
Aquaporin-9 in the Brain Inflammatory Response: Evidence from Mice Injected with the Parkinsonogenic Toxin MPP+

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Stereotaxic Surgery and Microinjection of MPP+
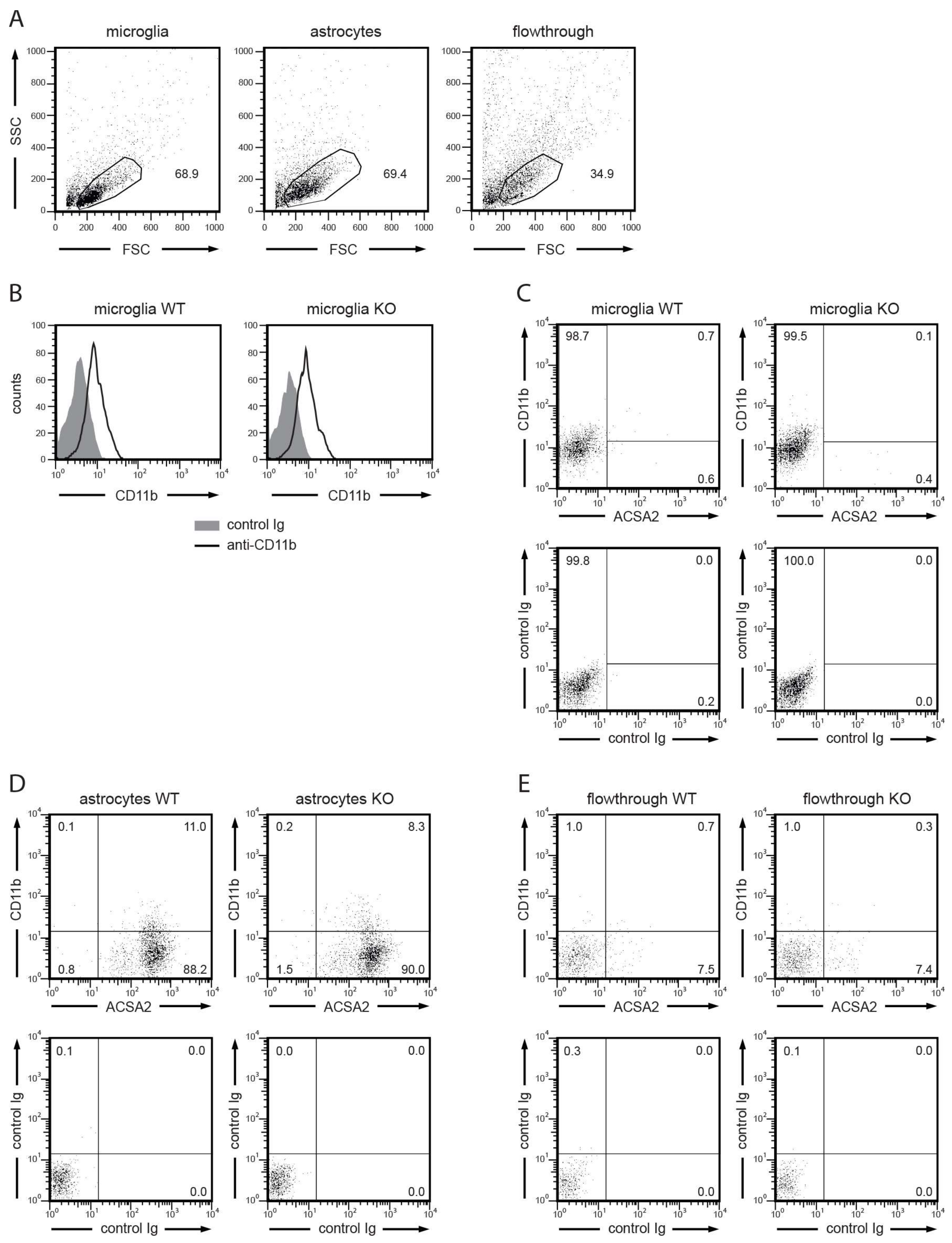
2.3. Microbead-Based Cell Separation of Microglia and Astrocytes
- a.
- Cell separation
- b.
- Debris removal
- c.
- Magnetic labeling of CD11b+ cells
- d.
- Magnetic separation of CD11b+ cells
- e.
- Magnetic labeling of ACSA-2+ cells
- f.
- Magnetic separation of ACSA-2+ cells
- g.
- Flow cytometric analysis
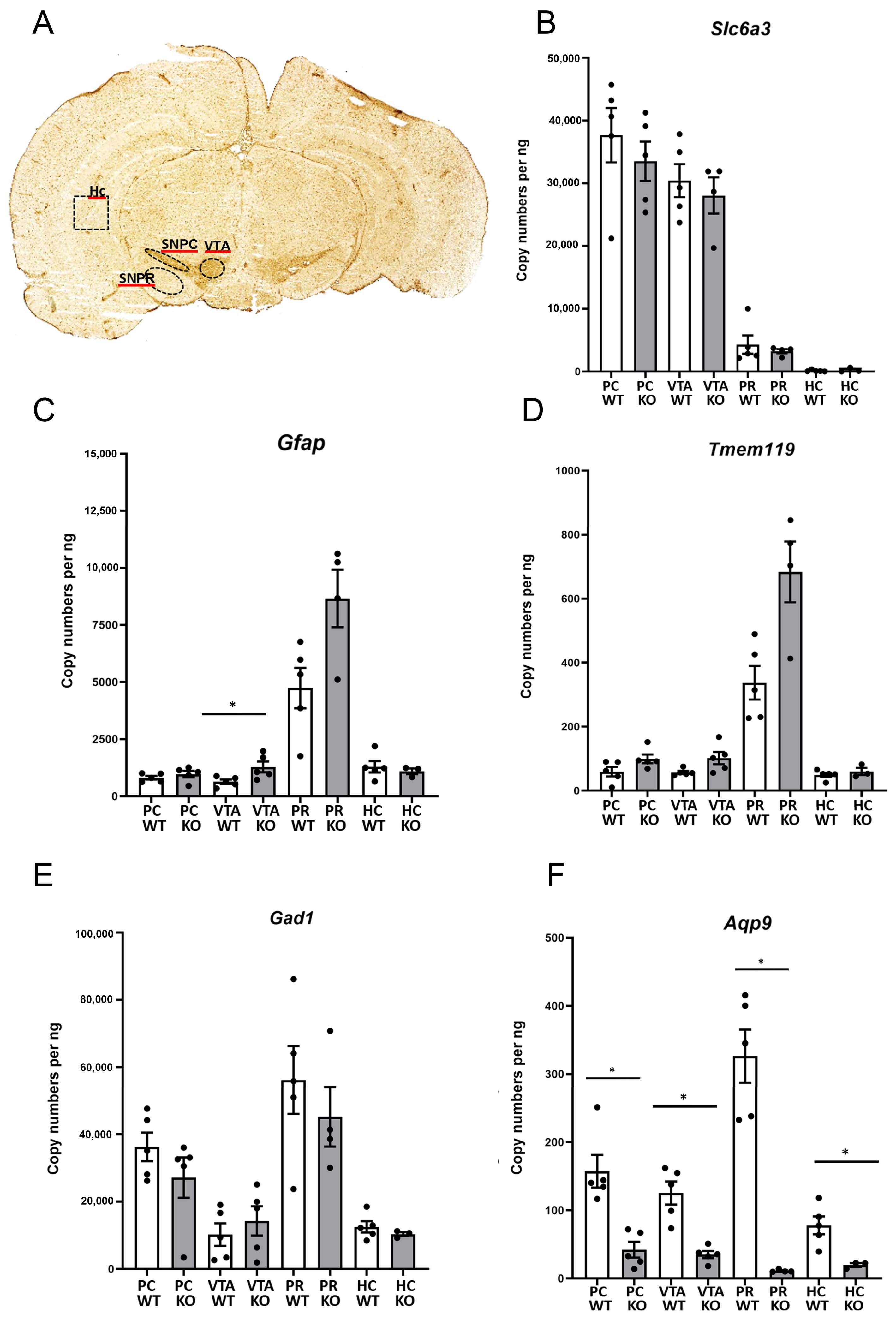
2.4. Laser Capture Microdissection (LCM)
- a.
- Preparation of LCM samples
- b.
- Tyrosine Hydroxylase (TH) immunostaining
- c.
- Laser capture microdissection (LCM)
2.5. RNA Isolation, cDNA Synthesis, and Quantitative Real Time qPCR
3. Results
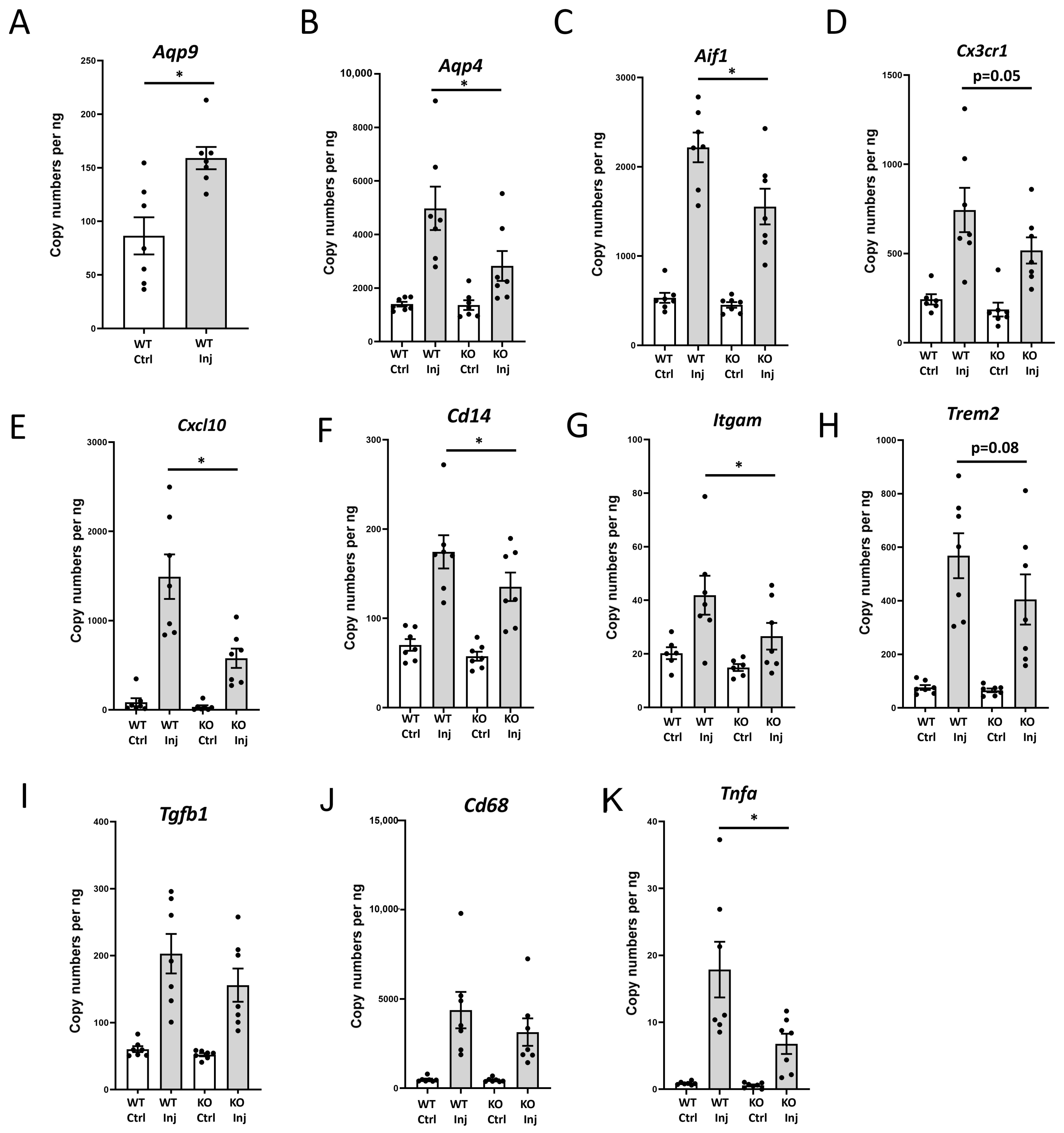
3.1. Genetic Deletion of Aqp9 Attenuates the Increase in the Transcript Levels of Proinflammatory Genes Following Unilateral Intrastriatal MPP+ Injections
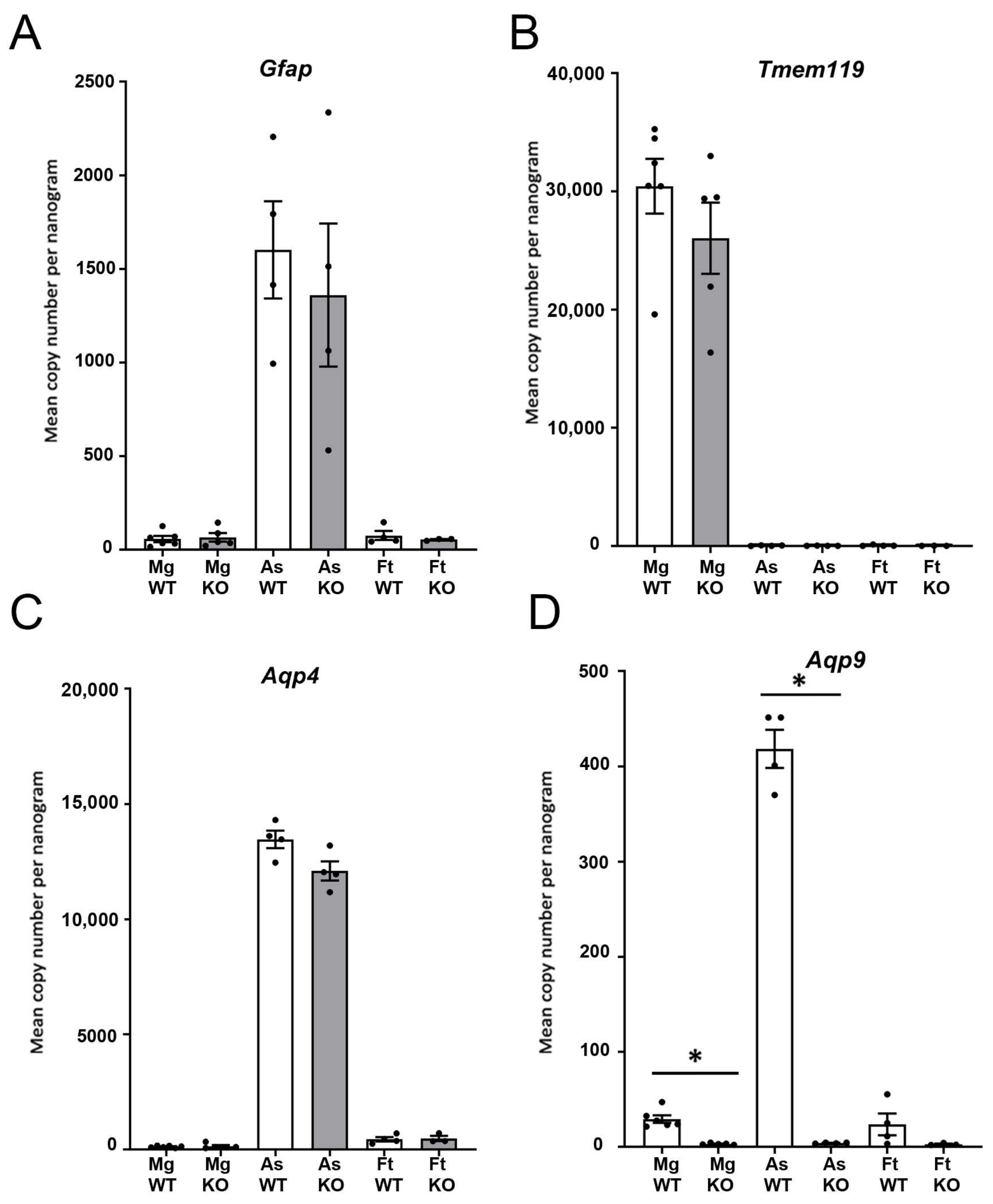
3.2. Isolation of Microglia and Astrocytes from the Brains of WT and Aqp9−/− Mice
3.3. Quantitative Real-Time PCR of Isolated Cells Shows Expression of Aqp9 in Microglia
3.4. The mRNA Levels of Aqp9 in Midbrain Subregions
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsukaguchi, H.; Shayakul, C.; Berger, U.V.; Mackenzie, B.; Devidas, S.; Guggino, W.B.; van Hoek, A.N.; Hediger, M.A. Molecular characterization of a broad selectivity neutral solute channel. J. Biol. Chem. 1998, 273, 24737–24743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elkjaer, M.; Vajda, Z.; Nejsum, L.N.; Kwon, T.; Jensen, U.B.; Amiry-Moghaddam, M.; Frokiaer, J.; Nielsen, S. Immunolocalization of AQP9 in liver, epididymis, testis, spleen, and brain. Biochem. Biophys. Res. Commun. 2000, 276, 1118–1128. [Google Scholar] [CrossRef] [PubMed]
- Badaut, J.; Petit, J.M.; Brunet, J.F.; Magistretti, P.J.; Charriaut-Marlangue, C.; Regli, L. Distribution of Aquaporin 9 in the adult rat brain: Preferential expression in catecholaminergic neurons and in glial cells. Neuroscience 2004, 128, 27–38. [Google Scholar] [CrossRef]
- Amiry-Moghaddam, M.; Lindland, H.; Zelenin, S.; Roberg, B.A.; Gundersen, B.B.; Petersen, P.; Rinvik, E.; Torgner, I.A.; Ottersen, O.P. Brain mitochondria contain aquaporin water channels: Evidence for the expression of a short AQP9 isoform in the inner mitochondrial membrane. FASEB J. 2005, 19, 1459–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mylonakou, M.N.; Petersen, P.H.; Rinvik, E.; Rojek, A.; Valdimarsdottir, E.; Zelenin, S.; Zeuthen, T.; Nielsen, S.; Ottersen, O.P.; Amiry-Moghaddam, M. Analysis of mice with targeted deletion of AQP9 gene provides conclusive evidence for expression of AQP9 in neurons. J. Neurosci. Res. 2009, 87, 1310–1322. [Google Scholar] [CrossRef]
- da Silva, I.V.; Garra, S.; Calamita, G.; Soveral, G. The Multifaceted Role of Aquaporin-9 in Health and Its Potential as a Clinical Biomarker. Biomolecules 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Tesse, A.; Gena, P.; Rutzler, M.; Calamita, G. Ablation of Aquaporin-9 Ameliorates the Systemic Inflammatory Response of LPS-Induced Endotoxic Shock in Mouse. Cells 2021, 10, 435. [Google Scholar] [CrossRef]
- Mohammad, S.; O’Riordan, C.E.; Verra, C.; Aimaretti, E.; Alves, G.F.; Dreisch, K.; Evenas, J.; Gena, P.; Tesse, A.; Rutzler, M.; et al. RG100204, A Novel Aquaporin-9 Inhibitor, Reduces Septic Cardiomyopathy and Multiple Organ Failure in Murine Sepsis. Front. Immunol. 2022, 13, 900906. [Google Scholar] [CrossRef]
- Prydz, A.; Stahl, K.; Zahl, S.; Skauli, N.; Skare, O.; Ottersen, O.P.; Amiry-Moghaddam, M. Pro-Inflammatory Role of AQP4 in Mice Subjected to Intrastriatal Injections of the Parkinsonogenic Toxin MPP. Cells 2020, 9, 2418. [Google Scholar] [CrossRef]
- Stahl, K.; Rahmani, S.; Prydz, A.; Skauli, N.; MacAulay, N.; Mylonakou, M.N.; Torp, R.; Skare, O.; Berg, T.; Leergaard, T.B.; et al. Targeted deletion of the aquaglyceroporin AQP9 is protective in a mouse model of Parkinson’s disease. PLoS ONE 2018, 13, e0194896. [Google Scholar] [CrossRef] [Green Version]
- Ginhoux, F.; Lim, S.; Hoeffel, G.; Low, D.; Huber, T. Origin and differentiation of microglia. Front. Cell. Neurosci. 2013, 7, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, R.; Mei, A.; Mathew, V.; Kashpur, O.; Wallingford, M.C. Interaction of extraembryonic microglia and neonatal brain development. Exp. Neurol. 2022, 351, 113986. [Google Scholar] [CrossRef] [PubMed]
- Rojek, A.M.; Skowronski, M.T.; Fuchtbauer, E.M.; Fuchtbauer, A.C.; Fenton, R.A.; Agre, P.; Frokiaer, J.; Nielsen, S. Defective glycerol metabolism in aquaporin 9 (AQP9) knockout mice. Proc. Natl. Acad. Sci. USA 2007, 104, 3609–3614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paxinos, G.; Franklin, K.B. The Mouse Brain in Sterotaxic Coordinates, 3rd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Kummari, E.; Guo-Ross, S.X.; Eells, J.B. Laser capture microdissection—A demonstration of the isolation of individual dopamine neurons and the entire ventral tegmental area. J. Vis. Exp. 2015, 96, e52336. [Google Scholar] [CrossRef] [Green Version]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.B.; Skauli, N.; Jovanovic, N.; Katoozi, S.; Frigeri, A.; Froehner, S.C.; Adams, M.E.; Ottersen, O.P.; Amiry-Moghaddam, M. Orchestrating aquaporin-4 and connexin-43 expression in brain: Differential roles of alpha1- and beta1-syntrophin. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183616. [Google Scholar] [CrossRef]
- Satoh, J.; Kino, Y.; Asahina, N.; Takitani, M.; Miyoshi, J.; Ishida, T.; Saito, Y. TMEM119 marks a subset of microglia in the human brain. Neuropathology 2016, 36, 39–49. [Google Scholar] [CrossRef]
- Badaut, J.; Brunet, J.F.; Regli, L. Aquaporins in the brain: From aqueduct to “multi-duct”. Metab. Brain Dis. 2007, 22, 251–263. [Google Scholar] [CrossRef]
- Papadopoulos, M.C.; Verkman, A.S. Aquaporin water channels in the nervous system. Nat. Rev. Neurosci. 2013, 14, 265–277. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, M.K.; Mestre, H.; Nedergaard, M. Fluid transport in the brain. Physiol. Rev. 2022, 102, 1025–1151. [Google Scholar] [CrossRef]
- Carbrey, J.M.; Gorelick-Feldman, D.A.; Kozono, D.; Praetorius, J.; Nielsen, S.; Agre, P. Aquaglyceroporin AQP9: Solute permeation and metabolic control of expression in liver. Proc. Natl. Acad. Sci. USA 2003, 100, 2945–2950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibashi, K.; Kuwahara, M.; Gu, Y.; Tanaka, Y.; Marumo, F.; Sasaki, S. Cloning and functional expression of a new aquaporin (AQP9) abundantly expressed in the peripheral leukocytes permeable to water and urea, but not to glycerol. Biochem. Biophys. Res. Commun. 1998, 244, 268–274. [Google Scholar] [CrossRef]
- Liu, Y.; Promeneur, D.; Rojek, A.; Kumar, N.; Frøkiaer, J.; Nielsen, S.; King, L.S.; Agre, P.; Carbrey, J.M. Aquaporin 9 is the major pathway for glycerol uptake by mouse erythrocytes, with implications for malarial virulence. Proc. Natl. Acad. Sci. USA 2007, 104, 12560–12564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, Y.; Kan, B.; Yang, B. Aquaporins in Immune System. Adv. Exp. Med. Biol. 2023, 1398, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Dheen, S.T.; Jun, Y.; Yan, Z.; Tay, S.S.; Ling, E.A. Retinoic acid inhibits expression of TNF-alpha and iNOS in activated rat microglia. Glia 2005, 50, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Haj-Yasein, N.N.; Vindedal, G.F.; Eilert-Olsen, M.; Gundersen, G.A.; Skare, O.; Laake, P.; Klungland, A.; Thoren, A.E.; Burkhardt, J.M.; Ottersen, O.P.; et al. Glial-conditional deletion of aquaporin-4 (Aqp4) reduces blood-brain water uptake and confers barrier function on perivascular astrocyte endfeet. Proc. Natl. Acad. Sci. USA 2011, 108, 17815–17820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Santis, S.; Serino, G.; Fiorentino, M.R.; Galleggiante, V.; Gena, P.; Verna, G.; Liso, M.; Massaro, M.; Lan, J.; Troisi, J.; et al. Aquaporin 9 Contributes to the Maturation Process and Inflammatory Cytokine Secretion of Murine Dendritic Cells. Front. Immunol. 2018, 9, 2355. [Google Scholar] [CrossRef] [Green Version]
- Moniaga, C.S.; Watanabe, S.; Honda, T.; Nielsen, S.; Hara-Chikuma, M. Aquaporin-9-expressing neutrophils are required for the establishment of contact hypersensitivity. Sci. Rep. 2015, 5, 15319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araújo, B.; Caridade-Silva, R.; Soares-Guedes, C.; Martins-Macedo, J.; Gomes, E.D.; Monteiro, S.; Teixeira, F.G. Neuroinflammation and Parkinson’s Disease-From Neurodegeneration to Therapeutic Opportunities. Cells 2022, 11, 2908. [Google Scholar] [CrossRef]
- McGeer, P.L.; McGeer, E.G. Inflammation and neurodegeneration in Parkinson’s disease. Parkinsonism Relat. Disord. 2004, 10 (Suppl. S1), S3–S7. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MPP+ Tissues | Separated Brain Cells | LCM | |
|---|---|---|---|
| RNA isolation kit | RNeasy® Mini Kit (QIAGEN, Hilden, Germany) | RNeasy® Plus Micro Kit (QIAGEN) | RNeasy® Plus Micro Kit (QIAGEN) |
| cDNA kit | 200ng RNA input GoScript Reverse Transcription System (Promega, Madison, WI, USA) | 100ng RNA input GoScript Reverse Transcription System (Promega) | 5 μL RNA input Sensiscript® Reverse Transcription Kit (QIAGEN) |
| Total cDNA template in qPCR | 1 ng | 2.5 ng | 5 μL |
| Evaluated endogenous reference genes | Ubc, Actb, Pgk1, H2afz | Ubc, H2afz, Hsp8a | Ubc, H2afz, Hsp8a |
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| Aqp9 | TTGCAACGGCAGTTGTGATG | CAAAAGACACCGCTGGGTTG |
| Aqp4 | TTTGGACCCGCAGTTATCAT | GTTGTCCTCCACCTCCATGT |
| Aif1 | CTGCCAGCCTAAGACAACCA | GGAATTGCTTGTTGATCCCCT |
| Cx3cr1 | TGCTCAGGACCTCACCATGTC | CTCAAGGCCAGGTTCAGGAG |
| Cxcl10 | ATGACGGGCCAGTGAGAATG | TCGTGGCAATGATCTCAACAC |
| Cd14 | CAGAGAACACCACCGCTGTA | CACGCTCCATGGTCGGTAGA |
| Itgam | TGCGCGAAGGAGATATCCAG | GCCTGCGTGTGTTGTTCTTT |
| Trem2 | CTGGAGGACCCTCTAGATGAC | CCACAGGATGAAACCTGCCT |
| Tgfb1 | AATTCCTGGCGTTACCTTGG | AGTGAGCGCTGAATCGAAAG |
| Cd68 | AGGACCGCTTATAGCCCAAG | TCATCGTGAAGGATGGCAGG |
| Tnfa | GGATGAGAAGTTCCCAAATGGC | ACTTGGTGGTTTGCTACGAC |
| Gad1 | CAGTCACCTGGAACCCTCAC | CACGGTGCCCTTTGCTTTC |
| Gfap | GCACTCAATACGAGGCAGTG | GCTCTAGGGACTCGTTCGTG |
| Tmem119 | CACCCAGAGCTGGTTCCATA | GTGACACAGAGTAGGCCACC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zahl, S.; Skauli, N.; Stahl, K.; Prydz, A.; Frey, M.M.; Dissen, E.; Ottersen, O.P.; Amiry-Moghaddam, M. Aquaporin-9 in the Brain Inflammatory Response: Evidence from Mice Injected with the Parkinsonogenic Toxin MPP+. Biomolecules 2023, 13, 588. https://doi.org/10.3390/biom13040588
Zahl S, Skauli N, Stahl K, Prydz A, Frey MM, Dissen E, Ottersen OP, Amiry-Moghaddam M. Aquaporin-9 in the Brain Inflammatory Response: Evidence from Mice Injected with the Parkinsonogenic Toxin MPP+. Biomolecules. 2023; 13(4):588. https://doi.org/10.3390/biom13040588
Chicago/Turabian StyleZahl, Soulmaz, Nadia Skauli, Katja Stahl, Agnete Prydz, Mina Martine Frey, Erik Dissen, Ole Petter Ottersen, and Mahmood Amiry-Moghaddam. 2023. "Aquaporin-9 in the Brain Inflammatory Response: Evidence from Mice Injected with the Parkinsonogenic Toxin MPP+" Biomolecules 13, no. 4: 588. https://doi.org/10.3390/biom13040588
APA StyleZahl, S., Skauli, N., Stahl, K., Prydz, A., Frey, M. M., Dissen, E., Ottersen, O. P., & Amiry-Moghaddam, M. (2023). Aquaporin-9 in the Brain Inflammatory Response: Evidence from Mice Injected with the Parkinsonogenic Toxin MPP+. Biomolecules, 13(4), 588. https://doi.org/10.3390/biom13040588