
Development of an Anti-HER2 Single-Chain Variable Antibody Fragment Construct for High-Yield Soluble Expression in Escherichia coli and One-Step Chromatographic Purification


 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. DNA Recombination
2.3. Protein Sample Preparation
2.4. Enzyme-Linked Immunosorbent Assay
2.5. Flow Cytometry
2.6. Statistics
3. Results
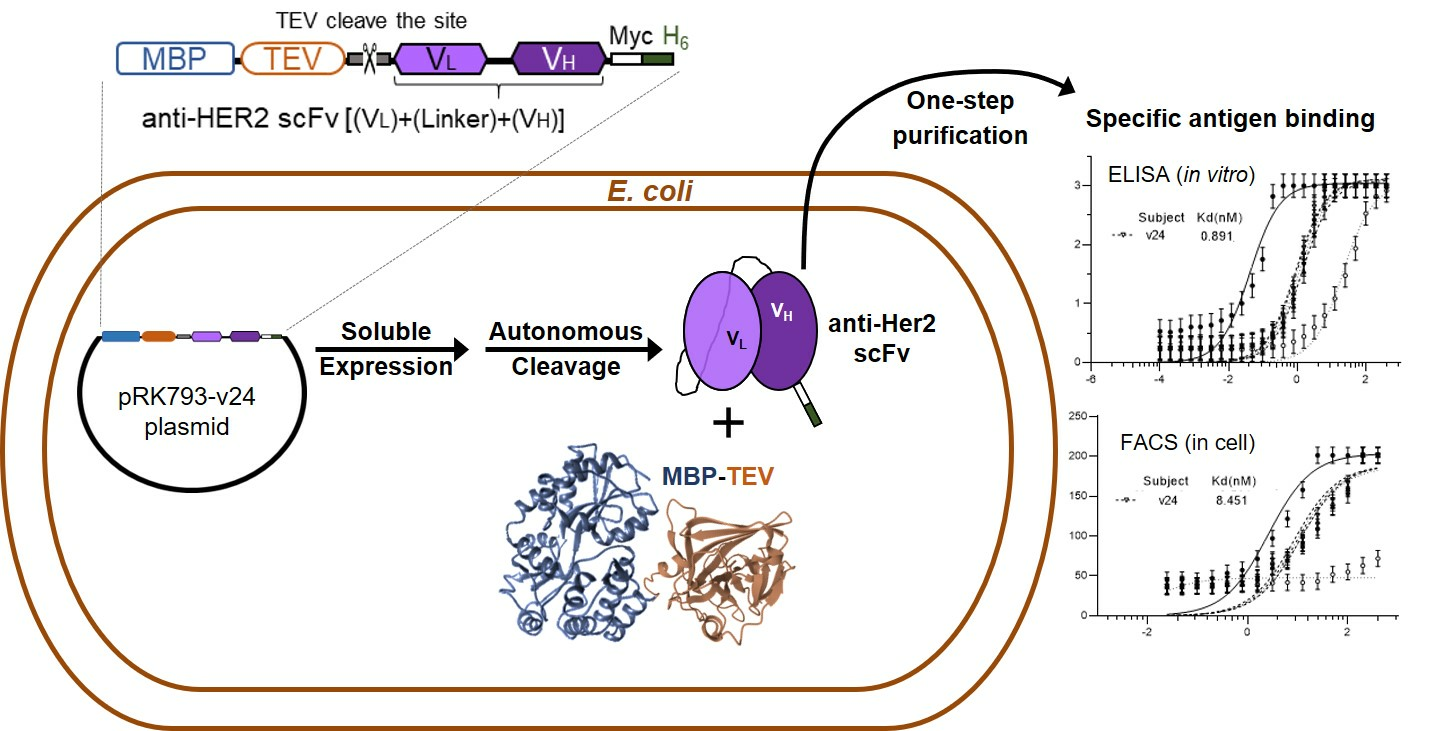
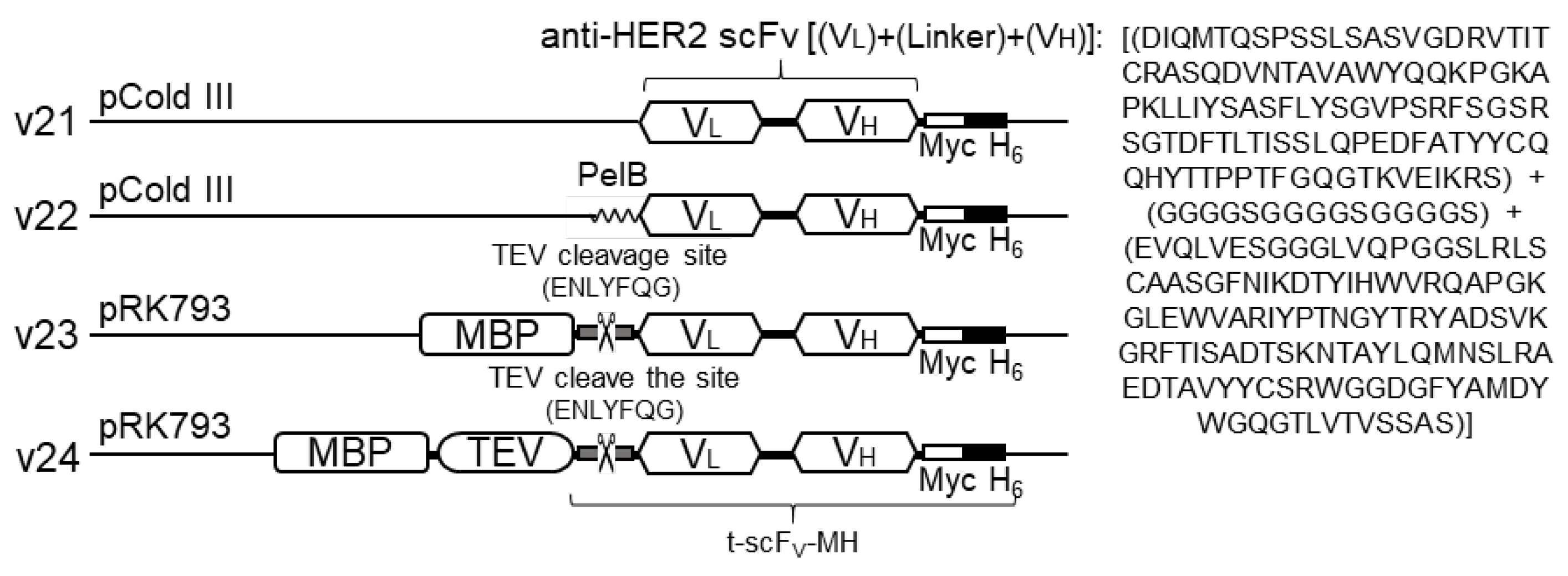
3.1. Design of Anti-HER2 scFv-Expressing Construts
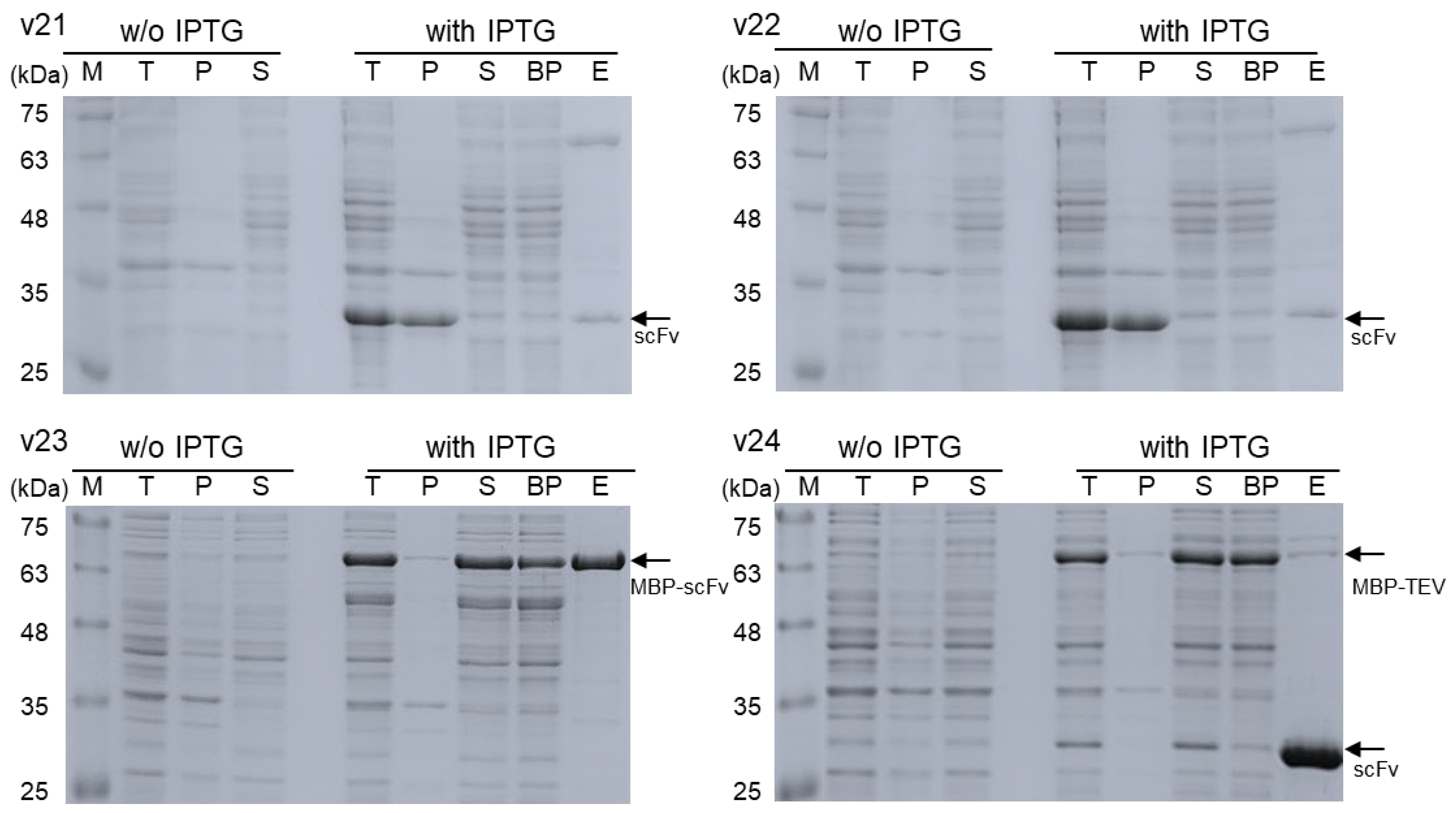
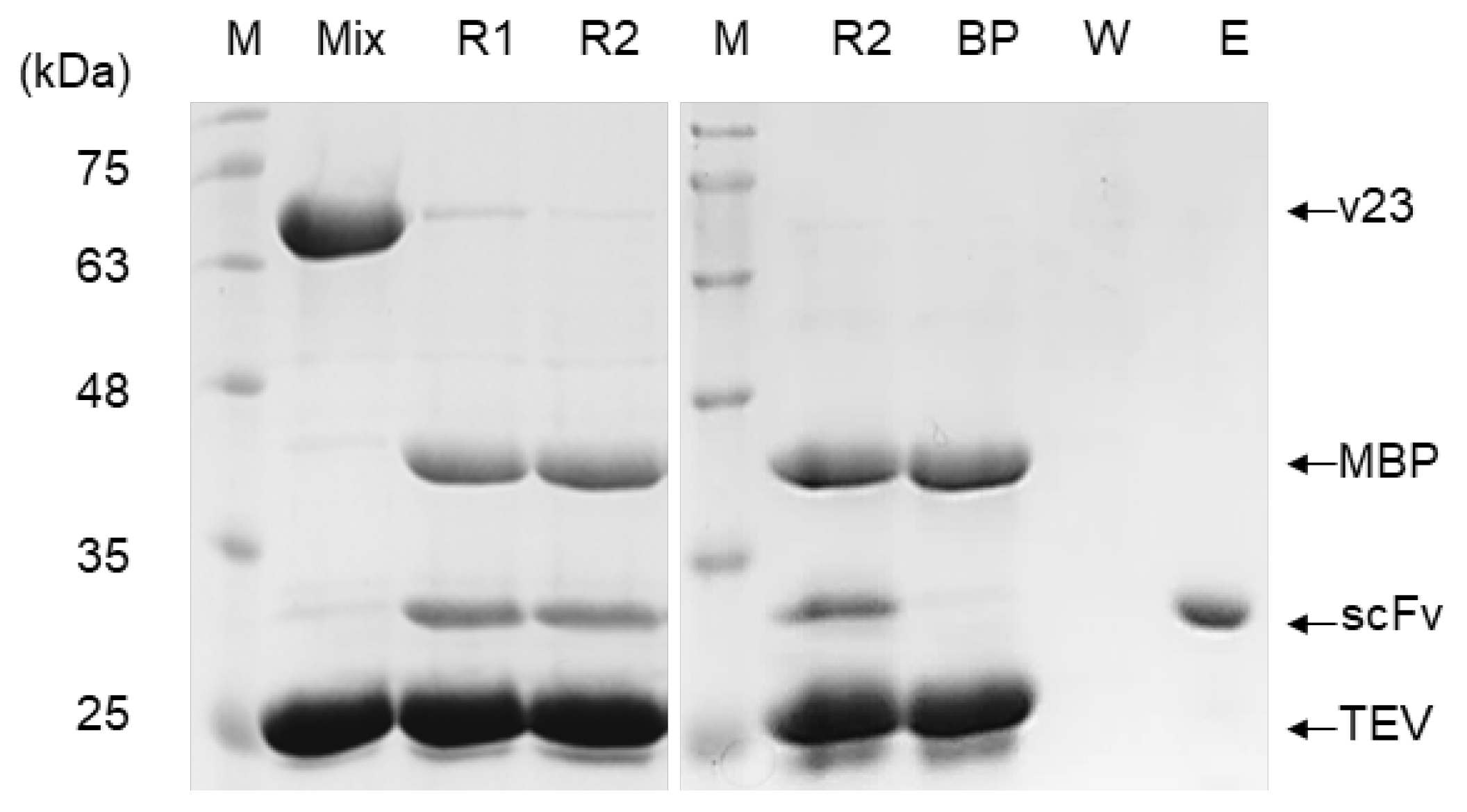
3.2. Comparison in Soluble Expression and Purification
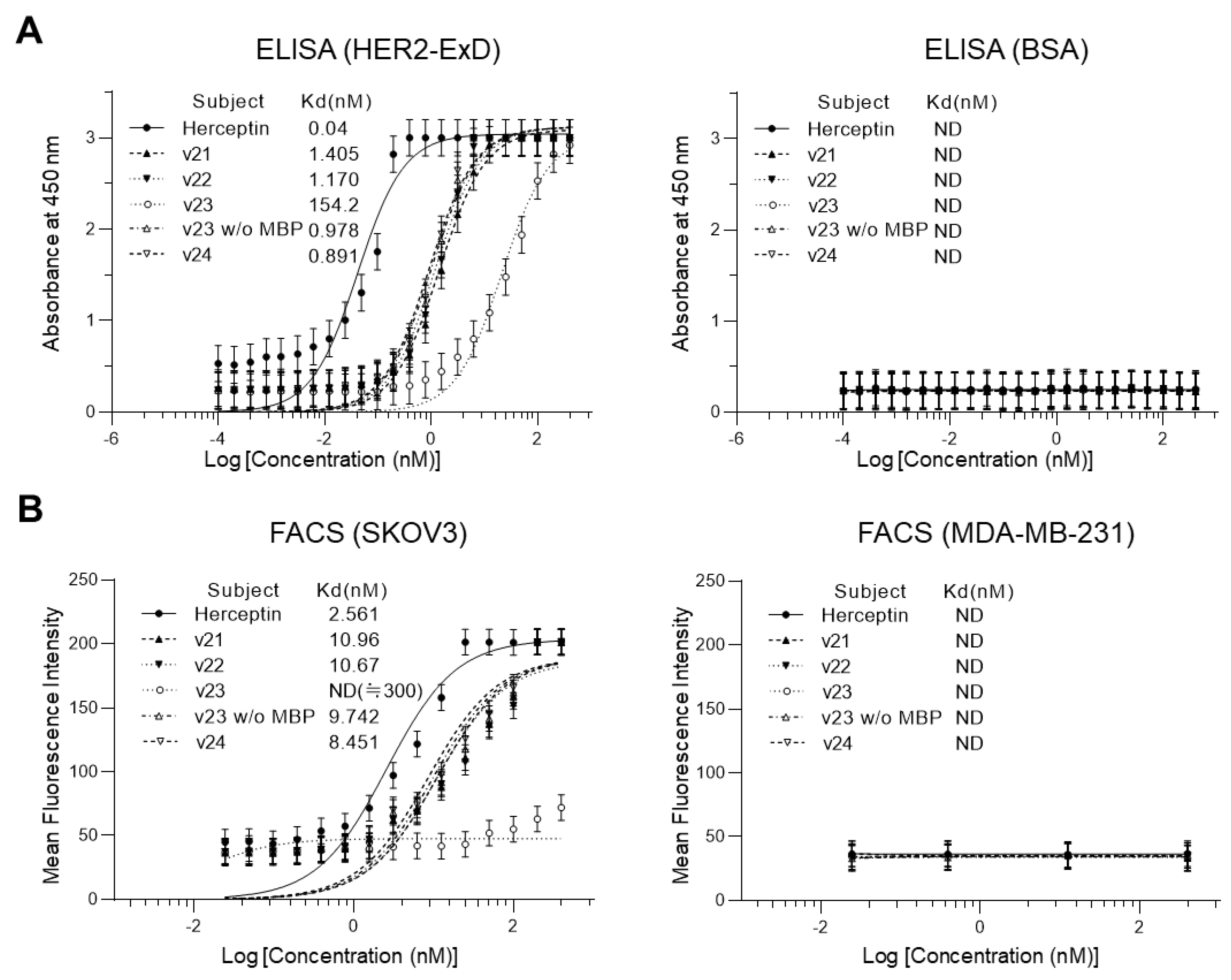
3.3. Comparison in Antigen-Binding Activity
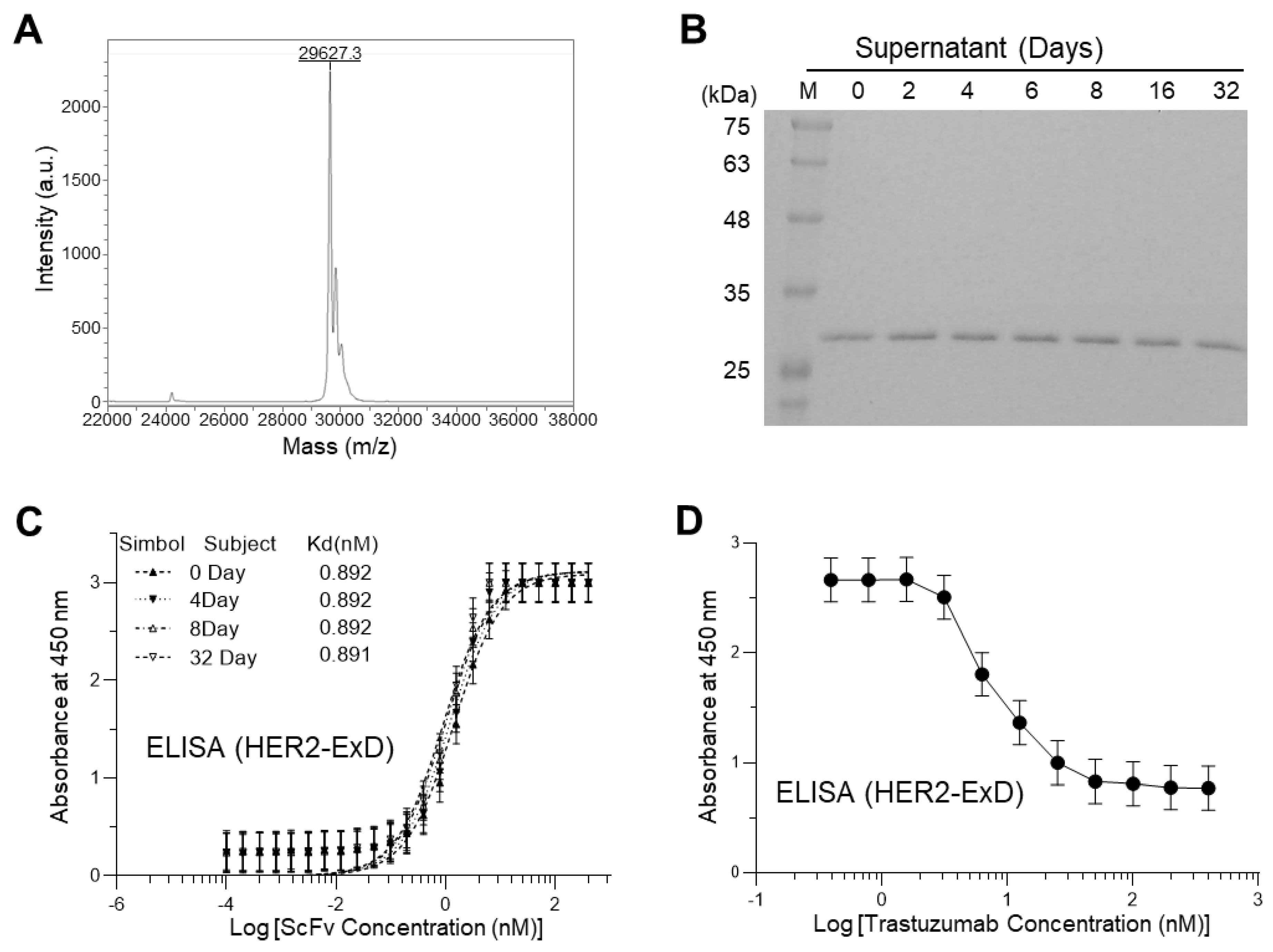
3.4. Identification and Validation of Functionality and Stability
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aguiar, S.; Dias, J.; Manuel, A.M.; Russo, R.; Gois, P.M.P.; da Silva, F.A.; Goncalves, J. Chimeric Small Antibody Fragments as Strategy to Deliver Therapeutic Payloads. Adv. Protein Chem. Struct. Biol. 2018, 112, 143–182. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.; Huang, M.; Lum, L.G. Bispecific antibody based therapeutics: Strengths and challenges. Blood Rev. 2018, 32, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Bates, A.; Power, C.A. David vs. Goliath: The Structure, Function, and Clinical Prospects of Antibody Fragments. Antibodies 2019, 8, 28. [Google Scholar] [CrossRef]
- Sim, D.; Song, J.; Kim, J.; Lee, J.; Chung, D.; Jo, K.; Kim, C.; Seo, M.; Kang, H.C.; Paeng, J.C. Enhancing bacterial production of a recombinant cetuximab-Fab by partial humanization and its utility for drug conjugation. Process Biochem. 2018, 69, 123–130. [Google Scholar] [CrossRef]
- Valldorf, B.; Hinz, S.C.; Russo, G.; Pekar, L.; Mohr, L.; Klemm, J.; Doerner, A.; Krah, S.; Hust, M.; Zielonka, S. Antibody display technologies: Selecting the cream of the crop. Biol. Chem. 2022, 403, 455–477. [Google Scholar] [CrossRef]
- Sergeeva, A.; Kolonin, M.G.; Molldrem, J.J.; Pasqualini, R.; Arap, W. Display technologies: Application for the discovery of drug and gene delivery agents. Adv. Drug Deliv. Rev. 2006, 58, 1622–1654. [Google Scholar] [CrossRef] [PubMed]
- Ellerman, D. Bispecific T-cell engagers: Towards understanding variables influencing the in vitro potency and tumor selectivity and their modulation to enhance their efficacy and safety. Methods 2019, 154, 102–117. [Google Scholar] [CrossRef]
- Yu, S.; Yi, M.; Qin, S.; Wu, K. Next generation chimeric antigen receptor T cells: Safety strategies to overcome toxicity. Mol. Cancer 2019, 18, 125. [Google Scholar] [CrossRef]
- Graff, S.L.; Yan, F.; Abdou, Y. Newly Approved and Emerging Agents in HER2-Positive Metastatic Breast Cancer. Clin. Breast Cancer 2023, 23, e380–e393. [Google Scholar] [CrossRef]
- Akbari, V.; Chou, C.P.; Abedi, D. New insights into affinity proteins for HER2-targeted therapy: Beyond trastuzumab. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188448. [Google Scholar] [CrossRef]
- Iqbal, N.; Iqbal, N. Human Epidermal Growth Factor Receptor 2 (HER2) in Cancers: Overexpression and Therapeutic Implications. Mol. Biol. Int. 2014, 2014, 852748. [Google Scholar] [CrossRef] [PubMed]
- Ueda, M.; Hisada, H.; Temma, T.; Shimizu, Y.; Kimura, H.; Ono, M.; Nakamoto, Y.; Togashi, K.; Saji, H. Gallium-68-labeled anti-HER2 single-chain Fv fragment: Development and in vivo monitoring of HER2 expression. Mol. Imaging Biol. 2015, 17, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.; Boleti, E.; George, A.J.T. Antibody engineering: Comparison of bacterial, yeast, insect and mammalian expression systems. J. Immunol. Methods 1998, 216, 165. [Google Scholar] [CrossRef]
- Kim, J.; Sim, D.; Park, D.; Jung, T.; Lee, S.; Oh, T.; Ha, J.; Seok, S.; Seo, M.; Kang, H.C.; et al. Bacterial production and structure-functional validation of a recombinant antigen-binding fragment (Fab) of an anti-cancer therapeutic antibody targeting epidermal growth factor receptor. Appl. Microbiol. Biotechnol. 2016, 100, 10521–10529. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.; Lee, K.; Kim, J.; Sim, D.; Byun, K.; Jung, S.; Paeng, J.C.; Kang, T.; Park, J.; Kim, C. Extracellular production of an anti-HER2 single-chain variable antibody fragment in Escherichia coli. Process Biochem. 2021, 111, 87–94. [Google Scholar] [CrossRef]
- Esposito, D.; Chatterjee, D.K. Enhancement of soluble protein expression through the use of fusion tags. Curr. Opin. Biotechnol. 2006, 17, 353–358. [Google Scholar] [CrossRef]
- Sonoda, H.; Kumada, Y.; Katsuda, T.; Yamaji, H. Effects of cytoplasmic and periplasmic chaperones on secretory production of single-chain Fv antibody in Escherichia coli. J. Biosci. Bioeng. 2011, 111, 465–470. [Google Scholar]
- De Marco, A.; De Marco, V. Bacteria co-transformed with recombinant proteins and chaperones cloned in independent plasmids are suitable for expression tuning. J. Biotechnol. 2004, 109, 45–52. [Google Scholar] [CrossRef]
- Fu, X. Extracellular accumulation of recombinant protein by Escherichia coli in a defined medium. Appl. Microbiol. Biotechnol. 2010, 88, 75–86. [Google Scholar] [CrossRef]
- Sadraeian, M.; Ghoshoon, M.B.; Mohkam, M.; Karimi, Z.; Rasoul-Amini, S.; Ghasemi, Y. Modification in media composition to obtain secretory production of STxB-based vaccines using Escherichia coli. Virol. Sin. 2013, 28, 43–48. [Google Scholar] [CrossRef]
- Kudou, M.; Ejima, D.; Sato, H.; Yumioka, R.; Arakawa, T.; Tsumoto, K. Refolding single-chain antibody (scFv) using lauroyl-L-glutamate as a solubilization detergent and arginine as a refolding additive. Protein Expr. Purif. 2011, 77, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Goulding, C.W.; Perry, L.J. Protein production in Escherichia coli for structural studies by X-ray crystallography. J. Struct. Biol. 2003, 142, 133–143. [Google Scholar] [CrossRef]
- Tropea, J.E.; Cherry, S.; Waugh, D.S. Expression and purification of soluble His 6-tagged TEV protease. In High Throughput Protein Expression and Purification: Methods and Protocols; Humana Press: Totowa, NJ, USA, 2009; pp. 297–307. [Google Scholar]
- Showalter, L.E. Linear Epitopes that Generate Anti-HER-2 Antibody Responses with Trastuzuab-(Herceptin) Like Biological Activity. Ph.D. Thesis, Kent State University, Kent, OH, USA, 2014. [Google Scholar]
- Qing, G.; Ma, L.; Khorchid, A.; Swapna, G.; Mal, T.K.; Takayama, M.M.; Xia, B.; Phadtare, S.; Ke, H.; Acton, T. Cold-shock induced high-yield protein production in Escherichia coli. Nat. Biotechnol. 2004, 22, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Castiñeiras, T.S.; Williams, S.G.; Hitchcock, A.; Cole, J.A.; Smith, D.C.; Overton, T.W. Development of a generic β-lactamase screening system for improved signal peptides for periplasmic targeting of recombinant proteins in Escherichia coli. Sci. Rep. 2018, 8, 6986. [Google Scholar] [CrossRef]
- Kapust, R.B.; Tözsér, J.; Fox, J.D.; Anderson, D.E.; Cherry, S.; Copeland, T.D.; Waugh, D.S. Tobacco etch virus protease: Mechanism of autolysis and rational design of stable mutants with wild-type catalytic proficiency. Protein Eng. 2001, 14, 993–1000. [Google Scholar] [CrossRef]
- Saerens, D.; Ghassabeh, G.H.; Muyldermans, S. Single-domain antibodies as building blocks for novel therapeutics. Curr. Opin. Pharmacol. 2008, 8, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Tong, Q.; Liu, K.; Lu, X.; Shu, X.; Wang, G. Construction and characterization of a novel fusion protein MG7-scFv/SEB against gastric cancer. BioMed Res. Int. 2010, 2010, 121094. [Google Scholar] [CrossRef]
- Jurado, P.; de Lorenzo, V.; Fernández, L.A. Thioredoxin Fusions Increase Folding of Single Chain Fv Antibodies in the Cytoplasm of Escherichia coli: Evidence that Chaperone Activity is the Prime Effect of Thioredoxin. J. Mol. Biol. 2006, 357, 49–61. [Google Scholar] [CrossRef]
- Davis, G.D.; Elisee, C.; Newham, D.M.; Harrison, R.G. New fusion protein systems designed to give soluble expression in Escherichia coli. Biotechnol. Bioeng. 1999, 65, 382–388. [Google Scholar] [CrossRef]
- DelProposto, J.; Majmudar, C.Y.; Smith, J.L.; Brown, W.C. Mocr: A novel fusion tag for enhancing solubility that is compatible with structural biology applications. Protein Expr. Purif. 2009, 63, 40–49. [Google Scholar] [CrossRef]
- Nallamsetty, S.; Waugh, D.S. Mutations that alter the equilibrium between open and closed conformations of Escherichia coli maltose-binding protein impede its ability to enhance the solubility of passenger proteins. Biochem. Biophys. Res. Commun. 2007, 364, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Kapust, R.B.; Waugh, D.S. Escherichia coli maltose-binding protein is uncommonly effective at promoting the solubility of polypeptides to which it is fused. Protein Sci. 1999, 8, 1668–1674. [Google Scholar] [CrossRef]
- Wang, R.; Gu, X.; Zhuang, Z.; Zhong, Y.; Yang, H.; Wang, S. Screening and molecular evolution of a single chain variable fragment antibody (scFv) against Citreoviridin toxin. J. Agric. Food Chem. 2016, 64, 7640–7648. [Google Scholar] [CrossRef] [PubMed]
- Gąciarz, A.; Ruddock, L.W. Complementarity determining regions and frameworks contribute to the disulfide bond independent folding of intrinsically stable scFv. PLoS ONE 2017, 12, e0189964. [Google Scholar] [CrossRef] [PubMed]
- Koçer, İ.; Cox, E.C.; DeLisa, M.P.; Çelik, E. Effects of variable domain orientation on anti-HER2 single-chain variable fragment antibody expressed in the Escherichia coli cytoplasm. Biotechnol. Prog. 2021, 37, e3102. [Google Scholar] [CrossRef] [PubMed]
- Akbari, V.; Sadeghi, H.M.M.; Jafarian-Dehkordi, A.; Abedi, D.; Chou, C.P. Improved biological activity of a single chain antibody fragment against human epidermal growth factor receptor 2 (HER2) expressed in the periplasm of Escherichia coli. Protein Expr. Purif. 2015, 116, 66–74. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Construct | Total Protein 1 (mg) | Final Product 2 (mg) |
|---|---|---|
| v21 | 217 | 0.143 |
| v22 | 221 | 0.171 |
| v23 | 286 | 2.142 |
| v24 | 274 | 3.025 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byun, K.T.; Kim, B.; Cho, J.; Lee, I.; Lee, M.G.; Park, D.; Kang, T.-B.; Won, H.-S.; Kim, C.G. Development of an Anti-HER2 Single-Chain Variable Antibody Fragment Construct for High-Yield Soluble Expression in Escherichia coli and One-Step Chromatographic Purification. Biomolecules 2023, 13, 1508. https://doi.org/10.3390/biom13101508
Byun KT, Kim B, Cho J, Lee I, Lee MG, Park D, Kang T-B, Won H-S, Kim CG. Development of an Anti-HER2 Single-Chain Variable Antibody Fragment Construct for High-Yield Soluble Expression in Escherichia coli and One-Step Chromatographic Purification. Biomolecules. 2023; 13(10):1508. https://doi.org/10.3390/biom13101508
Chicago/Turabian StyleByun, Kyu Tae, Boram Kim, Junmin Cho, Inbeom Lee, Myung Gu Lee, Dongsun Park, Tae-Bong Kang, Hyung-Sik Won, and Chan Gil Kim. 2023. "Development of an Anti-HER2 Single-Chain Variable Antibody Fragment Construct for High-Yield Soluble Expression in Escherichia coli and One-Step Chromatographic Purification" Biomolecules 13, no. 10: 1508. https://doi.org/10.3390/biom13101508
APA StyleByun, K. T., Kim, B., Cho, J., Lee, I., Lee, M. G., Park, D., Kang, T. -B., Won, H. -S., & Kim, C. G. (2023). Development of an Anti-HER2 Single-Chain Variable Antibody Fragment Construct for High-Yield Soluble Expression in Escherichia coli and One-Step Chromatographic Purification. Biomolecules, 13(10), 1508. https://doi.org/10.3390/biom13101508