
Opposing Spatially Segregated Function of Endogenous GDNF-RET Signaling in Cocaine Addiction


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Genotyping
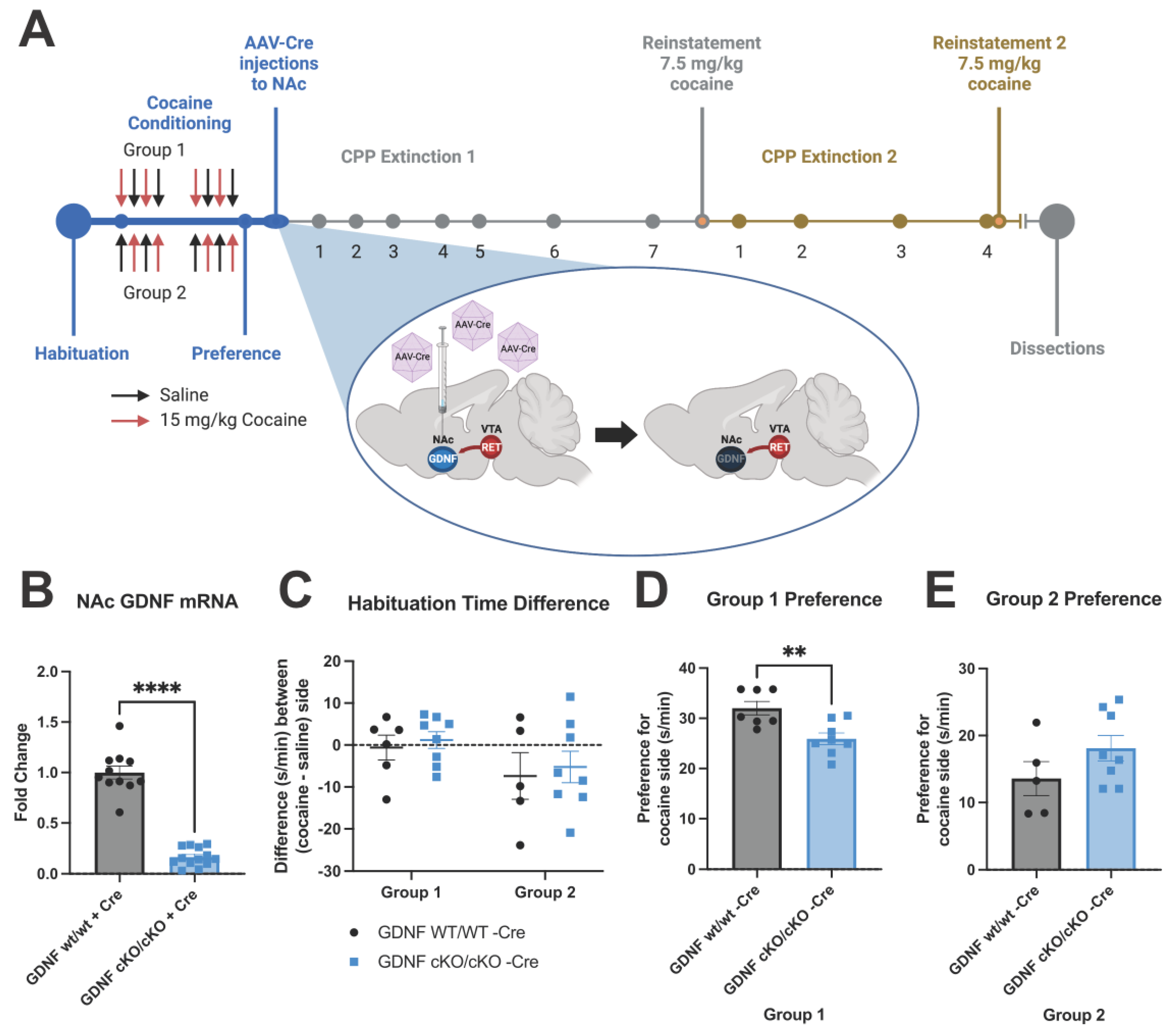
2.3. Intracranial AAV-Cre Injections
2.4. Conditioned Place Preference
2.5. Dissections
2.6. Reverse Transcription and Quantitative PCR
2.7. Statistical Analysis
3. Results
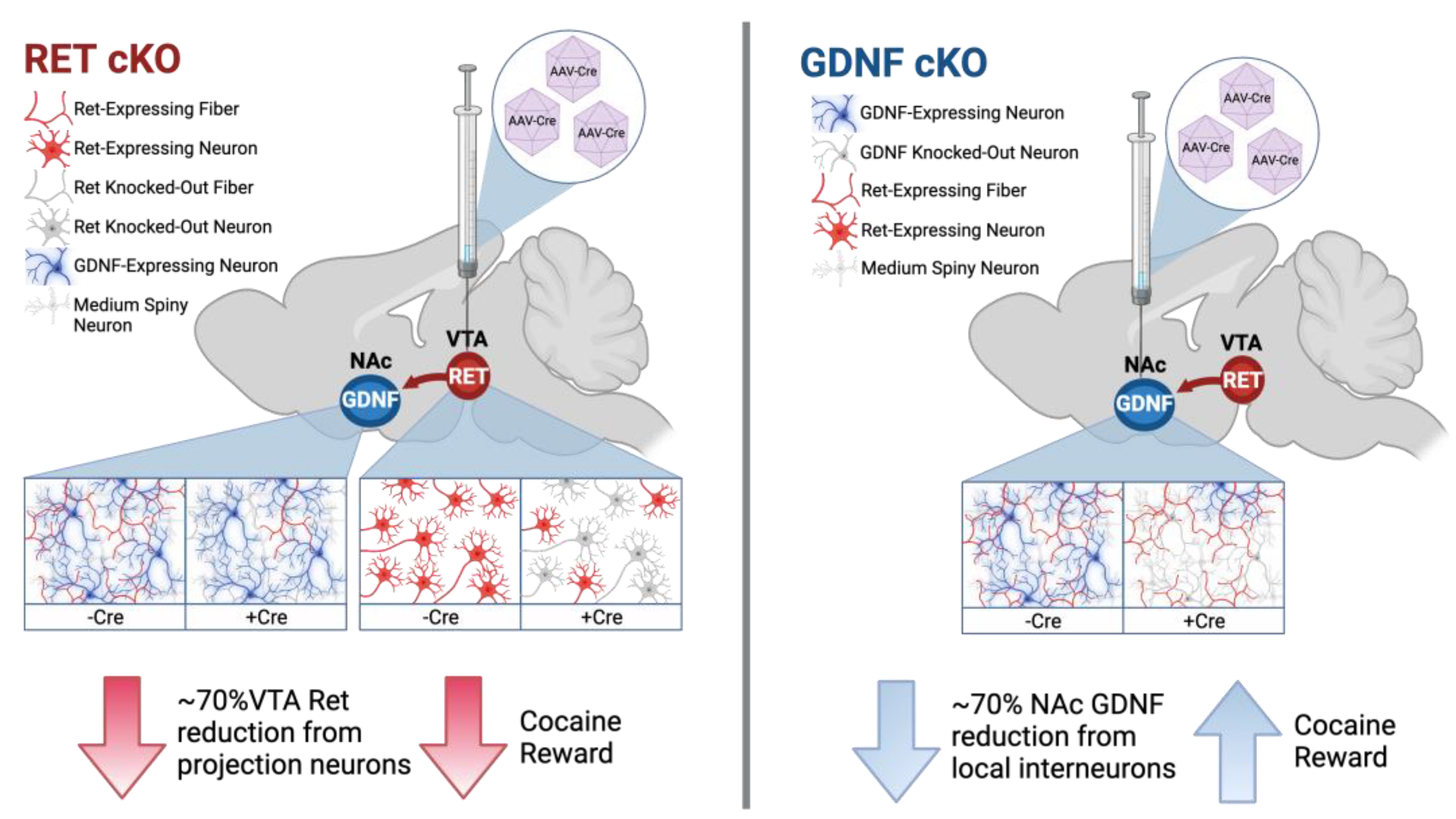
3.1. RET cKO Mice upon AAV-Cre Delivery Have about 60–70% Reduction of RET from the VTA

3.2. Animals with about 60–70% Reduction of VTA RET Have Hastened Extinction and Decreased Preference upon Cocaine Reinstatement
3.3. GDNF cKO Mice upon AAV-Cre Delivery Have about 70–80% Reduction of GDNF from the NAc
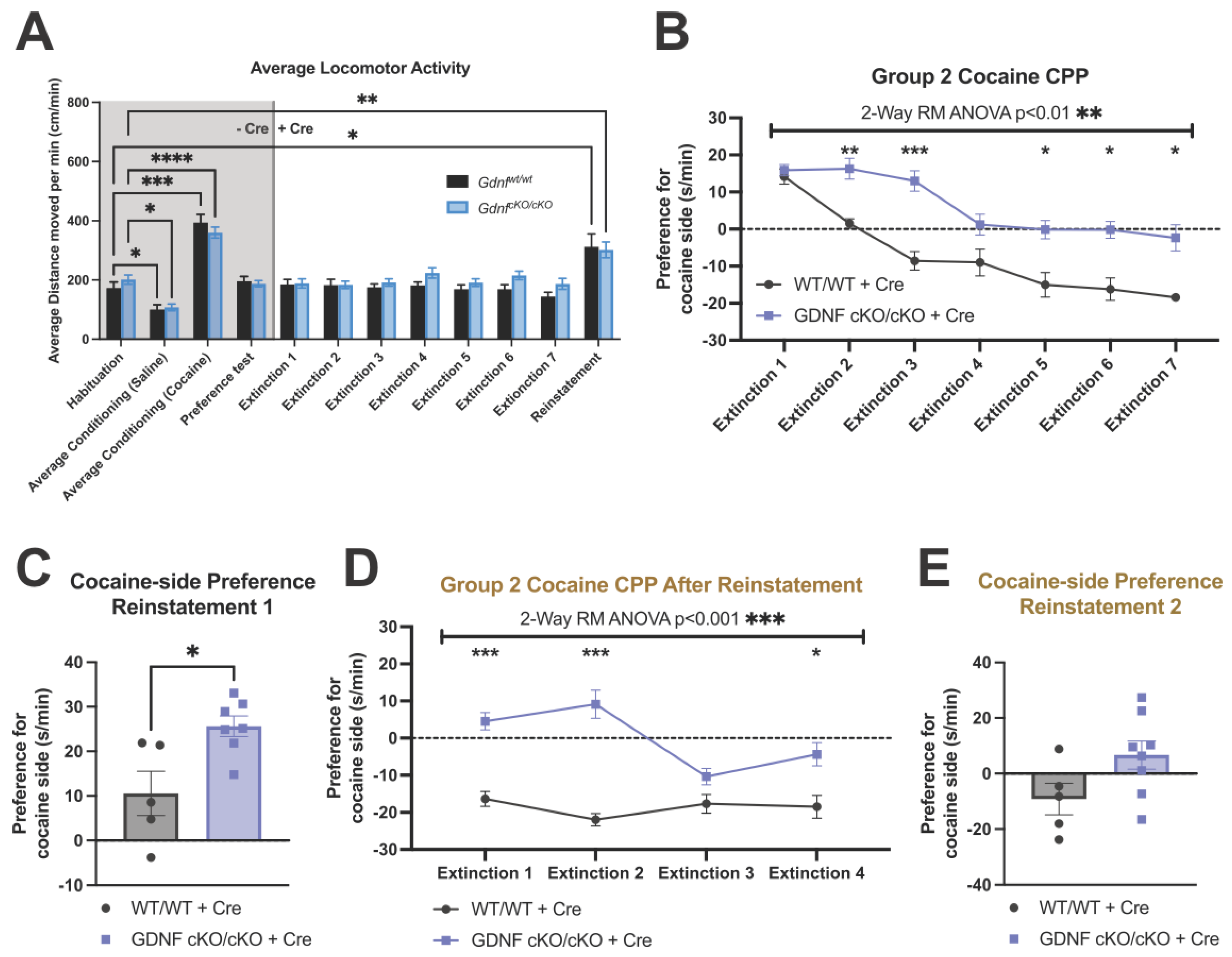
3.4. No Differences in Locomotor Activity Were Observed between the Mutants and Controls
3.5. Animals with about 70% Reduction in NAc GDNF Have Prolonged Extinction and Increased Preference upon Cocaine Reinstatement
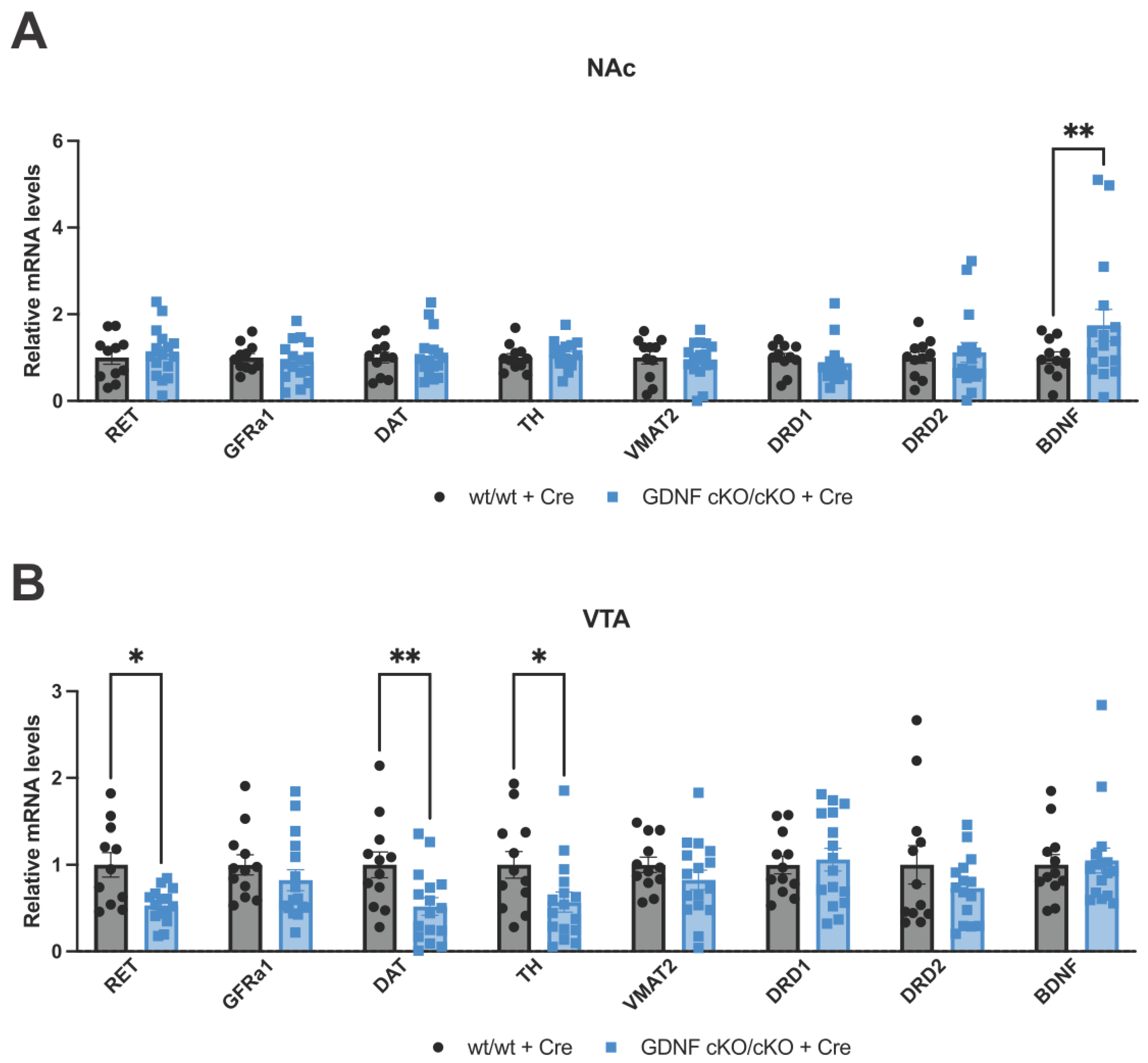
3.6. Analysis of mRNA Levels of Key Dopamine-Related and Plasticity Genes in the NAc and VTA after 70–80% GDNF Reduction in the NAc
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peacock, A.; Leung, J.; Larney, S.; Colledge, S.; Hickman, M.; Rehm, J.; Giovino, G.A.; West, R.; Hall, W.; Griffiths, P.; et al. Global statistics on alcohol, tobacco and illicit drug use: 2017 status report. Addiction 2018, 113, 1905–1926. [Google Scholar] [CrossRef]
- Bachi, K.; Mani, V.; Jeyachandran, D.; Fayad, Z.A.; Goldstein, R.Z.; Alia-Klein, N. Vascular disease in cocaine addiction. Atherosclerosis 2017, 262, 154–162. [Google Scholar] [CrossRef]
- Sulzer, D. How Addictive Drugs Disrupt Presynaptic Dopamine Neurotransmission. Neuron 2011, 69, 628–649. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Tilley, M.R.; Wei, H.; Zhou, F.; Zhou, F.-M.; Ching, S.; Quan, N.; Stephens, R.L.; Hill, E.R.; Nottoli, T.; et al. Abolished cocaine reward in mice with a cocaine-insensitive dopamine transporter. Proc. Natl. Acad. Sci. USA 2006, 103, 9333–9338. [Google Scholar] [CrossRef] [PubMed]
- Witkiewitz, K.; Litten, R.Z.; Leggio, L. Advances in the science and treatment of alcohol use disorder. Sci. Adv. 2019, 5, eaax4043. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Jones, E.B.; Einstein, E.B.; Wargo, E.M. Prevention and Treatment of Opioid Misuse and Addiction. JAMA Psychiatry 2019, 76, 208–216. [Google Scholar] [CrossRef]
- Chan, B.; Kondo, K.; Freeman, M.; Ayers, C.; Montgomery, J.; Kansagara, D. Pharmacotherapy for Cocaine Use Disorder—A Systematic Review and Meta-analysis. J. Gen. Intern. Med. 2019, 34, 2858–2873. [Google Scholar] [CrossRef]
- Bentzley, B.S.; Han, S.S.; Neuner, S.; Humphreys, K.; Kampman, K.M.; Halpern, C.H. Comparison of Treatments for Cocaine Use Disorder Among Adults. JAMA Netw. Open 2021, 4, e218049. [Google Scholar] [CrossRef]
- Bardo, M.T.; Bevins, R.A. Conditioned place preference: What does it add to our preclinical understanding of drug reward? Psychopharmacology 2000, 153, 31–43. [Google Scholar] [CrossRef]
- Tzschentke, T.M. Measuring reward with the conditioned place preference paradigm: A comprehensive review of drug effects, recent progress and new issues. Prog. Neurobiol. 1998, 56, 613–672. [Google Scholar] [CrossRef]
- Cunningham, C.L.; Gremel, C.M.; Groblewski, P.A. Drug-induced conditioned place preference and aversion in mice. Nat. Protoc. 2006, 1, 1662–1670. [Google Scholar] [CrossRef]
- Huston, J.P.; de Souza Silva, M.A.; Topic, B.; Müller, C.P. What’s conditioned in conditioned place preference? Trends Pharmacol. Sci. 2013, 34, 162–166. [Google Scholar] [CrossRef]
- Itzhak, Y.; Martin, J.L. Cocaine-induced Conditioned Place Preference in Mice Induction, Extinction and Reinstatement by Related Psychostimulants. Neuropsychopharmacology 2002, 26, 130–134. [Google Scholar] [CrossRef]
- Hiroi, N.; White, N.M. The amphetamine conditioned place preference: Differential involvement of dopamine receptor subtypes and two dopaminergic terminal areas. Brain Res. 1991, 552, 141–152. [Google Scholar] [CrossRef]
- Martinez, L.A.A.; Curis, E.; Mekdad, N.; Larrieu, C.; Courtin, C.; Jourdren, L.; Blugeon, C.; Laplanche, J.-L.; Megarbane, B.; Marie-Claire, C.; et al. Individual differences in cocaine-induced conditioned place preference in male rats: Behavioral and transcriptomic evidence. J. Psychopharmacol. 2022, 36, 1161–1175. [Google Scholar] [CrossRef]
- Shaw, J.K.; Alonso, I.P.; Lewandowski, S.I.; Scott, M.O.; O’connor, B.M.; Aggarwal, S.; De Biasi, M.; Mortensen, O.V.; España, R.A. Individual differences in dopamine uptake in the dorsomedial striatum prior to cocaine exposure predict motivation for cocaine in male rats. Neuropsychopharmacology 2021, 46, 1757–1767. [Google Scholar] [CrossRef]
- Wang, X.; Gallegos, D.A.; Pogorelov, V.M.; O’hare, J.K.; Calakos, N.; Wetsel, W.C.; West, A.E. Parvalbumin Interneurons of the Mouse Nucleus Accumbens are Required for Amphetamine-Induced Locomotor Sensitization and Conditioned Place Preference. Neuropsychopharmacology 2018, 43, 953–963. [Google Scholar] [CrossRef]
- Borges, A.C.; Duarte, R.B.; Nogueira, L.; Barros, M. Temporal and dose-dependent differences in simultaneously-induced cocaine hypervigilance and conditioned-place-preference in marmoset monkeys. Drug Alcohol Depend. 2015, 148, 188–194. [Google Scholar] [CrossRef]
- Childs, E.; de Wit, H. Amphetamine-Induced Place Preference in Humans. Biol. Psychiatry 2009, 65, 900–904. [Google Scholar] [CrossRef]
- Hoffer, B.J.; Hoffman, A.; Bowenkamp, K.; Huettl, P.; Hudson, J.; Martin, D.; Lin, L.-F.H.; Gerhardt, G.A. Glial cell line-derived neurotrophic factor reverses toxin-induced injury to midbrain dopaminergic neurons in vivo. Neurosci. Lett. 1994, 182, 107–111. [Google Scholar] [CrossRef]
- Lin, L.-F.H.; Doherty, D.H.; Lile, J.D.; Bektesh, S.; Collins, F. GDNF: A Glial Cell Line-Derived Neurotrophic Factor for Midbrain Dopaminergic Neurons. Science 1993, 260, 1130–1132. [Google Scholar] [CrossRef] [PubMed]
- Kordower, J.H.; Emborg, M.E.; Bloch, J.; Ma, S.Y.; Chu, Y.; Leventhal, L.; McBride, J.; Chen, E.Y.; Palfi, S.; Roitberg, B.Z.; et al. Neurodegeneration prevented by lentiviral vector delivery of GDNF in primate models of Parkinson's disease. Science 2000, 290, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Trupp, M.; Belluardo, N.; Funakoshi, H.; Ibáñez, C.F. Complementary and Overlapping Expression of Glial Cell Line-Derived Neurotrophic Factor (GDNF), c-ret Proto-Oncogene, and GDNF Receptor-α Indicates Multiple Mechanisms of Trophic Actions in the Adult Rat CNS. J. Neurosci. 1997, 17, 3554–3567. [Google Scholar] [CrossRef] [PubMed]
- Golden, J.P.; DeMaro, J.A.; Osborne, P.A.; Milbrandt, J.; Johnson, E.M. Expression of Neurturin, GDNF, and GDNF Family-Receptor mRNA in the Developing and Mature Mouse. Exp. Neurol. 1999, 158, 504–528. [Google Scholar] [CrossRef]
- Hidalgo-Figueroa, M.; Bonilla, S.; Gutiérrez, F.; Pascual, A.; López-Barneo, J. GDNF Is Predominantly Expressed in the PV+ Neostriatal Interneuronal Ensemble in Normal Mouse and after Injury of the Nigrostriatal Pathway. J. Neurosci. 2012, 32, 864–872. [Google Scholar] [CrossRef]
- Pascual, A.; Hidalgo-Figueroa, M.; Piruat, J.I.; Pintado, C.O.; Gómez-Díaz, R.; López-Barneo, J. Absolute requirement of GDNF for adult catecholaminergic neuron survival. Nat. Neurosci. 2008, 11, 755–761. [Google Scholar] [CrossRef]
- Durbec, P.; Marcos-Gutierrez, C.V.; Kilkenny, C.; Grigoriou, M.; Wartiowaara, K.; Suvanto, P.; Smith, D.; Ponder, B.; Costantini, F.; Saarma, M.; et al. GDNF signalling through the Ret receptor tyrosine kinase. Nature 1996, 381, 789–793. [Google Scholar] [CrossRef]
- Wang, J.; Carnicella, S.; Ahmadiantehrani, S.; He, D.-Y.; Barak, S.; Kharazia, V.; Ben Hamida, S.; Zapata, A.; Shippenberg, T.S.; Ron, D. Nucleus Accumbens-Derived Glial Cell Line-Derived Neurotrophic Factor Is a Retrograde Enhancer of Dopaminergic Tone in the Mesocorticolimbic System. J. Neurosci. 2010, 30, 14502–14512. [Google Scholar] [CrossRef]
- Wallén, A.; Castro, D.; Zetterström, R.; Karlén, M.; Olson, L.; Ericson, J.; Perlmann, T. Orphan Nuclear Receptor Nurr1 Is Essential for Ret Expression in Midbrain Dopamine Neurons and in the Brain Stem. Mol. Cell. Neurosci. 2001, 18, 649–663. [Google Scholar] [CrossRef]
- Trupp, M.; Arenas, E.; Fainzilber, M.; Nilsson, A.-S.; Sieber, B.-A.; Grigoriou, M.; Kilkenny, C.; Salazar-Grueso, E.; Pachnis, V.; Arumäe, U.; et al. Functional receptor for GDNF encoded by the c-ret proto-oncogene. Nature 1996, 381, 785–789. [Google Scholar] [CrossRef]
- Lüscher, C.; Malenka, R.C. Drug-Evoked Synaptic Plasticity in Addiction: From Molecular Changes to Circuit Remodeling. Neuron 2011, 69, 650–663. [Google Scholar] [CrossRef]
- Airaksinen, M.S.; Saarma, M. The GDNF family: Signalling, biological functions and therapeutic value. Nat. Rev. Neurosci. 2002, 3, 383–394. [Google Scholar] [CrossRef]
- Ibáñez, C.F.; Andressoo, J.-O. Biology of GDNF and its receptors—Relevance for disorders of the central nervous system. Neurobiol. Dis. 2017, 97, 80–89. [Google Scholar] [CrossRef]
- Drinkut, A.; Tillack, K.; Meka, D.P.; Schulz, J.B.; Kügler, S.; Kramer, E.R. Ret is essential to mediate GDNF’s neuroprotective and neuroregenerative effect in a Parkinson disease mouse model. Cell Death Dis. 2016, 7, e2359. [Google Scholar] [CrossRef]
- Taraviras, S.; Marcos-Gutierrez, C.V.; Durbec, P.; Jani, H.; Grigoriou, M.; Sukumaran, M.; Wang, L.-C.; Hynes, M.; Raisman, G.; Pachnis, V. Signalling by the RET receptor tyrosine kinase and its role in the development of the mammalian enteric nervous system. Development 1999, 126, 2785–2797. [Google Scholar] [CrossRef]
- Jing, S.; Wen, D.; Yu, Y.; Holst, P.L.; Luo, Y.; Fang, M.; Tamir, R.; Antonio, L.; Hu, Z.; Cupples, R.; et al. GDNF–Induced Activation of the Ret Protein Tyrosine Kinase Is Mediated by GDNFR-α, a Novel Receptor for GDNF. Cell 1996, 85, 1113–1124. [Google Scholar] [CrossRef]
- Yang, F.; Feng, L.; Zheng, F.; Johnson, S.W.; Du, J.; Shen, L.; Wu, C.-P.; Lu, B. GDNF acutely modulates excitability and A-type K+ channels in midbrain dopaminergic neurons. Nat. Neurosci. 2001, 4, 1071–1078. [Google Scholar] [CrossRef]
- Mätlik, K.; Garton, D.R.; Montaño-Rodríguez, A.R.; Olfat, S.; Eren, F.; Casserly, L.; Damdimopoulos, A.; Panhelainen, A.; Porokuokka, L.L.; Kopra, J.J.; et al. Elevated endogenous GDNF induces altered dopamine signalling in mice and correlates with clinical severity in schizophrenia. Mol. Psychiatry 2022, 27, 3247–3261. [Google Scholar] [CrossRef]
- Kumar, A.; Kopra, J.; Varendi, K.; Porokuokka, L.L.; Panhelainen, A.; Kuure, S.; Marshall, P.; Karalija, N.; Härma, M.-A.; Vilenius, C.; et al. GDNF Overexpression from the Native Locus Reveals its Role in the Nigrostriatal Dopaminergic System Function. PLOS Genet. 2015, 11, e1005710. [Google Scholar] [CrossRef]
- Kopra, J.J.; Panhelainen, A.; Bjerkén, S.A.; Porokuokka, L.L.; Varendi, K.; Olfat, S.; Montonen, H.; Piepponen, T.P.; Saarma, M.; Andressoo, J.-O. Dampened Amphetamine-Stimulated Behavior and Altered Dopamine Transporter Function in the Absence of Brain GDNF. J. Neurosci. 2017, 37, 1581–1590. [Google Scholar] [CrossRef]
- Morel, L.; Domingues, O.; Zimmer, J.; Michel, T. Revisiting the Role of Neurotrophic Factors in Inflammation. Cells 2020, 9, 865. [Google Scholar] [CrossRef] [PubMed]
- Mijatovic, J.; Airavaara, M.; Planken, A.; Auvinen, P.; Raasmaja, A.; Piepponen, T.P.; Costantini, F.; Ahtee, L.; Saarma, M. Constitutive Ret Activity in Knock-In Multiple Endocrine Neoplasia Type B Mice Induces Profound Elevation of Brain Dopamine Concentration via Enhanced Synthesis and Increases the Number of TH-Positive Cells in the Substantia Nigra. J. Neurosci. 2007, 27, 4799–4809. [Google Scholar] [CrossRef] [PubMed]
- Smith-Hicks, C.L.; Sizer, K.C.; Powers, J.; Tischler, A.; Costantini, F. C-cell hyperplasia, pheochromocytoma and sympathoadrenal malformation in a mouse model of multiple endocrine neoplasia type 2B. EMBO J. 2000, 19, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Kopra, J.; Villarta-Aguilera, M.; Savolainen, M.; Weingerl, S.; Myöhänen, T.T.; Rannanpää, S.; Salvatore, M.F.; Andressoo, J.-O.; Piepponen, T.P. Constitutive Ret signaling leads to long-lasting expression of amphetamine-induced place conditioning via elevation of mesolimbic dopamine. Neuropharmacology 2018, 128, 221–230. [Google Scholar] [CrossRef]
- Messer, C.J.; Eisch, A.J.; A Carlezon, W.; Whisler, K.; Shen, L.; Wolf, D.H.; Westphal, H.; Collins, F.; Russell, D.S.; Nestler, E.J. Role for GDNF in Biochemical and Behavioral Adaptations to Drugs of Abuse. Neuron 2000, 26, 247–257. [Google Scholar] [CrossRef]
- Koskela, M.; Piepponen, T.P.; Lindahl, M.; Harvey, B.K.; Andressoo, J.-O.; Võikar, V.; Airavaara, M. The overexpression of GDNF in nucleus accumbens suppresses alcohol-seeking behavior in group-housed C57Bl/6J female mice. J. Biomed. Sci. 2021, 28, 1–17. [Google Scholar] [CrossRef]
- Carnicella, S.; Ron, D. GDNF—A potential target to treat addiction. Pharmacol. Ther. 2009, 122, 9–18. [Google Scholar] [CrossRef]
- Luo, W.; Wickramasinghe, S.R.; Savitt, J.M.; Griffin, J.W.; Dawson, T.M.; Ginty, D.D. A Hierarchical NGF Signaling Cascade Controls Ret-Dependent and Ret-Independent Events during Development of Nonpeptidergic DRG Neurons. Neuron 2007, 54, 739–754. [Google Scholar] [CrossRef]
- Kopra, J.; Vilenius, C.; Grealish, S.; Härma, M.-A.; Varendi, K.; Lindholm, J.; Castrén, E.; Võikar, V.; Björklund, A.; Piepponen, T.P.; et al. GDNF is not required for catecholaminergic neuron survival in vivo. Nat. Neurosci. 2015, 18, 319–322. [Google Scholar] [CrossRef]
- Fleming, W.; Lee, J.; Briones, B.A.; Bolkan, S.S.; Witten, I.B. Cholinergic interneurons mediate cocaine extinction in male mice through plasticity across medium spiny neuron subtypes. Cell Rep. 2022, 39, 110874. [Google Scholar] [CrossRef]
- Cunningham, C.L.; Ferree, N.K.; Howard, M.A. Apparatus bias and place conditioning with ethanol in mice. Psychopharmacology 2003, 170, 409–422. [Google Scholar] [CrossRef]
- Olfat, S.; Mätlik, K.; Kopra, J.J.; Garton, D.R.; Iivanainen, V.H.; Bhattacharya, D.; Jakobsson, J.; Piepponen, T.P.; Andressoo, J.-O. Increased Physiological GDNF Levels Have No Effect on Dopamine Neuron Protection and Restoration in a Proteasome Inhibition Mouse Model of Parkinson’s Disease. Eneuro 2023, 10, 2. [Google Scholar] [CrossRef]
- McGinty, J.F.; Whitfield, T.W., Jr.; Berglind, W.J. Brain-derived neurotrophic factor and cocaine addiction. Brain Res. 2010, 1314, 183–193. [Google Scholar] [CrossRef]
- Lu, B.C.; Cebrian, C.; Chi, X.; Kuure, S.; Kuo, R.; Bates, C.M.; Arber, S.; Hassell, J.; MacNeil, L.; Hoshi, M.; et al. Etv4 and Etv5 are required downstream of GDNF and Ret for kidney branching morphogenesis. Nat. Genet. 2009, 41, 1295–1302. [Google Scholar] [CrossRef]
- Ola, R.; Jakobson, M.; Kvist, J.; Perälä, N.; Kuure, S.; Braunewell, K.-H.; Bridgewater, D.; Rosenblum, N.D.; Chilov, D.; Immonen, T.; et al. The GDNF Target Vsnl1 Marks the Ureteric Tip. J. Am. Soc. Nephrol. 2011, 22, 274–284. [Google Scholar] [CrossRef]
- Widenfalk, J.; Nosrat, C.; Tomac, A.; Westphal, H.; Hoffer, B.; Olson, L. Neurturin and Glial Cell Line-Derived Neurotrophic Factor Receptor-β (GDNFR-β), Novel Proteins Related to GDNF and GDNFR-α with Specific Cellular Patterns of Expression Suggesting Roles in the Developing and Adult Nervous System and in Peripheral Organs. J. Neurosci. 1997, 17, 8506–8519. [Google Scholar] [CrossRef]
- Horger, B.A.; Nishimura, M.C.; Armanini, M.P.; Wang, L.-C.; Poulsen, K.T.; Rosenblad, C.; Kirik, D.; Moffat, B.; Simmons, L.; Johnson, E., Jr.; et al. Neurturin Exerts Potent Actions on Survival and Function of Midbrain Dopaminergic Neurons. J. Neurosci. 1998, 18, 4929–4937. [Google Scholar] [CrossRef]
- Green-Sadan, T.; Kinor, N.; Roth-Deri, I.; Geffen-Aricha, R.; Schindler, C.J.; Yadid, G. Transplantation of glial cell line-derived neurotrophic factor-expressing cells into the striatum and nucleus accumbens attenuates acquisition of cocaine self-administration in rats. Eur. J. Neurosci. 2003, 18, 2093–2098. [Google Scholar] [CrossRef]
- Green-Sadan, T.; Kuttner, Y.; Lublin-Tennenbaum, T.; Kinor, N.; Boguslavsky, Y.; Margel, S.; Yadid, G. Glial cell line-derived neurotrophic factor-conjugated nanoparticles suppress acquisition of cocaine self-administration in rats. Exp. Neurol. 2005, 194, 97–105. [Google Scholar] [CrossRef]
- Lu, L.; Wang, X.; Wu, P.; Xu, C.; Zhao, M.; Morales, M.; Harvey, B.K.; Hoffer, B.J.; Shaham, Y. Role of Ventral Tegmental Area Glial Cell Line–Derived Neurotrophic Factor in Incubation of Cocaine Craving. Biol. Psychiatry 2009, 66, 137–145. [Google Scholar] [CrossRef]
- Ghitza, U.E.; Zhai, H.; Wu, P.; Airavaara, M.; Shaham, Y.; Lu, L. Role of BDNF and GDNF in drug reward and relapse: A review. Neurosci. Biobehav. Rev. 2010, 35, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.L.; Edwards, S.; Bachtell, R.; DiLeone, R.; Rios, M.; Self, D.W. Dynamic BDNF activity in nucleus accumbens with cocaine use increases self-administration and relapse. Nat. Neurosci. 2007, 10, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.L.; Krishnan, V.; Larson, E.; Graham, A.; Edwards, S.; Bachtell, R.; Simmons, D.; Gent, L.M.; Berton, O.; Bolanos, C.A.; et al. Tropomyosin-Related Kinase B in the Mesolimbic Dopamine System: Region-Specific Effects on Cocaine Reward. Biol. Psychiatry 2009, 65, 696–701. [Google Scholar] [CrossRef] [PubMed]
- Hall, F.S.; Drgonova, J.; Goeb, M.; Uhl, G.R. Reduced Behavioral Effects of Cocaine in Heterozygous Brain-Derived Neurotrophic Factor (BDNF) Knockout Mice. Neuropsychopharmacology 2003, 28, 1485–1490. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wolf, M.E. Multiple faces of BDNF in cocaine addiction. Behav. Brain Res. 2014, 279, 240–254. [Google Scholar] [CrossRef]
- Bahi, A.; Boyer, F.; Chandrasekar, V.; Dreyer, J.-L. Role of accumbens BDNF and TrkB in cocaine-induced psychomotor sensitization, conditioned-place preference, and reinstatement in rats. Psychopharmacology 2008, 200, 155. [Google Scholar] [CrossRef]
- Horger, B.A.; Iyasere, C.A.; Berhow, M.T.; Messer, C.J.; Nestler, E.J.; Taylor, J.R. Enhancement of Locomotor Activity and Conditioned Reward to Cocaine by Brain-Derived Neurotrophic Factor. J. Neurosci. 1999, 19, 4110–4122. [Google Scholar] [CrossRef]
- O'Neal, T.J.; Bernstein, M.X.; MacDougall, D.J.; Ferguson, S.M. A Conditioned Place Preference for Heroin Is Signaled by Increased Dopamine and Direct Pathway Activity and Decreased Indirect Pathway Activity in the Nucleus Accumbens. J. Neurosci. 2022, 42, 2011–2024. [Google Scholar] [CrossRef]
- Richardson, D.S.; Lai, A.Z.; Mulligan, L.M. RET ligand-induced internalization and its consequences for downstream signaling. Oncogene 2006, 25, 3206–3211. [Google Scholar] [CrossRef]
- Yang, L.X.; Nelson, P.G. Glia cell line-derived neurotrophic factor regulates the distribution of acetylcholine receptors in mouse primary skeletal muscle cells. Neuroscience 2004, 128, 497–509. [Google Scholar] [CrossRef]
- Di Liberto, V.; Mudò, G.; Belluardo, N. mGluR2/3 agonist LY379268, by enhancing the production of GDNF, induces a time-related phosphorylation of RET receptor and intracellular signaling Erk1/2 in mouse striatum. Neuropharmacology 2011, 61, 638–645. [Google Scholar] [CrossRef]
- Kramer, E.R.; Liss, B. GDNF-Ret signaling in midbrain dopaminergic neurons and its implication for Parkinson disease. FEBS Lett. 2015, 589, 3760–3772. [Google Scholar] [CrossRef]
- Kampman, K.M. New medications for the treatment of cocaine dependence. Psychiatry 2005, 2, 44–48. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward Sequence | Reverse Sequence |
|---|---|---|
| Ret cKO | ACT CCT TGG GCC TGC TGA G | GAG GCA GGA AGG CCT GTG |
| Gdnf cKO | 5′: CTC ATT TCC CAC AGG GAA CTG 3′: GAA ACC AAG GAG GAA CTG ATC | 3′: TCT TCT GCC TCT GCC TCC G |
| Primer | Forward Sequence | Reverse Sequence |
|---|---|---|
| mRet | TCC CTT CCA CAT GGA TTG A | ATC GGC TCT CGT GAG TGG TA |
| mGDNF | CGC TGA CCA GTG ACT CCA ATA TGC | TGC CGC TTG TTT ATC TGG TGA CC |
| mGfra1 | TTC CCA CAC ACG TTT TAC CA | GCC CGA TAC ATT GGA TTT CA |
| mTh | CCC AAG GGC TTC AGA AGA G | GGG CAT CCT CGA TGA GAC T |
| mVmat2 | ATG CTG CTC ACC GTC GTA GT | TTT TTC TCG TGC TTA ATG CTG T |
| mDat | AAC CTG TAC TGG CGG CTA TG | GCT GAC CAC GAC CAC TAC A |
| mDrd2 | ACA CAC CGT ACA GCT CCA AG | GGA GTA GAC GAC CAC GAA GGC AG |
| mDrd1 | GCG TGG TCT CCC AGA TCG | GCA TTT CTC CTT CAA GCC CCT |
| mBdnf | GGC CCA ACG AAG AAA ACC AT | AGC ATC ACC CGG GAA GTG T |
| mbActin | CTA AGG CCA ACC CTG AAA AG | ACC AGA GGC ATA CAG GGA CA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garton, D.R.; Turconi, G.; Iivanainen, V.; Andressoo, J.-O. Opposing Spatially Segregated Function of Endogenous GDNF-RET Signaling in Cocaine Addiction. Biomolecules 2023, 13, 761. https://doi.org/10.3390/biom13050761
Garton DR, Turconi G, Iivanainen V, Andressoo J-O. Opposing Spatially Segregated Function of Endogenous GDNF-RET Signaling in Cocaine Addiction. Biomolecules. 2023; 13(5):761. https://doi.org/10.3390/biom13050761
Chicago/Turabian StyleGarton, Daniel R., Giorgio Turconi, Vilma Iivanainen, and Jaan-Olle Andressoo. 2023. "Opposing Spatially Segregated Function of Endogenous GDNF-RET Signaling in Cocaine Addiction" Biomolecules 13, no. 5: 761. https://doi.org/10.3390/biom13050761
APA StyleGarton, D. R., Turconi, G., Iivanainen, V., & Andressoo, J. -O. (2023). Opposing Spatially Segregated Function of Endogenous GDNF-RET Signaling in Cocaine Addiction. Biomolecules, 13(5), 761. https://doi.org/10.3390/biom13050761