


A CircRNA–miRNA–mRNA Network for Exploring Doxorubicin- and Myocet-Induced Cardiotoxicity in a Translational Porcine Model

 , , , , , , ,
, , , , , , ,  and
and 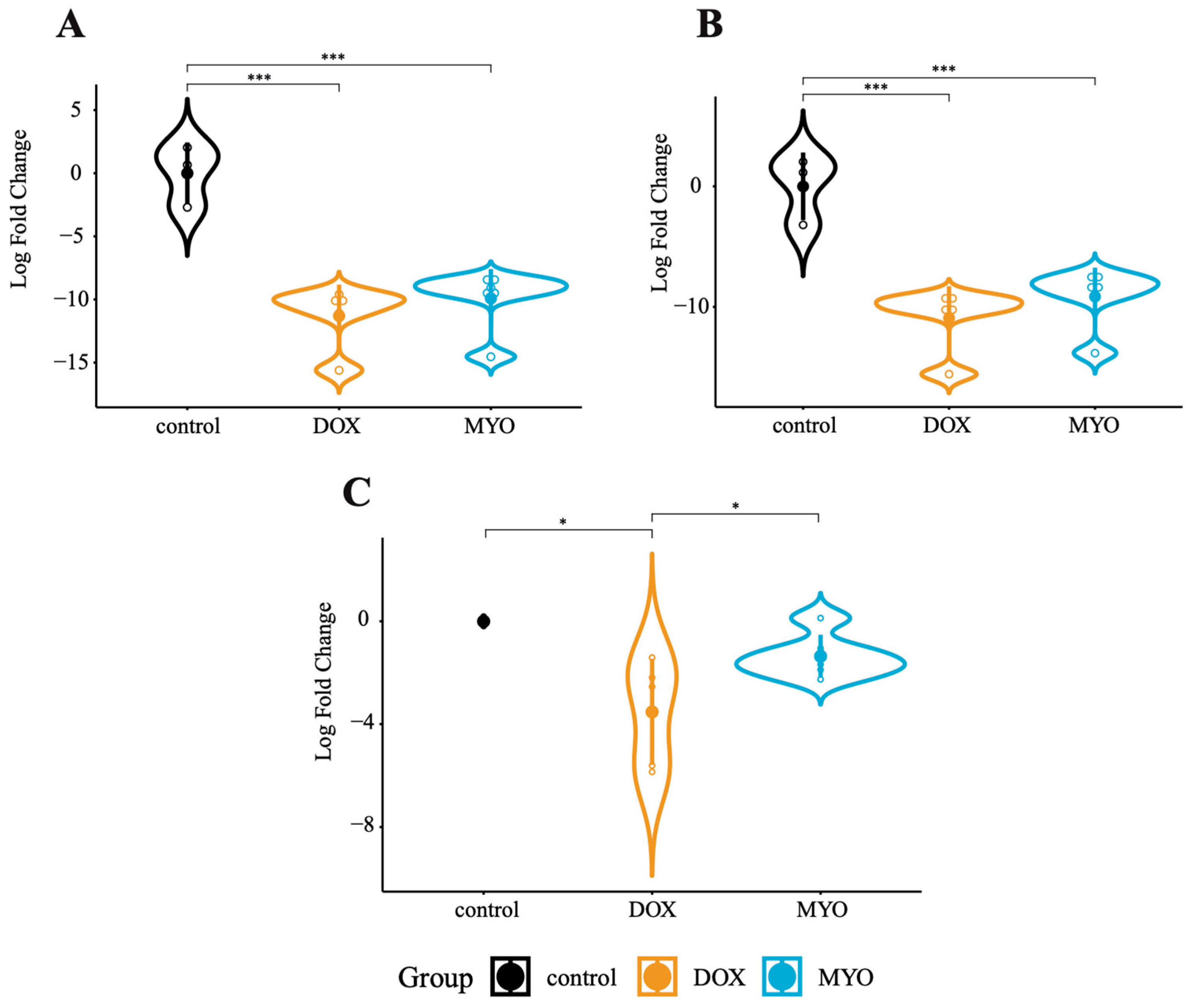{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Animal Study
2.3. RNA Extraction from Tissue
2.4. RNA Sequencing
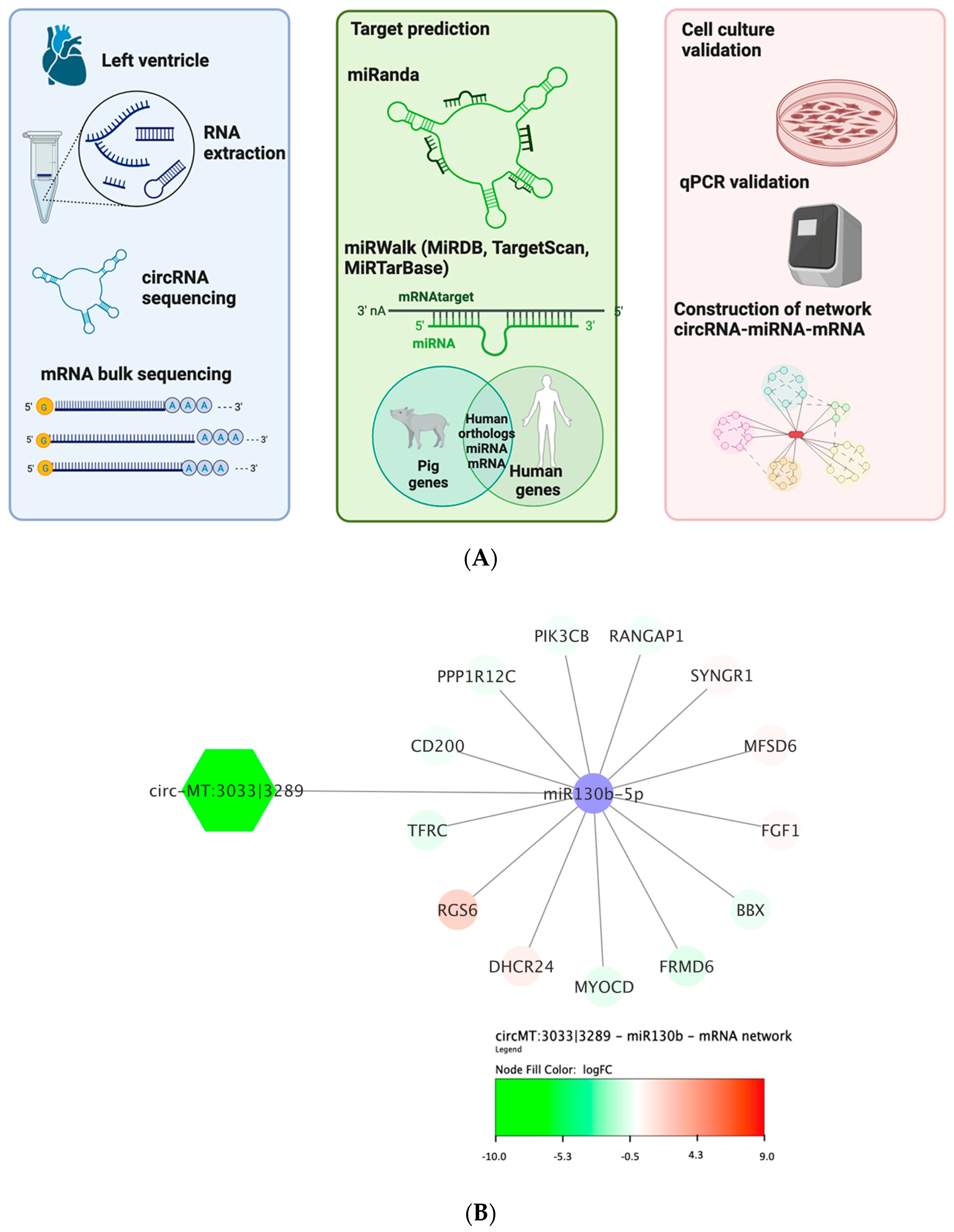
2.5. Bioinformatics
2.6. Network Analysis
2.7. Primary Cell Lineages
2.8. In Vitro Treatment of Doxorubicin and Myocet
2.9. RNA Isolation from Cells
2.10. cDNA Synthesis and Real-Time Quantitative PCR Analysis
2.11. Statistics
3. Results
3.1. Identification of circRNAs in Pig Hearts Using CIRIquant
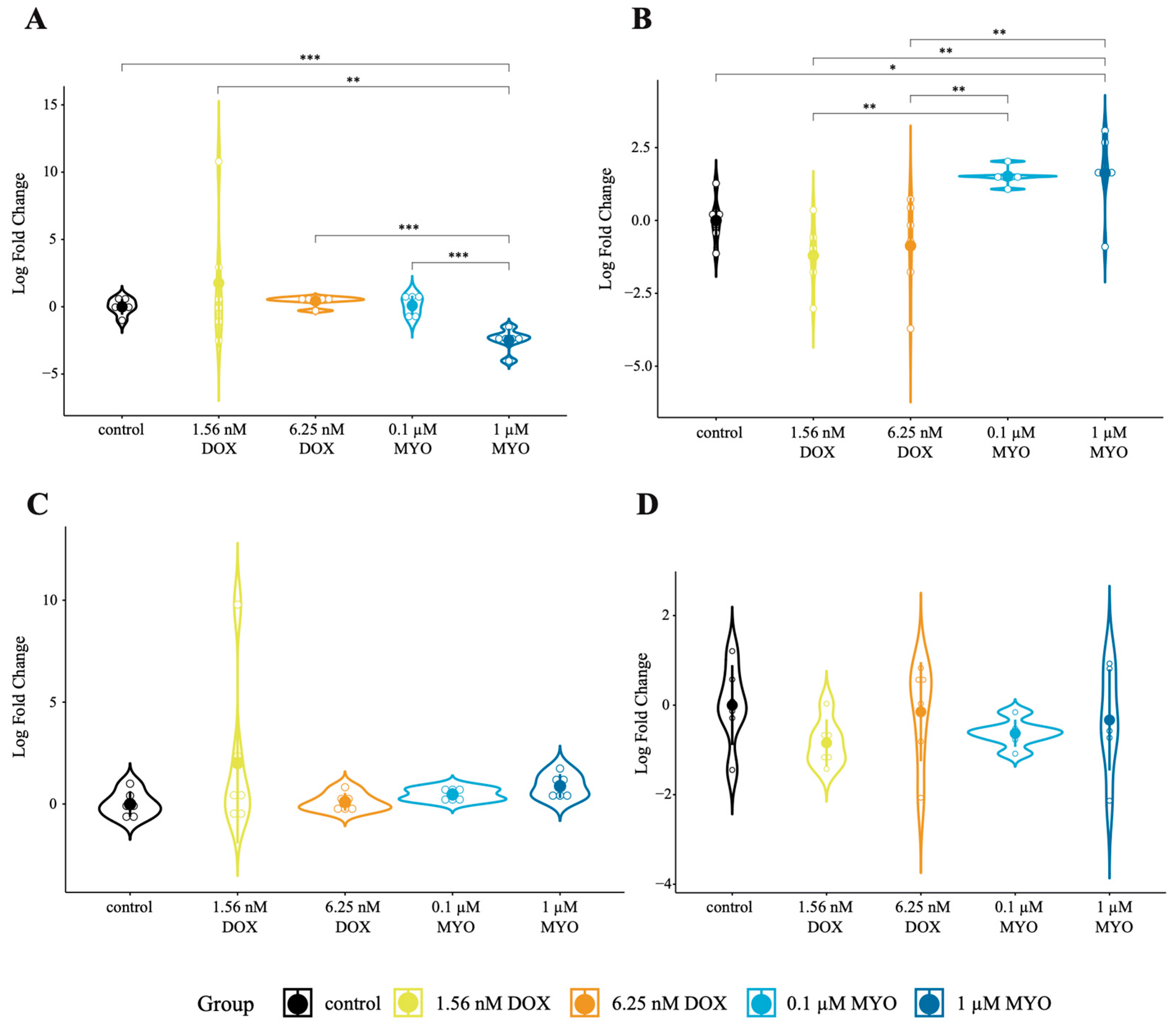
3.2. Concentration Finding of Doxorubicin and Myocet in pCPCs and pCFs In Vitro
3.3. CircRNAs in DOX and MYO-Treated pCPCs and pCFs
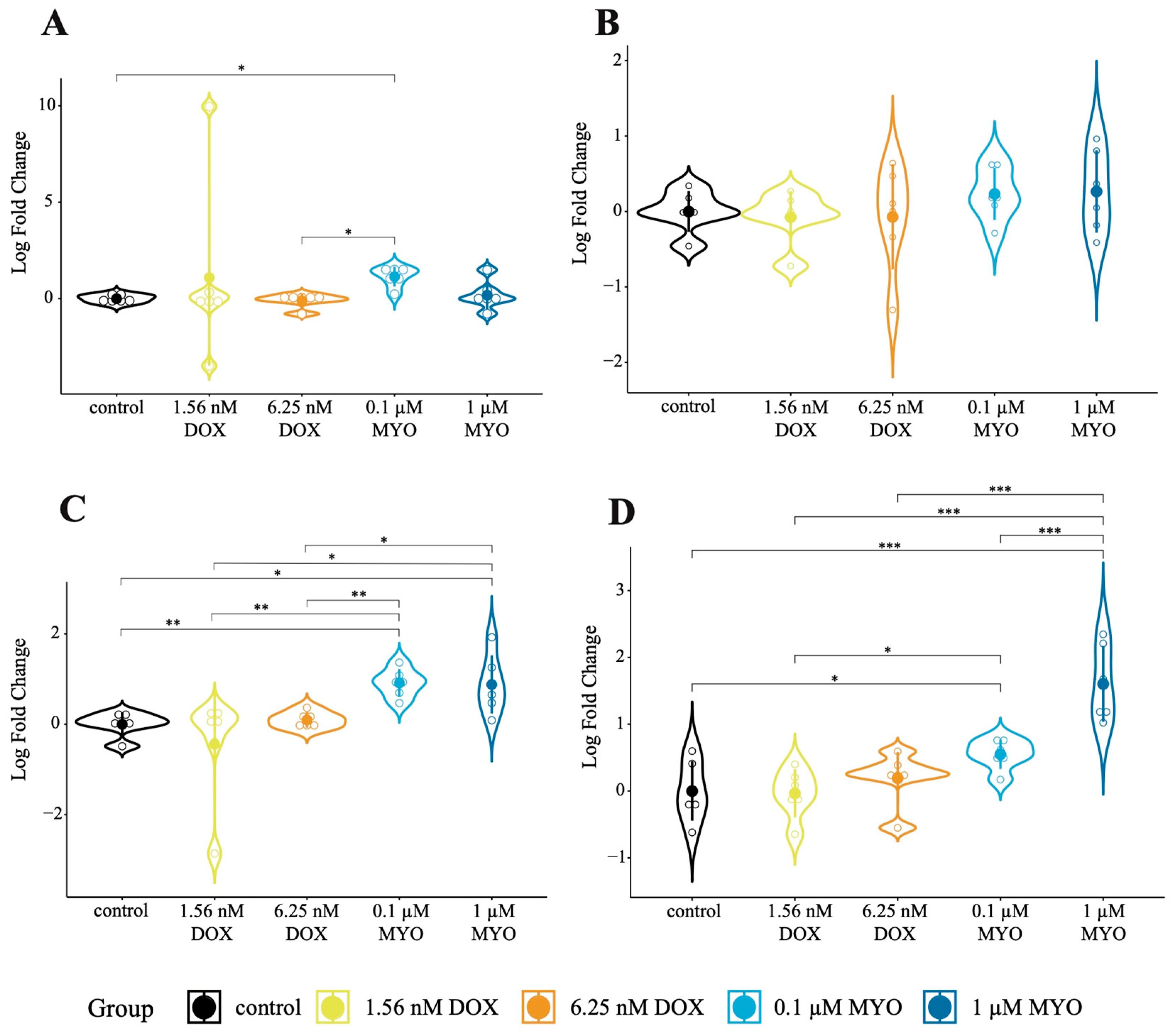
3.4. Expression of Cas3 and Ki67 in DOX- and MYO-Treated pCPCs and pCFs
3.5. CircRNAs-miRNA-Gene Network Analysis
4. Discussion
5. Conclusions
6. Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Raber, I.; Asnani, A. Cardioprotection in Cancer Therapy: Novel Insights with Anthracyclines. Cardiovasc. Res. 2019, 115, 915–921. [Google Scholar] [CrossRef]
- Sheibani, M.; Azizi, Y.; Shayan, M.; Nezamoleslami, S.; Eslami, F.; Farjoo, M.H.; Dehpour, A.R. Doxorubicin-Induced Cardiotoxicity: An Overview on Pre-Clinical Therapeutic Approaches. Cardiovasc. Toxicol. 2022, 22, 292–310. [Google Scholar] [CrossRef]
- Makwana, V.; Karanjia, J.; Haselhorst, T.; Anoopkumar-Dukie, S.; Rudrawar, S. Liposomal Doxorubicin as Targeted Delivery Platform: Current Trends in Surface Functionalization. Int. J. Pharm. 2021, 593, 120117. [Google Scholar] [CrossRef]
- Renu, K.; Abilash, V.G.; Tirupathi Pichiah, P.B.; Arunachalam, S. Molecular Mechanism of Doxorubicin-Induced Cardiomyopathy–An Update. Eur. J. Pharmacol. 2018, 818, 241–253. [Google Scholar] [CrossRef]
- Zhao, N.; Li, Q.; Sui, H.; Zhang, H. Role of Oxidation-Dependent CaMKII Activation in the Genesis of Abnormal Action Potentials in Atrial Cardiomyocytes: A Simulation Study. Biomed. Res. Int. 2020, 2020, 1597012. [Google Scholar] [CrossRef]
- Dhingra, R.; Rabinovich-Nikitin, I.; Rothman, S.; Guberman, M.; Gang, H.; Margulets, V.; Jassal, D.S.; Alagarsamy, K.N.; Dhingra, S.; Valenzuela Ripoll, C.; et al. Proteasomal Degradation of TRAF2 Mediates Mitochondrial Dysfunction in Doxorubicin-Cardiomyopathy. Circulation 2022, 146, 934–954. [Google Scholar] [CrossRef]
- Gomes, C.; Ágg, B.; Andova, A.; Arslan, S.; Baker, A.; Barteková, M.; Beis, D.; Betsou, F.; Wettinger, S.; Bugarski, B.; et al. Catalyzing Transcriptomics Research in Cardiovascular Disease: The CardioRNA COST Action CA17129. Noncoding RNA 2019, 5, 31. [Google Scholar] [CrossRef]
- Han, D.; Wang, Y.; Wang, Y.; Dai, X.; Zhou, T.; Chen, J.; Tao, B.; Zhang, J.; Cao, F. The Tumor-Suppressive Human Circular RNA CircITCH Sponges MiR-330-5p to Ameliorate Doxorubicin-Induced Cardiotoxicity through Upregulating SIRT6, Survivin, and SERCA2a. Circ. Res. 2020, 127, E108–E125. [Google Scholar] [CrossRef]
- Li, B.; Cai, X.; Wang, Y.; Zhu, H.; Zhang, P.; Jiang, P.; Yang, X.; Sun, J.; Hong, L.; Shao, L. Circ-Ska3 Enhances Doxorubicin Toxicity in Ac16 Cells through Mir-1303/Tlr4 Axis. Int. Heart J. 2021, 62, 1112–1123. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Liu, J.; Deng, H.; Ma, R.; Liao, J.-Y.; Liang, H.; Hu, J.; Li, J.; Guo, Z.; Cai, J.; et al. Targeting Mitochondria-Located CircRNA SCAR Alleviates NASH via Reducing MROS Output. Cell 2020, 183, 76–93.e22. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Liu, T.; Bao, Y.; Zhao, T.; Wang, J.; Wang, H.; Wang, A.; Gan, X.; Wu, Z.; Wang, L. CircRNA CRAPGEF5 Inhibits the Growth and Metastasis of Renal Cell Carcinoma via the MiR-27a-3p/TXNIP Pathway. Cancer Lett. 2020, 469, 68–77. [Google Scholar] [CrossRef]
- Zeng, K.; Chen, X.; Xu, M.; Liu, X.; Hu, X.; Xu, T.; Sun, H.; Pan, Y.; He, B.; Wang, S. CircHIPK3 Promotes Colorectal Cancer Growth and Metastasis by Sponging MiR-7. Cell Death Dis. 2018, 9, 417. [Google Scholar] [CrossRef]
- Zheng, X.; Huang, M.; Xing, L.; Yang, R.; Wang, X.; Jiang, R.; Zhang, L.; Chen, J. The CircRNA CircSEPT9 Mediated by E2F1 and EIF4A3 Facilitates the Carcinogenesis and Development of Triple-Negative Breast Cancer. Mol. Cancer 2020, 19, 73. [Google Scholar] [CrossRef]
- Karousi, P.; Artemaki, P.I.; Sotiropoulou, C.D.; Christodoulou, S.; Scorilas, A.; Kontos, C.K. Identification of Two Novel Circular RNAs Deriving from BCL2L12 and Investigation of Their Potential Value as a Molecular Signature in Colorectal Cancer. Int. J. Mol. Sci. 2020, 21, 8867. [Google Scholar] [CrossRef]
- Ji, W.; Qiu, C.; Wang, M.; Mao, N.; Wu, S.; Dai, Y. Hsa_circ_0001649: A Circular RNA and Potential Novel Biomarker for Colorectal Cancer. Biochem. Biophys. Res. Commun. 2018, 497, 122–126. [Google Scholar] [CrossRef]
- Ji, X.; Ding, W.; Xu, T.; Zheng, X.; Zhang, J.; Liu, M.; Liu, G.; Wang, J. MicroRNA-31-5p Attenuates Doxorubicin-Induced Cardiotoxicity via Quaking and Circular RNA Pan3. J. Mol. Cell Cardiol. 2020, 140, 56–67. [Google Scholar] [CrossRef]
- Lu, D.; Chatterjee, S.; Xiao, K.; Riedel, I.; Huang, C.-K.; Costa, A.; Cushman, S.; Neufeldt, D.; Rode, L.; Schmidt, A.; et al. A Circular RNA Derived from the Insulin Receptor Locus Protects against Doxorubicin-Induced Cardiotoxicity. Eur. Heart J. 2022, 43, 4496–4511. [Google Scholar] [CrossRef]
- Makkos, A.; Ágg, B.; Petrovich, B.; Varga, Z.V.; Görbe, A.; Ferdinandy, P. Systematic Review and Network Analysis of MicroRNAs Involved in Cardioprotection against Myocardial Ischemia/Reperfusion Injury and Infarction: Involvement of Redox Signalling. Free Radic. Biol. Med. 2021, 172, 237–251. [Google Scholar] [CrossRef]
- Perrino, C.; Barabási, A.-L.; Condorelli, G.; Davidson, S.M.; De Windt, L.; Dimmeler, S.; Engel, F.B.; Hausenloy, D.J.; Hill, J.A.; Van Laake, L.W.; et al. Epigenomic and Transcriptomic Approaches in the Post-Genomic Era: Path to Novel Targets for Diagnosis and Therapy of the Ischaemic Heart? Position Paper of the European Society of Cardiology Working Group on Cellular Biology of the Heart. Cardiovasc. Res. 2017, 113, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Guide for the Care and Use of Laboratory Animals; National Academies Press: Washington, DC, USA, 2011; ISBN 978-0-309-15400-0.
- Gyöngyösi, M.; Lukovic, D.; Zlabinger, K.; Spannbauer, A.; Gugerell, A.; Pavo, N.; Traxler, D.; Pils, D.; Maurer, G.; Jakab, A.; et al. Liposomal Doxorubicin Attenuates Cardiotoxicity via Induction of Interferon-Related DNA Damage Resistance. Cardiovasc. Res. 2019, 116, 970–982. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, S.; Yang, J.; Zhao, F. Accurate Quantification of Circular RNAs Identifies Extensive Circular Isoform Switching Events. Nat. Commun. 2020, 11, 90. [Google Scholar] [CrossRef]
- John, B.; Enright, A.J.; Aravin, A.; Tuschl, T.; Sander, C.; Marks, D.S. Human MicroRNA Targets. PLoS Biol. 2004, 2, e363. [Google Scholar] [CrossRef]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. MiRWalk: An Online Resource for Prediction of MicroRNA Binding Sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. G:Profiler: A Web Server for Functional Enrichment Analysis and Conversions of Gene Lists (2019 Update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef]
- Zlabinger, K.; Spannbauer, A.; Traxler, D.; Gugerell, A.; Lukovic, D.; Winkler, J.; Mester-Tonczar, J.; Podesser, B.; Gyöngyösi, M. MiR-21, MiR-29a, GATA4, and MEF2c Expression Changes in Endothelin-1 and Angiotensin II Cardiac Hypertrophy Stimulated Isl-1+Sca-1+c-Kit+ Porcine Cardiac Progenitor Cells In Vitro. Cells 2019, 8, 1416. [Google Scholar] [CrossRef]
- Toldo, S.; Goehe, R.W.; Lotrionte, M.; Mezzaroma, E.; Sumner, E.T.; Biondi-Zoccai, G.G.L.; Seropian, I.M.; Van Tassell, B.W.; Loperfido, F.; Palazzoni, G.; et al. Comparative Cardiac Toxicity of Anthracyclines In Vitro and In Vivo in the Mouse. PLoS ONE 2013, 8, e58421. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, Z.; Xu, J.; Feng, M.; Zhang, H.; Zhang, L.; Qian, L. Circular RNA Arhgap12 Modulates Doxorubicin-Induced Cardiotoxicity by Sponging MiR-135a-5p. Life Sci. 2021, 265, 118788. [Google Scholar] [CrossRef]
- Kciuk, M.; Gielecińska, A.; Mujwar, S.; Kołat, D.; Kałuzińska-Kołat, Ż.; Celik, I.; Kontek, R. Doxorubicin—An Agent with Multiple Mechanisms of Anticancer Activity. Cells 2023, 12, 659. [Google Scholar] [CrossRef]
- Everaert, C.; Luypaert, M.; Maag, J.L.V.; Cheng, Q.X.; Dinger, M.E.; Hellemans, J.; Mestdagh, P. Benchmarking of RNA-Sequencing Analysis Workflows Using Whole-Transcriptome RT-QPCR Expression Data. Sci. Rep. 2017, 7, 1559. [Google Scholar] [CrossRef]
- Coenye, T. Do Results Obtained with RNA-Sequencing Require Independent Verification? Biofilm 2021, 3, 100043. [Google Scholar] [CrossRef]
- Coban, N.; Ozuynuk, A.S.; Erkan, A.F.; Guclu-Geyik, F.; Ekici, B. Levels of MiR-130b-5p in Peripheral Blood Are Associated with Severity of Coronary Artery Disease. Mol. Biol. Rep. 2021, 48, 7719–7732. [Google Scholar] [CrossRef]
- Li, B.; Zhang, H. Knockdown of microRNA-130b improves doxorubicin sensitivity in bladder urothelial carcinoma by negatively regulating cylindromatosis expression. Arch. Med. Sci. 2021, 17, 1038–1043. [Google Scholar] [CrossRef]
- Zou, M.; Huang, C.; Li, X.; He, X.; Chen, Y.; Liao, W.; Liao, Y.; Sun, J.; Liu, Z.; Zhong, L.; et al. Circular RNA Expression Profile and Potential Function of Hsa_circRNA_101238 in Human Thoracic Aortic Dissection. Oncotarget 2017, 8, 81825–81837. [Google Scholar] [CrossRef]
- Li, B.; Zhu, L.; Lu, C.; Wang, C.; Wang, H.; Jin, H.; Ma, X.; Cheng, Z.; Yu, C.; Wang, S.; et al. CircNDUFB2 Inhibits Non-Small Cell Lung Cancer Progression via Destabilizing IGF2BPs and Activating Anti-Tumor Immunity. Nat. Commun. 2021, 12, 295. [Google Scholar] [CrossRef]
- Wu, W.; Zhou, J.; Wu, Y.; Tang, X.; Zhu, W. Overexpression of CircRNA CircFAT1 in Endometrial Cancer Cells Increases Their Stemness by Upregulating MiR-21 Through Methylation. Cancer Biother. Radiopharm. 2021, 37, 843–849. [Google Scholar] [CrossRef]
- Yu, M.-C.; Ding, G.-Y.; Ma, P.; Chen, Y.-D.; Zhu, X.-D.; Cai, J.-B.; Shen, Y.-H.; Zhou, J.; Fan, J.; Sun, H.-C.; et al. CircRNA UBAP2 Serves as a Sponge of MiR-1294 to Increase Tumorigenesis in Hepatocellular Carcinoma through Regulating c-Myc Expression. Carcinogenesis 2021, 42, 1293–1303. [Google Scholar] [CrossRef]
- Wu, Z.; Sun, H.; Wang, C.; Liu, W.; Liu, M.; Zhu, Y.; Xu, W.; Jin, H.; Li, J. Mitochondrial Genome-Derived CircRNA Mc-COX2 Functions as an Oncogene in Chronic Lymphocytic Leukemia. Mol. Ther. Nucleic Acids 2020, 20, 801–811. [Google Scholar] [CrossRef]
- Sun, W.; Lu, Y.; Zhang, H.; Zhang, J.; Fang, X.; Wang, J.; Li, M. Mitochondrial Non-Coding RNAs Are Potential Mediators of Mitochondrial Homeostasis. Biomolecules 2022, 12, 1863. [Google Scholar] [CrossRef]
- Tadokoro, T.; Ikeda, M.; Ide, T.; Deguchi, H.; Ikeda, S.; Okabe, K.; Ishikita, A.; Matsushima, S.; Koumura, T.; Yamada, K.; et al. Mitochondria-Dependent Ferroptosis Plays a Pivotal Role in Doxorubicin Cardiotoxicity. JCI Insight 2020, 5, e132747. [Google Scholar] [CrossRef]
- Ichikawa, Y.; Ghanefar, M.; Bayeva, M.; Wu, R.; Khechaduri, A.; Prasad, S.V.N.; Mutharasan, R.K.; Naik, T.J.; Ardehali, H. Cardiotoxicity of Doxorubicin Is Mediated through Mitochondrial Iron Accumulation. J. Clin. Investig. 2014, 124, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, X.; Li, J.; Hu, S.; Deng, Y.; Yin, H.; Bao, X.; Zhang, Q.C.; Wang, G.; Wang, B.; et al. Identification of MecciRNAs and Their Roles in the Mitochondrial Entry of Proteins. Sci. China Life Sci. 2020, 63, 1429–1449. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, Y.; Deng, Z.; Wang, Y.; Zheng, Y.; Jiang, W.; Jiang, L. MicroRNA Expression Profiling Involved in Doxorubicin-induced Cardiotoxicity Using High-throughput Deep-sequencing Analysis. Oncol. Lett. 2021, 22, 560. [Google Scholar] [CrossRef]
- Hevener, K.; Verstak, T.A.; Lutat, K.E.; Riggsbee, D.L.; Mooney, J.W. Recent Developments in Topoisomerase-Targeted Cancer Chemotherapy. Acta Pharm. Sin. B 2018, 8, 844–861. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mester-Tonczar, J.; Einzinger, P.; Hasimbegovic, E.; Kastner, N.; Schweiger, V.; Spannbauer, A.; Han, E.; Müller-Zlabinger, K.; Traxler-Weidenauer, D.; Bergler-Klein, J.; et al. A CircRNA–miRNA–mRNA Network for Exploring Doxorubicin- and Myocet-Induced Cardiotoxicity in a Translational Porcine Model. Biomolecules 2023, 13, 1711. https://doi.org/10.3390/biom13121711
Mester-Tonczar J, Einzinger P, Hasimbegovic E, Kastner N, Schweiger V, Spannbauer A, Han E, Müller-Zlabinger K, Traxler-Weidenauer D, Bergler-Klein J, et al. A CircRNA–miRNA–mRNA Network for Exploring Doxorubicin- and Myocet-Induced Cardiotoxicity in a Translational Porcine Model. Biomolecules. 2023; 13(12):1711. https://doi.org/10.3390/biom13121711
Chicago/Turabian StyleMester-Tonczar, Julia, Patrick Einzinger, Ena Hasimbegovic, Nina Kastner, Victor Schweiger, Andreas Spannbauer, Emilie Han, Katrin Müller-Zlabinger, Denise Traxler-Weidenauer, Jutta Bergler-Klein, and et al. 2023. "A CircRNA–miRNA–mRNA Network for Exploring Doxorubicin- and Myocet-Induced Cardiotoxicity in a Translational Porcine Model" Biomolecules 13, no. 12: 1711. https://doi.org/10.3390/biom13121711
APA StyleMester-Tonczar, J., Einzinger, P., Hasimbegovic, E., Kastner, N., Schweiger, V., Spannbauer, A., Han, E., Müller-Zlabinger, K., Traxler-Weidenauer, D., Bergler-Klein, J., Gyöngyösi, M., & Lukovic, D. (2023). A CircRNA–miRNA–mRNA Network for Exploring Doxorubicin- and Myocet-Induced Cardiotoxicity in a Translational Porcine Model. Biomolecules, 13(12), 1711. https://doi.org/10.3390/biom13121711