
Metabolites and Bioactivity of the Marine Xestospongia Sponges (Porifera, Demospongiae, Haplosclerida) of Southeast Asian Waters

 , , and
, , and
Abstract
:1. Introduction
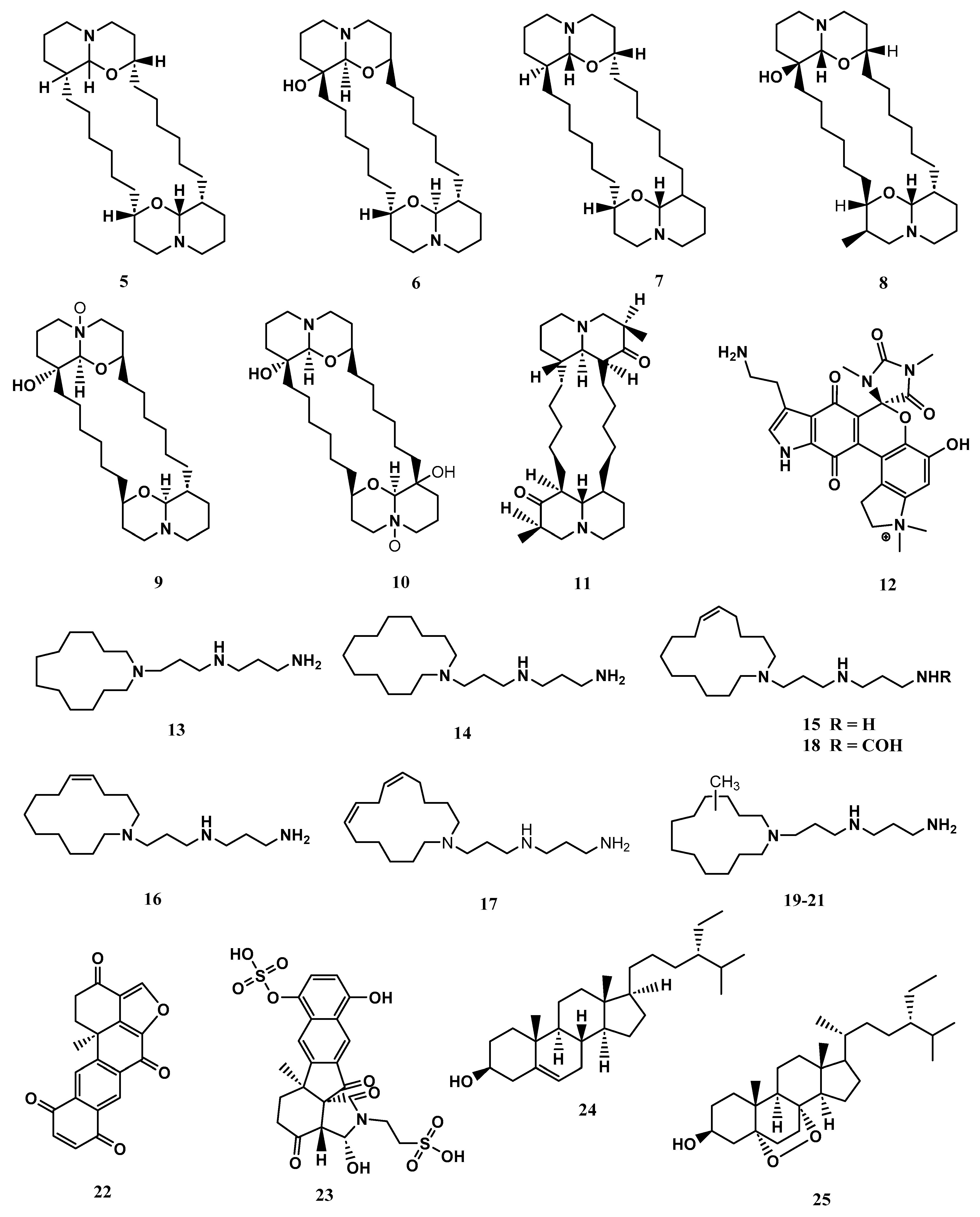
2. Secondary Metabolites in Xestospongia spp. and Their Bioactivity
2.1. Xestospongia bergquistia (Fromont, 1991)
2.2. Xestospongia muta (Schmidt, 1870)
2.3. Xestospongia exigua (Kirkpatrick, 1900)
2.4. Xestospongia testudinaria (Lamarck, 1815)
2.5. Xestospongia ashmorica (Hooper, 1984)
2.6. Xestospongia vansoesti (Bakus & Nishiyama, 2000)
3. Summary of Xestospongia Metabolites
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hooper, J.N.A.; Van Soest, R.W.M. Systema Porifera. A guide to the classification of sponges. In Systema Porifera; Springer: Berlin/Heidelberg, Germany, 2002; pp. 1–7. [Google Scholar]
- Van Soest, R.W.M.; Boury-Esnault, N.; Vacelet, J.; Dohrmann, M.; Erpenbeck, D.; De Voogd, N.J.; Santodomingo, N.; Vanhoorne, B.; Kelly, M.; Hooper, J.N.A. Global diversity of sponges (Porifera). PLoS ONE 2012, 7, e35105. [Google Scholar] [CrossRef] [PubMed]
- Laport, M.S.; Santos, O.C.S.; Muricy, G. Marine sponges: Potential sources of new antimicrobial drugs. Curr. Pharm. Biotechnol. 2009, 10, 86–105. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.E.; Johnson, T.W. First total synthesis of xestobergsterol A and active structural analogues of the xestobergsterols. Tetrahedron 2001, 57, 1449–1481. [Google Scholar] [CrossRef]
- Kobayashi, J.; Shinonaga, H.; Shigemori, H.; Umeyama, A.; Shoji, N.; Arihara, S. Xestobergsterol C, a new pentacyclic steroid from the okinawan marine sponge ircinia sp. and absolute stereochemistry of xestobergsterol A. J. Nat. Prod. 1995, 58, 312–318. [Google Scholar] [CrossRef]
- Shoji, N.; Umeyama, A.; Shin, K.; Takeda, K.; Arihara, S.; Kobayashi, J.; Takei, M. Two Unique Pentacyclic Steroids with Cis C/D Ring Junction from Xestospongia bergquistia Fromont, Powerful Inhibitors of Histamine Release. J. Org. Chem. 1992, 57, 2996–2997. [Google Scholar] [CrossRef]
- Ye, J.; Zhou, F.; Al-Kareef, A.M.Q.; Wang, H. Anticancer agents from marine sponges. J. Asian Nat. Prod. Res. 2015, 17, 64–88. [Google Scholar] [CrossRef]
- Mehbub, M.F.; Perkins, M.V.; Zhang, W.; Franco, C.M.M. New marine natural products from sponges (Porifera) of the order Dictyoceratida (2001 to 2012); a promising source for drug discovery, exploration and future prospects. Biotechnol. Adv. 2016, 34, 473–491. [Google Scholar] [CrossRef]
- Mehbub, M.F.; Lei, J.; Franco, C.; Zhang, W. Marine sponge derived natural products between 2001 and 2010: Trends and opportunities for discovery of bioactives. Mar. Drugs 2014, 12, 4539–4577. [Google Scholar] [CrossRef] [Green Version]
- Swierts, T.; Peijnenburg, K.T.C.A.; de Leeuw, C.; Cleary, D.F.R.; Hörnlein, C.; Setiawan, E.; Wörheide, G.; Erpenbeck, D.; de Voogd, N.J. Lock, stock and two different barrels: Comparing the genetic composition of morphotypes of the Indo-Pacific sponge Xestospongia testudinaria. PLoS ONE 2013, 8, e74396. [Google Scholar] [CrossRef]
- López-Legentil, S.; Pawlik, J.R. Genetic structure of the Caribbean giant barrel sponge Xestospongia muta using the I3-M11 partition of COI. Coral Reefs 2009, 28, 157–165. [Google Scholar] [CrossRef]
- Zea, S.; Rutzler, K. A new species of Xestospongia (Porifera: Demospongea) from the Colombian Caribbean. Caldasia 1983, 13, 817–831. [Google Scholar]
- De Carvalho, S.M.; Lopes, D.A.; Cosme, B.; Hajdu, E. Seven new species of sponges (Porifera) from deep-sea coral mounds at Campos Basin (SW Atlantic). Helgol. Mar. Res. 2016, 70, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Xu, T.; Yang, X.W.; Huang, R.; Yang, B.; Tang, L.; Liu, Y. Chemical and biological aspects of marine sponges of the genus Xestospongia. Chem. Biodivers. 2010, 7, 2201–2227. [Google Scholar] [CrossRef]
- Setiawan, E.; de Voogd, N.J.; Swierts, T.; Hooper, J.N.A.; Wörheide, G.; Erpenbeck, D. MtDNA diversity of the Indonesian giant barrel sponge Xestospongia testudinaria (Porifera: Haplosclerida)—Implications from partial cytochrome oxidase 1 sequences. J. Mar. Biol. Assoc. U. K. 2016, 96, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Fromont, J. Descriptions of species of the Petrosida (Porifera: Demospongiae) occurring in the tropical waters of the Great Barrier Reef. Beagle Rec. Museums Art Gall. North. Territ. 1991, 8, 73–95. [Google Scholar] [CrossRef]
- De Monte Lamarck, J.B.P. Suite des polypiers empâtés. Mémoirs du Muséum d’Histoire naturelle. Paris 1815, 1, 69–80. [Google Scholar]
- Schmidt, E.O. Grundzüge Einer Spongien-Fauna des Atlantischen Gebietes; Hansebooks: Norderstedt, Germany, 1870. [Google Scholar]
- Kirkpatrick, R. On the Sponges of Christmas Island. Proc. Zoolological Soc. London 1900, 68, 127–141. [Google Scholar]
- Hooper, J.N.A. A New Genus and Two New Species of Haplosclerid Sponges (Porifera: Demospongiae) from the Timor Sea, Northwest Australia; Stillwell and Co.: Melbourne, Australia, 1984. [Google Scholar]
- Bakus, G.J.; Nishiyama, G.K. Three species of toxic sponges from Cebu, Philippines (Porifera: Demospongiae). Proc. Biol. Soc. Washingt. 2000, 113, 1162–1172. [Google Scholar]
- Darumas, U.; Chavanich, S.; Suwanborirux, K. Distribution Patterns of the Renieramycin-Producing Sponge, Xestospongia sp., and Its Association with Other Reef Organisms in the Gulf of Thailand. Zool. Stud. 2007, 46, 695–704. [Google Scholar]
- Swierts, T.; Cleary, D.F.R.; de Voogd, N.J. Prokaryotic communities of Indo-Pacific giant barrel sponges are more strongly influenced by geography than host phylogeny. FEMS Microbiol. Ecol. 2018, 94, fiy194. [Google Scholar] [CrossRef] [Green Version]
- Chong, W.S.; Zaki, N.H.M.; Hossain, M.S.; Muslim, A.M.; Pour, A.B. Introducing Theil-Sen estimator for sun glint correction of UAV data for coral mapping. Geocarto Int. 2021, 37, 4527–4556. [Google Scholar] [CrossRef]
- Safuan, C.D.M.; Ismail, K.; Khalil, I.; Ali, A.; Chong, W.S.; Chan, A.A.; Ismail, M.N.; Repin, I.M.; Bachok, Z. Quantification of coral reef benthos for coral health assessment in Labuan Marine Park, Malaysia. J. Sustain. Sci. Manag. 2018, 13, 101–112. [Google Scholar]
- Zaki, N.H.M.; Chong, W.S.; Muslim, A.M.; Reba, M.N.M.; Hossain, M.S. Assessing optimal UAV-data pre-processing workflows for quality ortho-image generation to support coral reef mapping. Geocarto Int. 2022, 37, 1–25. [Google Scholar] [CrossRef]
- McGrath, E. Demography and Impacts of Habitat Degradation on the Giant Barrel Sponge Xestospongia spp. in the Indo-Pacific. Doctoral Thesis, Victoria University of Wellington, Wellington, New Zealand, 2018. [Google Scholar]
- Schönberg, C.H.L. No taxonomy needed: Sponge functional morphologies inform about environmental conditions. Ecol. Indic. 2021, 129, 107806. [Google Scholar] [CrossRef]
- Marty, M.J.; Vicente, J.; Oyler, B.L.; Place, A.; Hill, R.T. Sponge symbioses between Xestospongia deweerdtae and Plakortis spp. are not motivated by shared chemical defense against predators. PLoS ONE 2017, 12, e0174816. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.K.; Lee, J.H.; Lee, H.K. Microbial Symbiosis in Marine Sponges. J. Microbiol. 2001, 39, 254–264. [Google Scholar]
- Taylor, M.W.; Radax, R.; Steger, D.; Wagner, M. Sponge-Associated Microorganisms: Evolution, Ecology, and Biotechnological Potential. Microbiol. Mol. Biol. Rev. 2007, 71, 295–347. [Google Scholar] [CrossRef] [Green Version]
- Wulff, J.L. Rapid diversity and abundance decline in a Caribbean coral reef sponge community. Biol. Conserv. 2006, 127, 167–176. [Google Scholar] [CrossRef]
- Wulff, J.L. Disease prevalence and population density over time in three common Caribbean coral reef sponge species. J. Mar. Biol. Assoc. U. K. 2007, 87, 1715–1720. [Google Scholar] [CrossRef] [Green Version]
- Cowart, J.D.; Henkel, T.P.; McMurray, S.E.; Pawlik, J.R. Sponge orange band (SOB): A pathogenic-like condition of the giant barrel sponge, Xestospongia muta. Coral Reefs 2006, 25, 513. [Google Scholar] [CrossRef]
- Angermeier, H.; Kamke, J.; Abdelmohsen, U.R.; Krohne, G.; Pawlik, J.R.; Lindquist, N.L.; Hentschel, U. The pathology of sponge orange band disease affecting the Caribbean barrel sponge Xestospongia muta. FEMS Microbiol. Ecol. 2011, 75, 218–230. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Hernandez, J.E.; Tuohy, E.; Toledo-Rodríguez, D.A.; Sherman, C.; Schizas, N.V.; Weil, E. Detrimental conditions affecting Xestospongia muta across shallow and mesophotic coral reefs off the southwest coast of Puerto Rico. Dis. Aquat. Organ. 2021, 147, 47–61. [Google Scholar] [CrossRef]
- López-Legentil, S.; Song, B.; Mcmurray, S.E.; Pawlik, J.R. Bleaching and stress in coral reef ecosystems: hsp70 expression by the giant barrel sponge Xestospongia muta. Mol. Ecol. 2008, 17, 1840–1849. [Google Scholar] [CrossRef]
- Keumala, S.; Illahi, G.F.; Sakinah, R.; Razi, N.M.; Khairunnisa, K.; Kurnianda, V. Bioactivity of Indonesian’s Marine Sponge Xestospongia muta as Antidormant Mycobacterium smegmatis. Med. Chem. 2018, 8, 9. [Google Scholar] [CrossRef]
- Nagasawa, Y.; Ueoka, R.; Yamanokuchi, R.; Horiuchi, N.; Ikeda, T.; Rotinsulu, H.; Mangindaan, R.E.P.; Ukai, K.; Kobayashi, H.; Namikoshi, M.; et al. Isolation of salsolinol, a tetrahydroisoquinoline alkaloid, from the marine sponge Xestospongia cf. vansoesti as a proteasome inhibitor. Chem. Pharm. Bull. 2011, 59, 287–290. [Google Scholar] [CrossRef] [Green Version]
- Abdillah, S.; Nurhayati, A.P.D.; Nurhatika, S.; Setiawan, E.; Heffen, W.L. Cytotoxic and antioxidant activities of marine sponge diversity at Pecaron Bay Pasir Putih Situbondo East Java, Indonesia. J. Pharm. Res. 2013, 6, 685–689. [Google Scholar] [CrossRef]
- Rahmanisa, S.; Prajatelistia, E.; Wibowo, I.; Barlian, A. 3D Biosilica Scaffolds from Melophlus sarasinorum and Xestospongia testudinaria Indonesian Sponges are Biocompatible for Cell Growth and Differentiation of Human Wharton’s Jelly Mesenchymal Stem Cell in Bone Tissue Engineering. Indones. Biomed. J. 2022, 14, 382–392. [Google Scholar] [CrossRef]
- Edrada, R.A.; Proksch, P.; Wray, V.; Witte, L.; Müller, W.E.G.; Van Soest, R.W.M. Four new bioactive manzamine-type alkaloids from the Philippine marine sponge Xestospongia ashmorica. J. Nat. Prod. 1996, 59, 1056–1060. [Google Scholar] [CrossRef] [Green Version]
- Quah, Y.; Mohd Ismail, N.I.; Ooi, J.L.S.; Affendi, Y.A.; Abd Manan, F.; Wong, F.C.; Chai, T.T. Identification of Novel Cytotoxic Peptide KENPVLSLVNGMF from Marine Sponge Xestospongia testudinaria, with Characterization of Stability in Human Serum. Int. J. Pept. Res. Ther. 2018, 24, 189–199. [Google Scholar] [CrossRef]
- Azemi, N.A.; Azemi, A.K.; Abu-Bakar, L.; Sevakumaran, V.; Muhammad, T.S.T.; Ismail, N. Xestospongia muta Fraction-7 and Linoleic Acid: Effects on SR-BI Gene Expression and HDL Cholesterol Uptake. Mar. Drugs 2022, 20, 762. [Google Scholar] [CrossRef]
- Qaralleh, H.; Idid, S.; Saad, S.; Susanti, D.; Taher, M.; Khleifat, K. Antifungal and Antibacterial Activities of Four Malaysian Sponge Species (Petrosiidae). J. Mycol. Med. 2010, 20, 315–320. [Google Scholar] [CrossRef]
- Putchakarn, S. Species diversity of marine sponges along Chanthaburi and Trat Provinces, the eastern coast of the Gulf of Thailand. Publ. Seto Mar. Biol. Lab. 2011, 41, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Azzini, F.; Calcinai, B.; Cerrano, C.; Bavestrello, G.; Pansini, M. Sponges of the marine karst lakes and of the coast of the islands of Ha Long Bay (North Vietnam). Porifera Res. Biodevers. Innovat. Sustain. 2007, 157–164. [Google Scholar]
- Nguyen, X.C.; Longeon, A.; Pham, V.C.; Urvois, F.; Bressy, C.; Van Trinh, T.T.; Nguyen, H.N.; Phan, V.K.; Chau, V.M.; Briand, J.-F.; et al. Antifouling 26,27-cyclosterols from the Vietnamese marine sponge Xestospongia testudinaria. J. Nat. Prod. 2013, 76, 1313–1318. [Google Scholar] [CrossRef] [PubMed]
- Aiello, A. Steroids from sponges: Recent reports. Steroids 1999, 64, 687–714. [Google Scholar] [CrossRef]
- Terracciano, S.; Aquino, M.; Rodriquez, M.; Chiara Monti, M.; Casapullo, A.; Riccio, R.; Gomez-Paloma, L. Chemistry and Biology of Anti-Inflammatory Marine Natural Products: Molecules Interfering with Cyclooxygenase, NF-kB and Other Unidentified Targets. Curr. Med. Chem. 2006, 13, 1947–1969. [Google Scholar] [CrossRef]
- Leone, P.D.A.; Carroll, A.R.; Towerzey, L.; King, G.; McArdle, B.M.; Kern, G.; Fisher, S.; Hooper, J.N.A.; Quinn, R.J. Exiguaquinol: A novel pentacyclic hydroquinone from Neopetrosia exigua that inhibits Helicobacter pylori MurI. Org. Lett. 2008, 10, 2585–2588. [Google Scholar] [CrossRef]
- Roll, D.M.; Scheuer, P.J.; Matsumoto, G.K.; Clardy, J. Halenaquinone, a Pentacyclic Polyketide from a Marine Sponge. J. Am. Chem. Soc. 1983, 105, 6177–6178. [Google Scholar] [CrossRef]
- Cerqueira, F.; Watanadilok, R.; Sonchaeng, P.; Kijjoa, A.; Pinto, M.; Van Ufford, H.Q.; Kroes, B.; Beukelman, C.; Nascimento, M.S.J. Clionasterol: A potent inhibitor of complement component C1. Planta Med. 2003, 69, 174–176. [Google Scholar] [CrossRef]
- Zhu, S.; Jiao, W.; Xu, Y.; Hou, L.; Li, H.; Shao, J.; Zhang, X.; Wang, R.; Kong, D. Palmitic acid inhibits prostate cancer cell proliferation and metastasis by suppressing the PI3K/Akt pathway. Life Sci. 2021, 286, 120046. [Google Scholar] [CrossRef]
- De Araujo Junior, R.F.; Eich, C.; Jorquera, C.; Schomann, T.; Baldazzi, F.; Chan, A.B.; Cruz, L.J. Ceramide and palmitic acid inhibit macrophage-mediated epithelial–mesenchymal transition in colorectal cancer. Mol. Cell. Biochem. 2020, 468, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Librán-Pérez, M.; Pereiro, P.; Figueras, A.; Novoa, B. Antiviral activity of palmitic acid via autophagic flux inhibition in zebrafish (Danio rerio). Fish Shellfish Immunol. 2019, 95, 595–605. [Google Scholar] [CrossRef]
- Comendador, J.E.B. A barrel of sponges: A Xestospongia testudinaria species complex in the Verde Island Passage, Philippines. Master’s Thesis, San Francisco State University, San Francisco, CA, USA, 2020. [Google Scholar]
- Ashok, P.; Ganguly, S.; Murugesan, S. Manzamine alkaloids: Isolation, cytotoxicity, antimalarial activity and SAR studies. Drug Discov. Today 2014, 19, 1781–1791. [Google Scholar] [CrossRef]
- Ashok, P.; Lathiya, H.; Murugesan, S. Manzamine alkaloids as antileishmanial agents: A review. Eur. J. Med. Chem. 2015, 97, 928–936. [Google Scholar] [CrossRef]
- Iwagawa, T.; Kaneko, M.; Okamura, H.; Nakatani, M.; van Soest, R.W.M.; Shiro, M. A new quinolizidine alkaloid from the Papua New Guinean sponge Xestospongia exigua. J. Nat. Prod. 2000, 63, 1310–1311. [Google Scholar] [CrossRef]
- Majali, I.; Qaralleh, H.N.; Idid, S.Z.; Saad, S.; Susanti, D.; Althunibat, O.Y. Potential Antimicrobial Activity of Marine Sponge Neopetrosia exigua. J. Basic Appl. Res. 2015, 1, 1–13. [Google Scholar] [CrossRef]
- Nakagawa, M.; Endo, M.; Tanaka, N.; Gen-Pei, L. Structures of xestospongin A, B, C and D, novel vasodilativecompounds from marine sponge, xestospongia exigua. Tetrahedron Lett. 1984, 25, 3227–3230. [Google Scholar] [CrossRef]
- Orabi, K.Y.; El Sayed, K.A.; Hamann, M.T.; Chuck Dunbar, D.; Al-Said, M.S.; Higa, T.; Kelly, M. Araguspongines K and L, new bioactive bis-1-oxaquinolizidine N-oxide alkaloids from red sea specimens of Xestospongia exigua. J. Nat. Prod. 2002, 65, 1782–1785. [Google Scholar] [CrossRef] [Green Version]
- Brastianos, H.C.; Vottero, E.; Patrick, B.O.; Van Soest, R.; Matainaho, T.; Mauk, A.G.; Andersen, R.J. Exiguamine A, an indoleamine-2,3-dioxygenase (IDO) inhibitor isolated from the marine sponge Neopetrosia exigua. J. Am. Chem. Soc. 2006, 128, 16046–16047. [Google Scholar] [CrossRef]
- Williams, D.E.; Lassota, P.; Andersen, R.J. Motuporamines A−C, Cytotoxic Alkaloids Isolated from the Marine Sponge Xestospongia exigua (Kirkpatrick). J. Org. Chem. 1998, 63, 4838–4841. [Google Scholar] [CrossRef]
- Pereira, R.B.; Pereira, D.M.; Jiménez, C.; Rodríguez, J.; Nieto, R.M.; Videira, R.A.; Silva, O.; Andrade, P.B.; Valentaõ, P. Anti-inflammatory effects of 5α,8α-epidioxycholest-6-en-3β-ol, a steroidal endoperoxide isolated from aplysia depilans, based on bioguided fractionation and NMR analysis. Mar. Drugs 2019, 17, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieber, M.A.; Hegel, J.K.E. Azelaic acid: Properties and mode of action. Skin Pharmacol. Physiol. 2013, 27, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, R.; Jebanesan, A.; Govindarajan, M.; Rajasekar, P. Larvicidal and repellent activity of tetradecanoic acid against Aedes aegypti (Linn.) and Culex quinquefasciatus (Say.) (Diptera:Culicidae). Asian Pac. J. Trop. Med. 2011, 4, 706–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Yuan, Y.; Lu, D.; Du, B.; Xiong, L.; Shi, J.; Yang, L.; Liu, W.; Yuan, X.; Zhang, G.; et al. Two natural products, trans-phytol and (22E)-ergosta-6,9,22-triene-3β,5α,8α-triol, inhibit the biosynthesis of estrogen in human ovarian granulosa cells by aromatase (CYP19). Toxicol. Appl. Pharmacol. 2014, 279, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E.; Lin, Z.; Imir, G.; Amin, S.; Demura, M.; Yilmaz, B.; Martin, R.; Utsunomiya, H.; Thung, S.; Gurates, B.; et al. Regulation of aromatase expression in estrogen-responsive breast and uterine disease: From bench to treatment. Pharmacol. Rev. 2005, 57, 359–383. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.; Islam, M.; Santos, P.; Ferreira, P.; Oliveira, G.; Alencar, M.; Paz, M.; Ferreira, É.; Feitosa, C.; Citó, A.; et al. Evaluation of Antioxidant Activity of Phytol Using Non- and Pre-Clinical Models. Curr. Pharm. Biotechnol. 2016, 17, 1278–1284. [Google Scholar] [CrossRef]
- Marotta, L.L.C.; Almendro, V.; Marusyk, A.; Shipitsin, M.; Schemme, J.; Walker, S.R.; Bloushtain-Qimron, N.; Kim, J.J.; Choudhury, S.A.; Maruyama, R.; et al. The JAK2/STAT3 signaling pathway is required for growth of CD44 +CD24- stem cell-like breast cancer cells in human tumors. J. Clin. Investig. 2011, 121, 2723–2735. [Google Scholar] [CrossRef]
- To, N.B.; Nguyen, Y.T.K.; Moon, J.Y.; Ediriweera, M.K.; Cho, S.K. Pentadecanoic acid, an odd-chain fatty acid, suppresses the stemness of MCF-7/SC human breast cancer stem-like cells through JAK2/STAT3 signaling. Nutrients 2020, 12, 1663. [Google Scholar] [CrossRef]
- Dilika, F.; Bremner, P.D.; Meyer, J.J.M. Antibacterial activity of linoleic and oleic acids isolated from Helichrysum pedunculatum: A plant used during circumcision rites. Fitoterapia 2000, 71, 450–452. [Google Scholar] [CrossRef]
- Vanitha, V.; Vijayakumar, S.; Nilavukkarasi, M.; Punitha, V.N.; Vidhya, E.; Praseetha, P.K. Heneicosane—A novel microbicidal bioactive alkane identified from Plumbago zeylanica L. Ind. Crops Prod. 2020, 154, 112748. [Google Scholar] [CrossRef]
- Kim, D.Y.; Won, K.J.; Il Hwang, D.; Kim, N.Y.; Kim, B.; Lee, H.M. 1-Iodohexadecane Alleviates 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis in Mice: Possible Involvements of the Skin Barrier and Mast Cell SNARE Proteins. Molecules 2022, 27, 1560. [Google Scholar] [CrossRef]
- Swantara, M.D.; Rita, W.S.; Suartha, N.; Agustina, K.K. Anticancer activities of toxic isolate of Xestospongia testudinaria sponge. Vet. World 2019, 12, 1434–1440. [Google Scholar] [CrossRef]
- Cita, Y.P.; Suhermanto, A.; Radjasa, O.K.; Sudharmono, P. Antibacterial activity of marine bacteria isolated from sponge Xestospongia testudinaria from Sorong, Papua. Asian Pac. J. Trop. Biomed. 2017, 7, 450–454. [Google Scholar] [CrossRef]
- El-Shitany, N.A.; Shaala, L.A.; Abbas, A.T.; Abdel-dayem, U.A.; Azhar, E.I.; Ali, S.S.; van Soest, R.W.M.; Youssef, D.T.A. Evaluation of the Anti-Inflammatory, Antioxidant and Immunomodulatory Effects of the Organic Extract of the Red Sea Marine Sponge Xestospongia testudinaria against Carrageenan Induced Rat Paw Inflammation. PLoS ONE 2015, 10, e0138917. [Google Scholar] [CrossRef] [Green Version]
- Swantara, I.M.D.; Rita, W.S. Toxicity of sponge extract Xestospongia testudinaria. Int. J. Eng. Sci. Invent. 2018, 7, 55–58. [Google Scholar]
- Zhou, C.; Yuan, K.; Tang, X.; Hu, N.; Peng, W. Molecular genetic evidence for polyandry in Ascaris suum. Parasitol. Res. 2011, 108, 703–708. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Metabolites | Class | MF | MW | Bioactivity | References |
|---|---|---|---|---|---|---|
| X. bergquistia | Xestobergsterol A (1) | Steroid | C27H44O5 | 448.6 | Anti-Inflammatory, Cytotoxic | [4,5,6,49,50] |
| Xestobergsterol B (2) | Steroid | C27H44O7 | 480.6 | Anti-Inflammatory, Cytotoxic | ||
| Xestobergsterol C (3) | Steroid | C27H44O6 | 464.6 | Cytotoxic | [49,50] | |
| X. muta | Manzamine C (4) | Alkaloid | C23H29N3 | 347.5 | Cytotoxic | [58,59] |
| X.exigua | Xestospongin A (5) | Alkaloid | C28H50N2O2 | 446.7 | Vasodilator | [62] |
| Xestospongin B (6) | Alkaloid | C29H52N2O3 | 476.7 | Vasodilator | ||
| Xestospongin C (7) | Alkaloid | C28H50N2O2 | 446.7 | Vasodilator | ||
| Xestospongin D (8) | Alkaloid | C28H50N2O3 | 462.7 | Vasodilator | ||
| Araguspongine K (9) | Alkaloid | C28H52N2O4 | Vasodilation | [62,63] | ||
| Araguspongine L (10) | Alkaloid | C28H52N2O5 | Vasodilation | |||
| Exiguamine A (12) | Alkaloid | C25H26N5O6 | 492.5 | Indoleamine-2,3-Dioxygenase Inhibitor | [64] | |
| Motuporamine A (13) | Alkaloid | C18H39N3 | 297.5 | Cytotoxicity | [65] | |
| Halenaquinone (22) | Quinone | C20H12O5 | 332.3 | Antibacterial | [52] | |
| Exiguaquinol (23) | Quinone | C22H21NO12S2 | 555.5 | Helicobacter Pylori Glutamate Racemase (Muri) Inhibitor | [51] | |
| Clionasterol (24) | Steroid | C29H50O | 414.7 | Inhibit The Human Complement System | [53] | |
| 5α,8α-Epidioxy-24αethylcholest-6-en-βb-ol (25) | Steroid | C29H48O3 | Anti-Inflammatory, Inhibit the Human Complement System | [53,66] | ||
| X. testudinaria | Nonanedioic Acid (26) | Fatty acid | C9H16O4 | 188 | Antibacterial And Anti-inflammatory | [67] |
| TetradecanoicAcid (27) | Fatty acid | C14H28O2 | 228 | Larvicidal | [68] | |
| Trans-Phytol (28) | Fatty acid | C20H40O | 296 | Aromatase Inhibitor, Antioxidant | [69,71] | |
| Pentadecanoic Acid (29) | Fatty acid | C15H30O2 | 242 | Anticancer | [73] | |
| Palmitic Acid (30) | Fatty acid | C16H32O2 | 256 | Antiviral, Anticancer | [54,55,56] | |
| 9,12-Octadecadienoic Acid (31) | Fatty acid | C19H34O2 | 294 | Antibacterial | [74] | |
| Heneicosane (32) | Alkane | C21H44 | 296 | Antimicrobial | [75] | |
| 1-Iodohexadecane (33) | Alkane | C16H33I | 352 | Anti-inflammatory | [76] | |
| X. ashmorica | Manzamine A (34) | Alkaloid | C36H44N4O | 548.8 | Cytotoxic | [42] |
| Manzamine E (35) | Alkaloid | C36H44N4O2 | 564.8 | Cytotoxic | [42] | |
| Manzamine F (36) | Alkaloid | C36H44N4O3 | 580.8 | Cytotoxic | [42] | |
| X. vansoesti | Salsolinol (37) | Alkaloid | C10H13NO2 | 179.2 | Cytotoxic | [39] |
| Norsalsolinol (38) | Alkaloid | C9H11NO2 | 165 | Cytotoxic | [39] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khodzori, F.A.; Mazlan, N.B.; Chong, W.S.; Ong, K.H.; Palaniveloo, K.; Shah, M.D. Metabolites and Bioactivity of the Marine Xestospongia Sponges (Porifera, Demospongiae, Haplosclerida) of Southeast Asian Waters. Biomolecules 2023, 13, 484. https://doi.org/10.3390/biom13030484
Khodzori FA, Mazlan NB, Chong WS, Ong KH, Palaniveloo K, Shah MD. Metabolites and Bioactivity of the Marine Xestospongia Sponges (Porifera, Demospongiae, Haplosclerida) of Southeast Asian Waters. Biomolecules. 2023; 13(3):484. https://doi.org/10.3390/biom13030484
Chicago/Turabian StyleKhodzori, Fikri Akmal, Nurzafirah Binti Mazlan, Wei Sheng Chong, Kuan Hung Ong, Kishneth Palaniveloo, and Muhammad Dawood Shah. 2023. "Metabolites and Bioactivity of the Marine Xestospongia Sponges (Porifera, Demospongiae, Haplosclerida) of Southeast Asian Waters" Biomolecules 13, no. 3: 484. https://doi.org/10.3390/biom13030484
APA StyleKhodzori, F. A., Mazlan, N. B., Chong, W. S., Ong, K. H., Palaniveloo, K., & Shah, M. D. (2023). Metabolites and Bioactivity of the Marine Xestospongia Sponges (Porifera, Demospongiae, Haplosclerida) of Southeast Asian Waters. Biomolecules, 13(3), 484. https://doi.org/10.3390/biom13030484