
Design and Assembly of a Biofactory for (2S)-Naringenin Production in Escherichia coli: Effects of Oxygen Transfer on Yield and Gene Expression

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pathway Selection and Sensitivity Analysis
2.2. Heterologous Pathway Construction and Assembly
2.3. Strains and Media
2.4. 50 mL Shake Flask (2S)-Naringenin Culture
2.5. 4-L Cultures
2.6. Analytical Procedures
2.7. Gene Expression Analysis
2.8. Statistical Analysis
3. Results
3.1. Prediction of the Shortest Pathway for Heterologous (2S)-Naringenin Production
3.2. Selection of Pathway Genes Sequence
3.3. Selection of Genes Sensitive to Oxygen Variation
3.4. (2S)-Naringenin Production at a Laboratory Scale
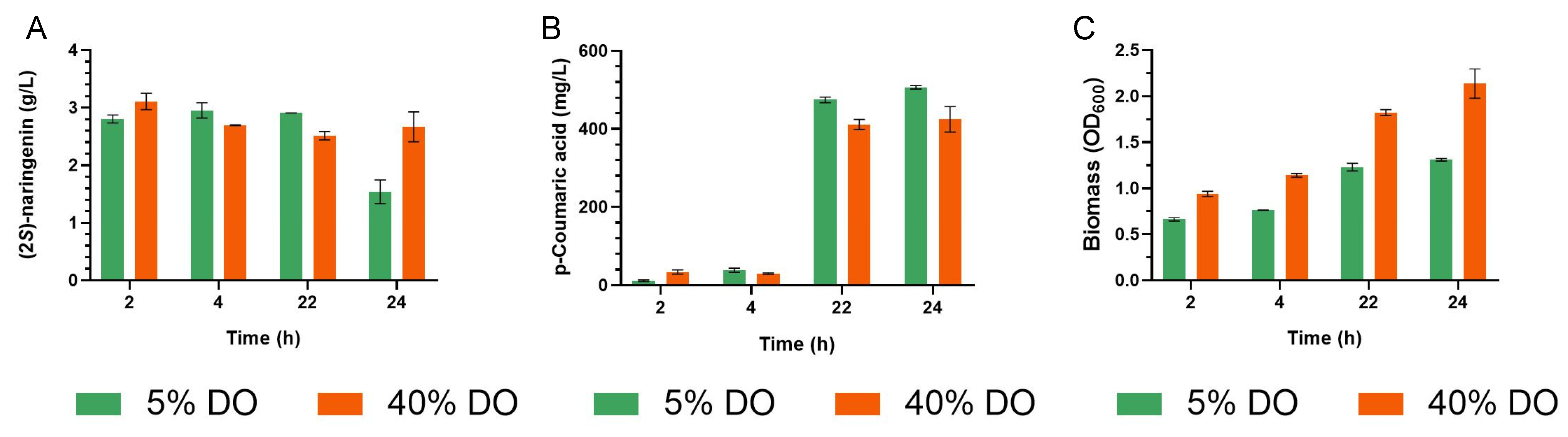
3.5. (2S)-Naringenin Production at Pilot Scale
3.6. Gene Expression Analysis at Different Oxygen Transfer Conditions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brglez Mojzer, E.; Knez Hrnčič, M.; Škerget, M.; Knez, Ž.; Bren, U. Polyphenols: Extraction Methods, Antioxidative Action, Bioavailability and Anticarcinogenic Effects. Molecules 2016, 21, 901. [Google Scholar] [CrossRef]
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P.E. Polyphenols and Human Health: Prevention of Disease and Mechanisms of Action. Nutrients 2010, 2, 1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, Biological Functions, and Biotechnological Applications. Front. Plant. Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Zhou, T.; Du, G.; Zhou, J.; Chen, J. Modular Optimization of Heterologous Pathways for de Novo Synthesis of (2S)-Naringenin in Escherichia coli. PLoS ONE 2014, 9, e101492. [Google Scholar] [CrossRef] [PubMed]
- Carocho, M.; Ferreira, I.C.F.R. A Review on Antioxidants, Prooxidants and Related Controversy: Natural and Synthetic Compounds, Screening and Analysis Methodologies and Future Perspectives. Food Chem. Toxicol. 2013, 51, 15–25. [Google Scholar] [CrossRef]
- Karim, N.; Jia, Z.; Zheng, X.; Cui, S.; Chen, W. A Recent Review of Citrus Flavanone Naringenin on Metabolic Diseases and Its Potential Sources for High Yield-Production. Trends Food Sci. Technol. 2018, 79, 35–54. [Google Scholar] [CrossRef]
- Alberca, R.W.; Teixeira, F.M.E.; Beserra, D.R.; de Oliveira, E.A.; Andrade, M.M.D.S.; Pietrobon, A.J.; Sato, M.N. Perspective: The Potential Effects of Naringenin in COVID-19. Front. Immunol. 2020, 11, 919. [Google Scholar] [CrossRef]
- Clementi, N.; Scagnolari, C.; D’Amore, A.; Palombi, F.; Criscuolo, E.; Frasca, F.; Pierangeli, A.; Mancini, N.; Antonelli, G.; Clementi, M.; et al. Naringenin Is a Powerful Inhibitor of SARS-CoV-2 Infection in Vitro. Pharm. Res. 2021, 163, 105255. [Google Scholar] [CrossRef]
- Salehi, B.; Fokou, P.V.T.; Sharifi-Rad, M.; Zucca, P.; Pezzani, R.; Martins, N.; Sharifi-Rad, J. The Therapeutic Potential of Naringenin: A Review of Clinical Trials. Pharmaceuticals 2019, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Tutunchi, H.; Naeini, F.; Ostadrahimi, A.; Hosseinzadeh-Attar, M.J. Naringenin, a Flavanone with Antiviral and Anti-Inflammatory Effects: A Promising Treatment Strategy against COVID-19. Phytother. Res. 2020, 34, 3137–3147. [Google Scholar] [CrossRef]
- Sotelo, L.; Alvis, A.; Arrázola, G. Evaluation of Epicatechin, Theobromine and Caffeine in Cacao Husks (Theobroma Cacao, L.), Determination of the Antioxidant Capacity. Scielo 2015, 9, 124–134. [Google Scholar] [CrossRef]
- Koopman, F.; Beekwilder, J.; Crimi, B.; van Houwelingen, A.; Hall, R.D.; Bosch, D.; van Maris, A.J.A.; Pronk, J.T.; Daran, J. De Novo Production of the Flavonoid Naringenin in Engineered Saccharomyces cerevisiae. Microb. Cell Factories 2012, 11, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, H.; Song, M.C.; Lee, J.Y.; Yoon, Y.J. Microbial Production of O-Methylated Flavanones from Methylated Phenylpropanoic Acids in Engineered Escherichia coli. J. Ind. Microbiol Biotechnol. 2019, 46, 1707–1713. [Google Scholar] [CrossRef] [PubMed]
- Miyahisa, I.; Kaneko, M.; Funa, N.; Kawasaki, H.; Kojima, H.; Ohnishi, Y.; Horinouchi, S. Efficient Production of (2S)-Flavanones by Escherichia coli Containing an Artificial Biosynthetic Gene Cluster. Appl. Microbiol. Biotechnol. 2005, 68, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Ranganathan, S.; Fowler, Z.L.; Maranas, C.D.; Koffas, M.A.G. Genome-Scale Metabolic Network Modeling Results in Minimal Interventions That Cooperatively Force Carbon Flux towards Malonyl-CoA. Metab. Eng. 2011, 13, 578–587. [Google Scholar] [CrossRef]
- Zhou, S.; Hao, T.; Zhou, J. Fermentation and Metabolic Pathway Optimization to de Novo Synthesize (2S)-Naringenin in Escherichia coli. J. Microbiol. Biotechnol. 2020, 30, 1574–1582. [Google Scholar] [CrossRef]
- Gao, S.; Lyu, Y.; Zeng, W.; Du, G.; Zhou, J.; Chen, J. Efficient Biosynthesis of (2S)-Naringenin from p-Coumaric Acid in Saccharomyces cerevisiae. J. Agric. Food. Chem. 2020, 68, 1015–1021. [Google Scholar] [CrossRef]
- Fowler, Z.L.; Gikandi, W.W.; Koffas, M.A.G. Increased Malonyl Coenzyme A Biosynthesis by Tuning the Escherichia coli Metabolic Network and Its Application to Flavanone Production. Appl. Environ. Microbiol. 2009, 75, 5831–5839. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Yuan, S.-F.; Nair, P.H.; Alper, H.S.; Deng, Y.; Zhou, J. Development of a Growth Coupled Dynamic Regulation Network Balancing Malonyl-CoA Node to Enhance (2S)-Naringenin Synthesis in E. coli. bioRxiv 2020, 67, 41–52. [Google Scholar] [CrossRef]
- Santos, C.N.S.; Koffas, M.; Stephanopoulos, G. Optimization of a Heterologous Pathway for the Production of Flavonoids from Glucose. Metab. Eng. 2011, 13, 392–400. [Google Scholar] [CrossRef]
- Chowdhury, A.; Maranas, C.D. Designing Overall Stoichiometric Conversions and Intervening Metabolic Reactions. Sci. Rep. 2015, 5, 16009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Vansiri, A.; Bhan, N.; Koffas, M.A. EPathBrick: A Synthetic Biology Platform for Engineering Metabolic Pathways in E. coli EPathBrick: A Synthetic Biology Platform for Engineering Metabolic Pathways. ACS Synth. Biol. 2012, 1, 256–266. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Tafur Rangel, A.E.; Camelo Valera, L.C.; Gómez Ramírez, J.M.; González Barrios, A.F. Effects of Metabolic Engineering on Downstream Processing Operational Cost and Energy Consumption: The Case of Escherichia coli’s Glycerol Conversion to Succinic Acid. J. Chem. Technol. Biotechnol. 2018, 93, 2011–2020. [Google Scholar] [CrossRef]
- Edwards, J.S.; Palsson, B.O. Robustness Analysis of the Escherichia coli Metabolic Network. Biotechnol. Prog. 2000, 16, 927–939. [Google Scholar] [CrossRef]
- López-Agudelo, V.A.; Baena, A.; Ramirez-Malule, H.; Ochoa, S.; Barrera, L.F.; Ríos-Estepa, R. Metabolic Adaptation of Two in Silico Mutants of Mycobacterium Tuberculosis during Infection. BMC Syst. Biol. 2017, 11, 107. [Google Scholar] [CrossRef] [Green Version]
- Winck, F.V.; Páez Melo, D.O.; Riaño-Pachón, D.M.; Martins, M.C.M.; Caldana, C.; González Barrios, A.F. Analysis of Sensitive CO2 Pathways and Genes Related to Carbon Uptake and Accumulation in Chlamydomonas Reinhardtii through Genomic Scale Modeling and Experimental Validation. Front. Plant. Sci. 2016, 7, 43. [Google Scholar] [CrossRef] [Green Version]
- Schomburg, I.; Chang, A.; Schomburg, D. BRENDA, Enzyme Data and Metabolic Information. Nucleic Acids Res. 2002, 30, 47–49. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hsieh, C.Y.; Huang, Y.H.; Yeh, H.H.; Hong, P.Y.; Hsiao, C.J.; Hsieh, L.S. Phenylalanine, Tyrosine, and Dopa Are Bona Fide Substrates for Bambusa oldhamii Bopal4. Catalysts 2021, 11, 1263. [Google Scholar] [CrossRef]
- Lussier, F.X.; Colatriano, D.; Wiltshire, Z.; Page, J.E.; Martin, V.J.J. Engineering Microbes for Plant Polyketide Biosynthesis. Comput. Struct. Biotechnol. J. 2012, 3, e201210020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, L.S.; Ma, G.J.; Yang, C.C.; Lee, P.D. Cloning, Expression, Site-Directed Mutagenesis and Immunolocalization of Phenylalanine Ammonia-Lyase in Bambusa oldhamii. Phytochemistry 2010, 71, 1999–2009. [Google Scholar] [CrossRef] [PubMed]
- Alberstein, M.; Eisenstein, M.; Abeliovich, H. Removing Allosteric Feedback Inhibition of Tomato 4-Coumarate.:CoA Ligase by Directed Evolution. Plant J. 2012, 69, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Falkenstein-Paul, H.; Schmidt, W.; Beerhues, L. Benzophenone Synthase and Chalcone Synthase from Hypericum androsaemum Cell Cultures: CDNA Cloning, Functional Expression, and Site-Directed Mutagenesis of Two Polyketide Synthases. Plant J. 2003, 34, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Joung, J.Y.; Mangai Kasthuri, G.; Park, J.Y.; Kang, W.J.; Kim, H.S.; Yoon, B.S.; Joung, H.; Jeon, J.H. An Overexpression of Chalcone Reductase of Pueraria montana var. lobata Alters Biosynthesis of Anthocyanin and 5′-Deoxyflavonoids in Transgenic Tobacco. Biochem. Biophys. Res. Commun. 2003, 303, 326–331. [Google Scholar] [CrossRef]
- Kyndt, J.A.; Meyer, T.E.; Cusanovich, M.A.; van Beeumen, J.J. Characterization of a Bacterial Tyrosine Ammonia Lyase, a Biosynthetic Enzyme for the Photoactive Yellow Protein. FEBS Lett. 2002, 512, 240–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jez, J.M.; Noel, J.P. Reaction Mechanism of Chalcone Isomerase. J. Biol. Chem. 2002, 277, 1361–1369. [Google Scholar] [CrossRef] [Green Version]
- Skern, R.; Frost, P.; Nilsen, F. Relative Transcript Quantification by Quantitative PCR: Roughly Right or Precisely Wrong? BMC Mol. Biol. 2005, 6, 1–3. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Biosynthesis Genes | Titer (mg/L) | Reference |
|---|---|---|---|
| E. coli BLR (DE3) | pal, 4cl, chs, chi and acc | 57 | [14] |
| E. coli BL21 StarTM | 4cl, chs, chi, acc, pgk, pdh | 474 | [15] |
| E. coli rpoA14R | tal, 4cl, chi, chs | 84 | [20] |
| S. cerevisiae CEN.PK2-1c | pal, c4h, cpr, 4cl, chs, chi, tal | 109 | [12] |
| E. coli BL21 (DE3) | 4cl, chs, chi | 588 | [19] |
| S. cerevisiae CEN.PK2-1D | tal, 4cl, chs, chi, aroL, ARO7fb, ARO4fbr | 648.93 | [17] |
| Strain | Genotype | |
|---|---|---|
| Escherichia coli BL21(DE3) | F− ompT hsdSB (rB−, mB−) gal dcm (DE3) | |
| Escherichia coli DH5α | F–φ80lacZΔM15 Δ(lacZYA-argF)U169 recA1 endA1 hsdR17(rK−, mK+) phoA supE44 λ–thi-1 gyrA96 relA1 | |
| Plasmid | Description | Source of reference |
| pETM6 | T7 promoter, ampicillin resistance | Addgene |
| pRSM3 | T7 promoter, kanamycin resistance | Addgene |
| pEPACH | pETM6 carrying pal and chi genes | This study |
| pR4CCH | pRSM3 carrying 4cl and chs genes | This study |
| Organism | Km (mM) | Mutation Presence | References |
|---|---|---|---|
| pal gene | |||
| B. oldhamii/BoPAL1 | 0.230 | Yes | [32] |
| B. oldhamii/BoPAL2 | 0.333 | Yes | [32] |
| B. oldhamii/BoPAL4 | 0.097 | Yes | [32] |
| R. toruloides | 0.180 | No | [36] |
| 4cl gene | |||
| S. lycopersicum Q274H | 0.179 | Yes | [33] |
| S. lycopersicum Q274H-F269L | 0.036 | Yes | [33] |
| chs gene | |||
| H. androsaemum | 0.023 | Yes | [34] |
| M. sativa | 0.023 | Yes | [37] |
| chi gene | |||
| H. androsaemum | 0.023 | Yes | [35] |
| Descriptive Name | Function |
|---|---|
| (fadE) Acyl-coenzyme A dehydrogenase | Fatty acids |
| (fadB) Fatty acid oxidation complex, alpha subunit | Fatty acids |
| (tesB) Acyl-CoA thioesterase 2 | Fatty acids |
| (fadI) 3-ketoacyl-CoA thiolase | Fatty acids |
| (pyrD) Dihydroorotate dehydrogenase | Pyrimide metabolism |
| (frdA) Fumarate reductase flavoprotein subunit | Electron transport gene |
| (nuoA) NADH-quinone oxidoreductase subunit A | Electron transport gene |
| (cyoD) Cytochrome bo(3) ubiquinol oxidase subunit 4 | Electron transport gene |
| (mraY) phospho-N-acetylmuramoyl-pentapeptide-transferase | Division cell |
| (murG) UDP-N-acetylglucosamine-acetylglucosamine transferase | Division cell |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parra Daza, L.E.; Suarez Medina, L.; Tafur Rangel, A.E.; Fernández-Niño, M.; Mejía-Manzano, L.A.; González-Valdez, J.; Reyes, L.H.; González Barrios, A.F. Design and Assembly of a Biofactory for (2S)-Naringenin Production in Escherichia coli: Effects of Oxygen Transfer on Yield and Gene Expression. Biomolecules 2023, 13, 565. https://doi.org/10.3390/biom13030565
Parra Daza LE, Suarez Medina L, Tafur Rangel AE, Fernández-Niño M, Mejía-Manzano LA, González-Valdez J, Reyes LH, González Barrios AF. Design and Assembly of a Biofactory for (2S)-Naringenin Production in Escherichia coli: Effects of Oxygen Transfer on Yield and Gene Expression. Biomolecules. 2023; 13(3):565. https://doi.org/10.3390/biom13030565
Chicago/Turabian StyleParra Daza, Laura E., Lina Suarez Medina, Albert E. Tafur Rangel, Miguel Fernández-Niño, Luis Alberto Mejía-Manzano, José González-Valdez, Luis H. Reyes, and Andrés Fernando González Barrios. 2023. "Design and Assembly of a Biofactory for (2S)-Naringenin Production in Escherichia coli: Effects of Oxygen Transfer on Yield and Gene Expression" Biomolecules 13, no. 3: 565. https://doi.org/10.3390/biom13030565
APA StyleParra Daza, L. E., Suarez Medina, L., Tafur Rangel, A. E., Fernández-Niño, M., Mejía-Manzano, L. A., González-Valdez, J., Reyes, L. H., & González Barrios, A. F. (2023). Design and Assembly of a Biofactory for (2S)-Naringenin Production in Escherichia coli: Effects of Oxygen Transfer on Yield and Gene Expression. Biomolecules, 13(3), 565. https://doi.org/10.3390/biom13030565