Google Scholar] [CrossRef]
- Wróbel, M.; Bronowicka-Adamska, P.; Bentke, A. Hydrogen sulfide generation from L-cysteine in the human glioblastoma-astrocytoma U-87 MG and neuroblastoma SHSY5Y cell lines. Acta Biochim. Pol. 2017, 64, 171–176. [Google Scholar] [CrossRef]
- Gai, J.-W.; Wahafu, W.; Guo, H.; Liu, M.; Wang, X.-C.; Xiao, Y.-X.; Zhang, L.; Xin, Z.-C.; Jin, J. Further evidence of endogenous hydrogen sulphide as a mediator of relaxation in human and rat bladder. Asian J. Androl. 2013, 15, 692–696. [Google Scholar] [CrossRef] [Green Version]
- Ramasamy, S.; Singh, S.; Taniere, P.; Langman, M.J.S.; Eggo, M.C. Sulfide-detoxifying enzymes in the human colon are decreased in cancer and upregulated in differentiation. Am. J. Physiol.-Gastrointest. Liver Physiol. 2006, 291, G288–G296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuhra, K.; Sousa, P.M.F.; Paulini, G.; Lemos, A.R.; Kalme, Z.; Bisenieks, I.; Bisenieks, E.; Vigante, B.; Duburs, G.; Bandeiras, T.M.; et al. Screening Pyridine Derivatives against Human Hydrogen Sulfide-synthesizing Enzymes by Orthogonal Methods. Sci. Rep. 2019, 9, 684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiedler, H.; Wood, J.L. SPECIFICITY STUDIES ON THE β-MERCAPTOPYRUVATE-CYANIDE TRANSSULFURATION SYSTEM. J. Biol. Chem. 1956, 222, 387–397. [Google Scholar] [CrossRef]
- Shen, X.; Pattillo, C.B.; Pardue, S.; Bir, S.C.; Wang, R.; Kevil, C.G. Measurement of plasma hydrogen sulfide in vivo and in vitro. Free. Radic. Biol. Med. 2011, 50, 1021–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vachek, H.; Wood, J.L. Purification and properties of mercaptopyruvate sulfur transferase of Escherichia coli. Biochim. et Biophys. Acta (BBA)—Enzym. 1972, 258, 133–146. [Google Scholar] [CrossRef]
- Taniguchi, T.; Kimura, T. Role of 3-mercaptopyruvate sulfurtransferase in the formation of the iron-sulfur chromophore of adrenal ferredoxin. Biochim. et Biophys. Acta (BBA)—Enzym. 1974, 364, 284–295. [Google Scholar] [CrossRef]
- Crawhall, J.C.; Parker, R.; Sneddon, W.; Young, E.P.; Ampola, M.G.; Efron, M.L.; Bixby, E.M. Beta Mercaptolactate-Cysteine Disulfide: Analog of Cystine in the Urine of a Mentally Retarded Patient. Science 1968, 160, 419–420. [Google Scholar] [CrossRef]
- Hannestad, U.; Mårtensson, J.; Sjödahl, R.; Sörbo, B. 3-Mercaptolactate cysteine disulfiduria: Biochemical studies on affected and unaffected members of a family. Biochem. Med. 1981, 26, 106–114. [Google Scholar] [CrossRef]
- Niederwieser, A.; Giliberti, P.; Baerlocher, K. β-mercaptolactate cysteine disulfiduria in two normal sisters. Isolation and characterization of β-mercaptolactate cysteine disulfide. Clin. Chim. Acta 1973, 43, 405–416. [Google Scholar] [CrossRef]
- Nagahara, N.; Nagano, M.; Ito, T.; Shimamura, K.; Akimoto, T.; Suzuki, H. Antioxidant enzyme, 3-mercaptopyruvate sulfurtransferase-knockout mice exhibit increased anxiety-like behaviors: A model for human mercaptolactate-cysteine disulfiduria. Sci. Rep. 2013, 3, srep01986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, H.; Shibuya, N.; Kimura, Y. Hydrogen Sulfide Is a Signaling Molecule and a Cytoprotectant. Antioxidants Redox Signal. 2012, 17, 45–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendel, R.R.; Leimkühler, S. The biosynthesis of the molybdenum cofactors. JBIC J. Biol. Inorg. Chem. 2014, 20, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Stallmeyer, B.; Drugeon, G.; Reiss, J.; Haenni, A.; Mendel, R. Human Molybdopterin Synthase Gene: Identification of a Bicistronic Transcript with Overlapping Reading Frames. Am. J. Hum. Genet. 1999, 64, 698–705. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, J.; Chowdhury, M.M.; Hänzelmann, P.; Nimtz, M.; Lee, E.-Y.; Schindelin, H.; Leimkühler, S. The Sulfurtransferase Activity of Uba4 Presents a Link between Ubiquitin-like Protein Conjugation and Activation of Sulfur Carrier Proteins. Biochemistry 2008, 47, 6479–6489. [Google Scholar] [CrossRef]
- Hille, R.; Hall, J.; Basu, P. The Mononuclear Molybdenum Enzymes. Chem. Rev. 2014, 114, 3963–4038. [Google Scholar] [CrossRef] [Green Version]
- Cohen, H.J.; Fridovich, I. Hepatic Sulfite Oxidase. Purification and Properties. J. Biol. Chem. 1971, 246, 359–366. [Google Scholar] [CrossRef]
- Woo, W.H.; Yang, H.; Wong, K.P.; Halliwell, B. Sulphite oxidase gene expression in human brain and in other human and rat tissues. Biochem. Biophys. Res. Commun. 2003, 305, 619–623. [Google Scholar] [CrossRef]
- Velayutham, M.; Hemann, C.F.; Cardounel, A.J.; Zweier, J.L. Sulfite oxidase activity of cytochrome c: Role of hydrogen peroxide. Biochem. Biophys. Rep. 2016, 5, 96–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohl, J.B.; Mellis, A.-T.; Schwarz, G. Homeostatic impact of sulfite and hydrogen sulfide on cysteine catabolism. J. Cereb. Blood Flow Metab. 2018, 176, 554–570. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.L.; Duran, M. Molybdenum Cofactor Deficiency and Isolated Sulfite Oxidase Deficiency. In The Online Metabolic and Molecular Bases of Inherited Disease; Valle, D.L., Antonarakis, S., Ballabio, A., Beaudet, A.L., Mitchell, G.A., Eds.; McGraw-Hill Education: New York, NY, USA, 2019. [Google Scholar]
- Mellis, A.-T.; Roeper, J.; Misko, A.L.; Kohl, J.; Schwarz, G. Sulfite Alters the Mitochondrial Network in Molybdenum Cofactor Deficiency. Front. Genet. 2021, 11, 594828. [Google Scholar] [CrossRef] [PubMed]
- Agris, P.F. The importance of being modified: An unrealized code to RNA structure and function. RNA 2015, 21, 552–554. [Google Scholar] [CrossRef]
- Björk, G.R.; Huang, B.; Persson, O.P.; Byström, A.S. A conserved modified wobble nucleoside (mcm5s2U) in lysyl-tRNA is required for viability in yeast. RNA 2007, 13, 1245–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klassen, R.; Grunewald, P.; Thüring, K.L.; Eichler, C.; Helm, M.; Schaffrath, R. Loss of Anticodon Wobble Uridine Modifications Affects tRNALys Function and Protein Levels in Saccharomyces cerevisiae. PLoS ONE 2015, 10, e0119261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leidel, S.; Pedrioli, P.G.A.; Bucher, T.; Brost, R.; Costanzo, M.; Schmidt, A.; Aebersold, R.; Boone, C.; Hofmann, K.; Peter, M. Ubiquitin-related modifier Urm1 acts as a sulphur carrier in thiolation of eukaryotic transfer RNA. Nature 2009, 458, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Nedialkova, D.D.; Leidel, S.A. Optimization of Codon Translation Rates via tRNA Modifications Maintains Proteome Integrity. Cell 2015, 161, 1606–1618. [Google Scholar] [CrossRef] [Green Version]
- Yarian, C.; Townsend, H.; Czestkowski, W.; Sochacka, E.; Malkiewicz, A.J.; Guenther, R.; Miskiewicz, A.; Agris, P.F. Accurate Translation of the Genetic Code Depends on tRNA Modified Nucleosides. J. Biol. Chem. 2002, 277, 16391–16395. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, M.M.; Dosche, C.; Löhmannsröben, H.-G.; Leimkühler, S. Dual Role of the Molybdenum Cofactor Biosynthesis Protein MOCS3 in tRNA Thiolation and Molybdenum Cofactor Biosynthesis in Humans. J. Biol. Chem. 2012, 287, 17297–17307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leimkühler, S.; Bühning, M.; Beilschmidt, L. Shared Sulfur Mobilization Routes for tRNA Thiolation and Molybdenum Cofactor Biosynthesis in Prokaryotes and Eukaryotes. Biomolecules 2017, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Van der Veen, A.G.; Schorpp, K.; Schlieker, C.; Buti, L.; Damon, J.R.; Spooner, E.; Ploegh, H.L.; Jentsch, S. Role of the ubiquitin-like protein Urm1 as a noncanonical lysine-directed protein modifier. Proc. Natl. Acad. Sci. USA 2011, 108, 1763–1770. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Huang, B.; Eliasson, M.; Rydén, P.; Byström, A.S. Elongator Complex Influences Telomeric Gene Silencing and DNA Damage Response by Its Role in Wobble Uridine tRNA Modification. PLOS Genet. 2011, 7, e1002258. [Google Scholar] [CrossRef] [Green Version]
- Pedrioli, P.G.A.; Leidel, S.; Hofmann, K. Urm1 at the crossroad of modifications. EMBO Rep. 2008, 9, 1196–1202. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, J.; Wang, L.; Zhou, J.; Huang, H.; Wu, J.; Zhong, Y.; Shi, Y. Solution structure of Urm1 and its implications for the origin of protein modifiers. Proc. Natl. Acad. Sci. USA 2006, 103, 11625–11630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Wada, T.; Saigo, K.; Watanabe, K. Taurine as a constituent of mitochondrial tRNAs: New insights into the functions of taurine and human mitochondrial diseases. EMBO J. 2002, 21, 6581–6589. [Google Scholar] [CrossRef] [Green Version]
- Kirino, Y.; Suzuki, T. Human Mitochondrial Diseases Associated with tRNA Wobble Modification Deficiency. RNA Biol. 2005, 2, 41–44. [Google Scholar] [CrossRef] [Green Version]
- Umeda, N.; Suzuki, T.; Yukawa, M.; Ohya, Y.; Shindo, H.; Watanabe, K.; Suzuki, T. Mitochondria-specific RNA-modifying Enzymes Responsible for the Biosynthesis of the Wobble Base in Mitochondrial tRNAs. J. Biol. Chem. 2005, 280, 1613–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T. Biosynthesis and function of tRNA wobble modifications. In Fine-Tuning of RNA Functions by Modification and Editing; Springer: Berlin/Heidelberg, Germany, 2005; pp. 23–69. [Google Scholar] [CrossRef]
- Abe, K.; Kimura, H. The possible role of hydrogen sulfide as an endogenous neuromodulator. J. Neurosci. 1996, 16, 1066–1071. [Google Scholar] [CrossRef] [Green Version]
- Hosoki, R.; Matsuki, N.; Kimura, H. The Possible Role of Hydrogen Sulfide as an Endogenous Smooth Muscle Relaxant in Synergy with Nitric Oxide. Biochem. Biophys. Res. Commun. 1997, 237, 527–531. [Google Scholar] [CrossRef]
- Nakai, Y.; Nakai, M.; Hayashi, H. Thio-modification of Yeast Cytosolic tRNA Requires a Ubiquitin-related System That Resembles Bacterial Sulfur Transfer Systems. J. Biol. Chem. 2008, 283, 27469–27476. [Google Scholar] [CrossRef]
- Land, T.; Rouault, T.A. Targeting of a Human Iron–Sulfur Cluster Assembly Enzyme, nifs, to Different Subcellular Compartments Is Regulated through Alternative AUG Utilization. Mol. Cell 1998, 2, 807–815. [Google Scholar] [CrossRef]
- Marelja, Z.; Chowdhury, M.M.; Dosche, C.; Hille, C.; Baumann, O.; Löhmannsröben, H.-G.; Leimkühler, S. The L-Cysteine Desulfurase NFS1 Is Localized in the Cytosol where it Provides the Sulfur for Molybdenum Cofactor Biosynthesis in Humans. PLoS ONE 2013, 8, e60869. [Google Scholar] [CrossRef] [Green Version]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guide design resources—Zhang Lab. Available online: https://zlab.bio/guide-design-resources (accessed on 16 August 2022).
- Addgene: CRISPR Plasmids and Resources. Available online: https://www.addgene.org/crispr/ (accessed on 1 September 2017).
- Freimoser, F.M.; Jakob, C.A.; Aebi, M.; Tuor, U. The MTT [3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide] Assay Is a Fast and Reliable Method for Colorimetric Determination of Fungal Cell Densities. Appl. Environ. Microbiol. 1999, 65, 3727–3729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beinert, H.; Kennedy, M.C. Aconitase, a two-faced protein: Enzyme and iron regulatory factor 1 2. FASEB J. 1993, 7, 1442–1449. [Google Scholar] [CrossRef] [PubMed]
- Uhrigshardt, H.; Singh, A.; Kovtunovych, G.; Ghosh, M.; Rouault, T.A. Characterization of the human HSC20, an unusual DnaJ type III protein, involved in iron–sulfur cluster biogenesis. Hum. Mol. Genet. 2010, 19, 3816–3834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.L.; Rajagopalan, K.V.; Lanman, J.T.; Schutgens, R.B.H.; van Gennip, A.H.; Sorensen, P.; Applegarth, D.A. Prenatal diagnosis of molybdenum cofactor deficiency by assay of sulphite oxidase activity in chorionic villus samples. J. Inherit. Metab. Dis. 1991, 14, 932–937. [Google Scholar] [CrossRef]
- Gehrke, C.W.; Kuo, K.C. Ribonucleoside analysis by reversed-phase high-performance liquid chromatography. J. Chromatogr. A 1989, 471, 3–36. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.L.; Hainline, B.E.; Rajagopalan, K.V.; Arison, B.H. The pterin component of the molybdenum cofactor. Structural characterization of two fluorescent derivatives. J. Biol. Chem. 1984, 259, 5414–5422. [Google Scholar] [CrossRef]
- Fogo, J.K.; Popowsky, M. Spectrophotometric Determination of Hydrogen Sulfide. Anal. Chem. 1949, 21, 732–734. [Google Scholar] [CrossRef]
- Wu, D.; Yotnda, P. Production and Detection of Reactive Oxygen Species (ROS) in Cancers. J. Vis. Exp. 2011, 57, e3357. [Google Scholar] [CrossRef] [PubMed]
- van der Windt, G.J.; Chang, C.; Pearce, E.L. Measuring Bioenergetics in T Cells Using a Seahorse Extracellular Flux Analyzer. Curr. Protoc. Immunol. 2016, 113, 3.16B.1–3.16B.14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neukranz, Y.; Kotter, A.; Beilschmidt, L.; Marelja, Z.; Helm, M.; Gräf, R.; Leimkühler, S. Analysis of the Cellular Roles of MOCS3 Identifies a MOCS3-Independent Localization of NFS1 at the Tips of the Centrosome. Biochemistry 2019, 58, 1786–1798. [Google Scholar] [CrossRef] [PubMed]
- Greiner, R.; Pálinkás, Z.; Bäsell, K.; Becher, D.; Antelmann, H.; Nagy, P.; Dick, T.P. Polysulfides Link H2S to Protein Thiol Oxidation. Antioxidants Redox Signal. 2013, 19, 1749–1765. [Google Scholar] [CrossRef] [Green Version]
- Szabo, C.; Ransy, C.; Módis, K.; Andriamihaja, M.; Murghes, B.; Coletta, C.; Olah, G.; Yanagi, K.; Bouillaud, F. Regulation of mitochondrial bioenergetic function by hydrogen sulfide. Part I. Biochemical and physiological mechanisms. Br. J. Pharmacol. 2014, 171, 2099–2122. [Google Scholar] [CrossRef] [Green Version]
- Módis, K.; Coletta, C.; Erdélyi, K.; Papapetropoulos, A.; Szabo, C. Intramitochondrial hydrogen sulfide production by 3-mercaptopyruvate sulfurtransferase maintains mitochondrial electron flow and supports cellular bioenergetics. FASEB J. 2012, 27, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.M.; Schwarz, G. Cofactor-dependent maturation of mammalian sulfite oxidase links two mitochondrial import pathways. J. Cell Sci. 2012, 125, 4876–4885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warnhoff, K.; Ruvkun, G. Molybdenum cofactor transfer from bacteria to nematode mediates sulfite detoxification. Nat. Chem. Biol. 2019, 15, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Jüdes, A.; Ebert, F.; Bär, C.; Thüring, K.L.; Harrer, A.; Klassen, R.; Helm, M.; Stark, M.J.; Schaffrath, R. Urmylation and tRNA thiolation functions of ubiquitin-like Uba4·Urm1 systems are conserved from yeast to man. FEBS Lett. 2015, 589, 904–909. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Ding, L.; Xie, Z.Z.; Yang, Y.; Whiteman, M.; Moore, P.K.; Bian, J.S. A Review of Hydrogen Sulfide Synthesis, Metabolism, and Measurement: Is Modulation of Hydrogen Sulfide a Novel Therapeutic for Cancer? Antioxidants Redox Signal. 2019, 31, 1–38. [Google Scholar] [CrossRef]
- Módis, K.; Ju, Y.; Ahmad, A.; Untereiner, A.A.; Altaany, Z.; Wu, L.; Szabo, C.; Wang, R. S- Sulfhydration of ATP synthase by hydrogen sulfide stimulates mitochondrial bioenergetics. Pharmacol. Res. 2016, 113, 116–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borisov, V.B.; Forte, E. Impact of Hydrogen Sulfide on Mitochondrial and Bacterial Bioenergetics. Int. J. Mol. Sci. 2021, 22, 12688. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, O.V.; Piroddi, M.; Galli, F.; Lushchak, V.I. Aconitase post-translational modification as a key in linkage between Krebs cycle, iron homeostasis, redox signaling, and metabolism of reactive oxygen species. Redox Rep. 2013, 19, 8–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhami, N.; Trivedi, D.K.; Goodacre, R.; Mainwaring, D.; Humphreys, D.P. Mitochondrial aconitase is a key regulator of energy production for growth and protein expression in Chinese hamster ovary cells. Metabolomics 2018, 14, 136. [Google Scholar] [CrossRef] [PubMed]
- Ray, W.K.; Zeng, G.; Potters, M.B.; Mansuri, A.M.; Larson, T.J. Characterization of a 12-Kilodalton Rhodanese Encoded by glpE of Escherichia coli and Its Interaction with Thioredoxin. J. Bacteriol. 2000, 182, 2277–2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spallarossa, A.; Donahue, J.L.; Larson, T.J.; Bolognesi, M.; Bordo, D. Escherichia coli GlpE Is a Prototype Sulfurtransferase for the Single-Domain Rhodanese Homology Superfamily. Structure 2001, 9, 1117–1125. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Gladyshev, V.N. Molybdoproteomes and Evolution of Molybdenum Utilization. J. Mol. Biol. 2008, 379, 881–899. [Google Scholar] [CrossRef] [Green Version]
- Kessler, D. Enzymatic activation of sulfur for incorporation into biomolecules in prokaryotes. FEMS Microbiol. Rev. 2006, 30, 825–840. [Google Scholar] [CrossRef] [Green Version]
- Hidese, R.; Mihara, H.; Esaki, N. Bacterial cysteine desulfurases: Versatile key players in biosynthetic pathways of sulfur-containing biofactors. Appl. Microbiol. Biotechnol. 2011, 91, 47–61. [Google Scholar] [CrossRef]
- Rydz, L.; Wróbel, M.; Jurkowska, H. Sulfur Administration in Fe–S Cluster Homeostasis. Antioxidants 2021, 10, 1738. [Google Scholar] [CrossRef]
- Billaut-Laden, I.; Rat, E.; Allorge, D.; Crunelle-Thibaut, A.; Cauffiez, C.; Chevalier, D.; Lo-Guidice, J.-M.; Broly, F. Evidence for a functional genetic polymorphism of the human mercaptopyruvate sulfurtransferase (MPST), a cyanide detoxification enzyme. Toxicol. Lett. 2006, 165, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Duran, M.; Beemer, F.A.; Heiden, C.V.D.; Korteland, J.; de Bree, P.K.; Brink, M.; Wadman, S.K.; Lombeck, I. Combined deficiency of xanthine oxidase and sulphite oxidase: A defect of molybdenum metabolism or transport? J. Inherit. Metab. Dis. 1978, 1, 175–178. [Google Scholar] [CrossRef]
- Benchoam, D.; Cuevasanta, E.; Möller, M.N.; Alvarez, B. Hydrogen Sulfide and Persulfides Oxidation by Biologically Relevant Oxidizing Species. Antioxidants 2019, 8, 48. [Google Scholar] [CrossRef] [Green Version]
- Łowicka, E.; Bełtowski, J. Hydrogen sulfide (H2S)—The third gas of interest for pharmacologists. Pharmacol Rep. 2007, 59, 4–24. [Google Scholar] [PubMed]
- Paul, B.D.; Snyder, S.H. H2S signalling through protein sulfhydration and beyond. Nat. Rev. Mol. Cell Biol. 2012, 13, 499–507. [Google Scholar] [CrossRef]
- Koike, S.; Nishimoto, S.; Ogasawara, Y. Cysteine persulfides and polysulfides produced by exchange reactions with H2S protect SH-SY5Y cells from methylglyoxal-induced toxicity through Nrf2 activation. Redox Biol. 2017, 12, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Hydrogen Sulfide and Polysulfide Signaling. Antioxidants Redox Signal. 2017, 27, 619–621. [Google Scholar] [CrossRef] [PubMed]
- Chiku, T.; Padovani, D.; Zhu, W.; Singh, S.; Vitvitsky, V.; Banerjee, R. H2S Biogenesis by Human Cystathionine γ-Lyase Leads to the Novel Sulfur Metabolites Lanthionine and Homolanthionine and Is Responsive to the Grade of Hyperhomocysteinemia. J. Biol. Chem. 2009, 284, 11601–11612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, B.D.; Snyder, S.H.; Kashfi, K. Effects of hydrogen sulfide on mitochondrial function and cellular bioenergetics. Redox Biol. 2020, 38, 101772. [Google Scholar] [CrossRef]
- Damon, J.R.; Pincus, D.; Ploegh, H.L. tRNA thiolation links translation to stress responses in Saccharomyces cerevisiae. Mol. Biol. Cell 2015, 26, 270–282. [Google Scholar] [CrossRef]
- Martínez-Reyes, I.; Diebold, L.P.; Kong, H.; Schieber, M.; Huang, H.; Hensley, C.T.; Mehta, M.M.; Wang, T.; Santos, J.H.; Woychik, R.; et al. TCA Cycle and Mitochondrial Membrane Potential Are Necessary for Diverse Biological Functions. Mol. Cell 2015, 61, 199–209. [Google Scholar] [CrossRef] [Green Version]
- Finkelstein, J.D. Methionine metabolism in mammals. J. Nutr. Biochem. 1990, 1, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Módis, K.; Asimakopoulou, A.; Coletta, C.; Papapetropoulos, A.; Szabo, C. Oxidative stress suppresses the cellular bioenergetic effect of the 3-mercaptopyruvate sulfurtransferase/hydrogen sulfide pathway. Biochem. Biophys. Res. Commun. 2013, 433, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, T.M.; Grieshaber, M.K. Three enzymatic activities catalyze the oxidation of sulfide to thiosulfate in mammalian and invertebrate mitochondria. FEBS J. 2008, 275, 3352–3361. [Google Scholar] [CrossRef]
- Fu, M.; Zhang, W.; Wu, L.; Yang, G.; Li, H.; Wang, R. Hydrogen sulfide (H2S) metabolism in mitochondria and its regulatory role in energy production. Proc. Natl. Acad. Sci. USA 2012, 109, 2943–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trautwein, B.; Merz, T.; Denoix, N.; Szabo, C.; Calzia, E.; Radermacher, P.; McCook, O. ΔMST and the Regulation of Cardiac CSE and OTR Expression in Trauma and Hemorrhage. Antioxidants 2021, 10, 233. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.; Bhattacharya, R.; Mukherjee, P. Hydrogen sulfide signaling in mitochondria and disease. FASEB J. 2019, 33, 13098–13125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Módis, K.; Ramanujam, V.-M.S.; Govar, A.A.; Lopez, E.; Anderson, K.E.; Wang, R.; Szabo, C. Cystathionine-γ-lyase (CSE) deficiency increases erythropoiesis and promotes mitochondrial electron transport via the upregulation of coproporphyrinogen III oxidase and consequent stimulation of heme biosynthesis. Biochem. Pharmacol. 2019, 169, 113604. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Vincent, A.S.; Halliwell, B.; Wong, K.P. A Mechanism of Sulfite Neurotoxicity. J. Biol. Chem. 2004, 279, 43035–43045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herst, P.M.; Tan, A.S.; Scarlett, D.-J.G.; Berridge, M.V. Cell surface oxygen consumption by mitochondrial gene knockout cells. Biochim. et Biophys. Acta 2004, 1656, 79–87. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogunkola, M.O.; Guiraudie-Capraz, G.; Feron, F.; Leimkühler, S. The Human Mercaptopyruvate Sulfurtransferase TUM1 Is Involved in Moco Biosynthesis, Cytosolic tRNA Thiolation and Cellular Bioenergetics in Human Embryonic Kidney Cells. Biomolecules 2023, 13, 144. https://doi.org/10.3390/biom13010144
Ogunkola MO, Guiraudie-Capraz G, Feron F, Leimkühler S. The Human Mercaptopyruvate Sulfurtransferase TUM1 Is Involved in Moco Biosynthesis, Cytosolic tRNA Thiolation and Cellular Bioenergetics in Human Embryonic Kidney Cells. Biomolecules. 2023; 13(1):144. https://doi.org/10.3390/biom13010144
Chicago/Turabian StyleOgunkola, Moses Olalekan, Gaelle Guiraudie-Capraz, Francois Feron, and Silke Leimkühler. 2023. "The Human Mercaptopyruvate Sulfurtransferase TUM1 Is Involved in Moco Biosynthesis, Cytosolic tRNA Thiolation and Cellular Bioenergetics in Human Embryonic Kidney Cells" Biomolecules 13, no. 1: 144. https://doi.org/10.3390/biom13010144
APA StyleOgunkola, M. O., Guiraudie-Capraz, G., Feron, F., & Leimkühler, S. (2023). The Human Mercaptopyruvate Sulfurtransferase TUM1 Is Involved in Moco Biosynthesis, Cytosolic tRNA Thiolation and Cellular Bioenergetics in Human Embryonic Kidney Cells. Biomolecules, 13(1), 144. https://doi.org/10.3390/biom13010144