
Sphingosine-1-Phosphate Signaling in Cardiovascular Diseases


Abstract
:1. Introduction
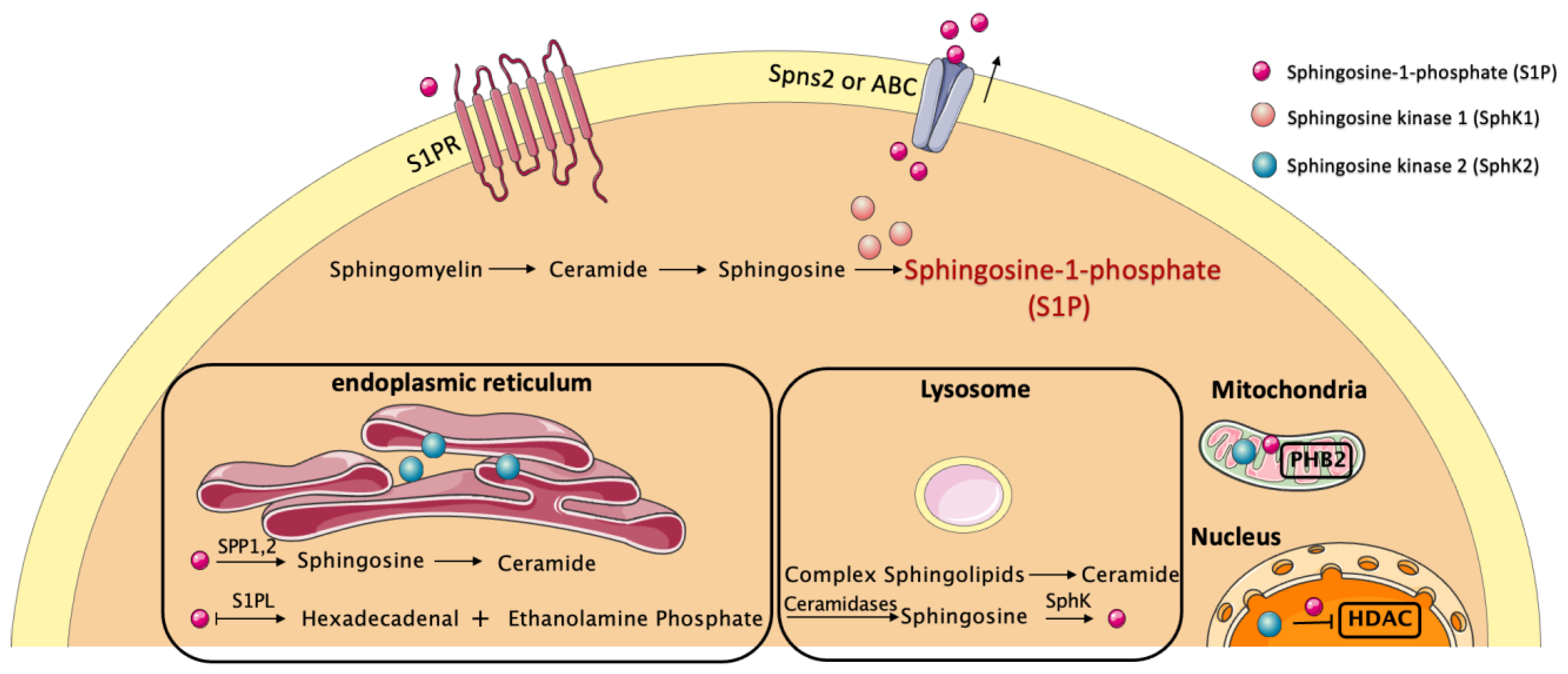
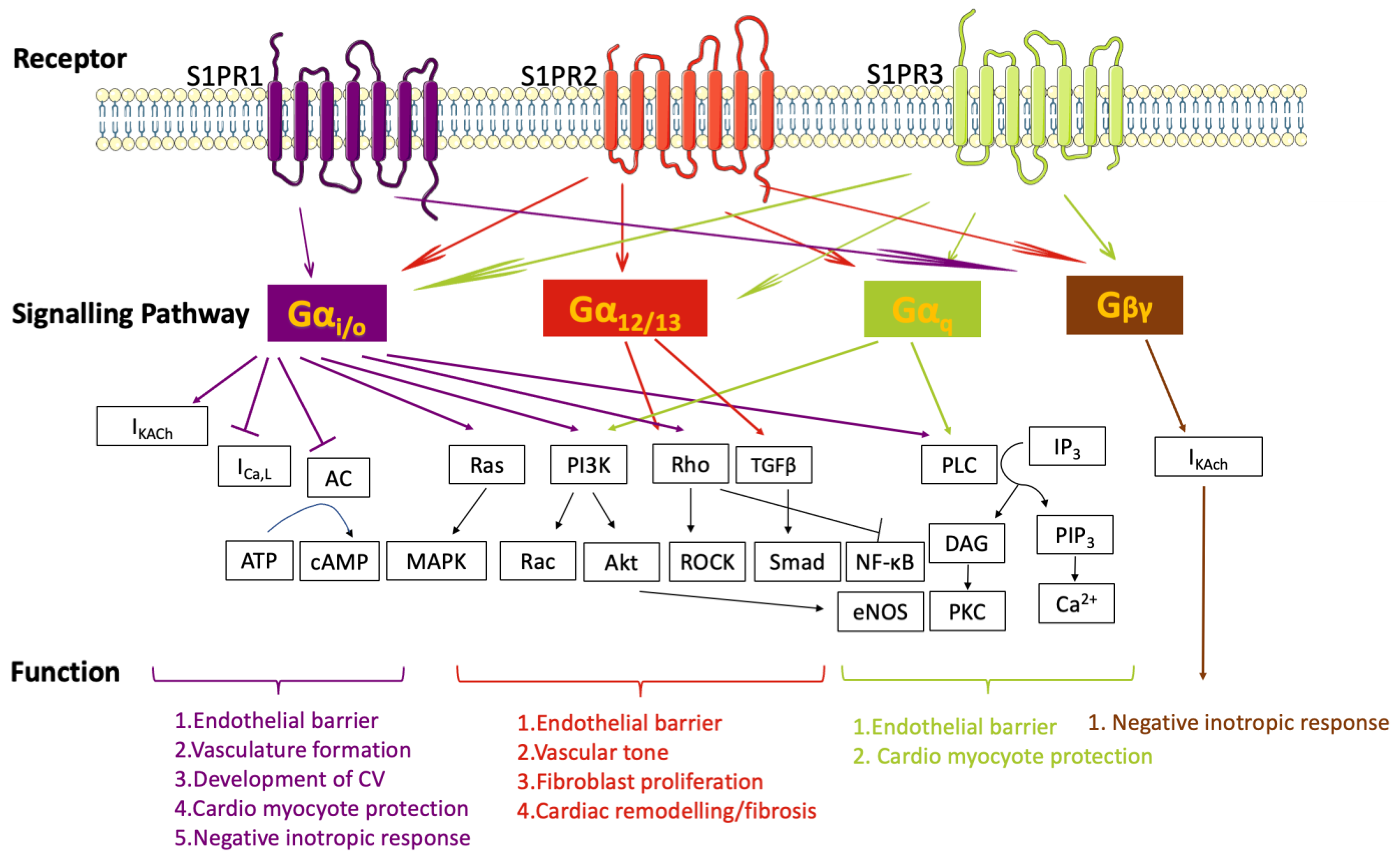
2. Regulatory Network of S1P and S1PRs
3. S1P and Cardiovascular Functions
3.1. Effects of S1P in Blood Vessels
3.2. Effects of S1P in the Heart
4. S1P and Cardiovascular Diseases
4.1. Anti-Atherosclerotic Effect of S1P
4.2. S1P Promotes Angiogenesis
4.3. Complex Action of S1P in Vascular Tone Control and Hypertension
4.4. S1P Reduces Myocardial Ischemia and Reinfusion Injury
4.5. Cardioprotective Effects of S1P in Myocardial Infarction, Fibrosis and Heart Failure
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stoffel, W.; Sticht, G.; LeKim, D. Metabolism of sphingosine bases. Ix. Degradation in vitro of dihydrospingosine and dihydrospingosine phosphate to palmitaldehyde and ethanolamine phosphate. Hoppe Seylers Z. Physiol. Chem. 1968, 349, 1745–1748. [Google Scholar] [CrossRef] [PubMed]
- Yatomi, Y.; Ozaki, Y.; Ohmori, T.; Igarashi, Y. Sphingosine 1-phosphate: Synthesis and release. Prostaglandins Other Lipid Mediat. 2001, 64, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Perez-Jeldres, T.; Alvarez-Lobos, M.; Rivera-Nieves, J. Targeting sphingosine-1-phosphate signaling in immune-mediated diseases: Beyond multiple sclerosis. Drugs 2021, 81, 985–1002. [Google Scholar] [CrossRef] [PubMed]
- Maceyka, M.; Spiegel, S. Sphingolipid metabolites in inflammatory disease. Nature 2014, 510, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, S.; Milstien, S.; Grant, S. Endogenous modulators and pharmacological inhibitors of histone deacetylases in cancer therapy. Oncogene 2012, 31, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Hait, N.C.; Maiti, A. The role of sphingosine-1-phosphate and ceramide-1-phosphate in inflammation and cancer. Mediat. Inflamm. 2017, 2017, 4806541. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Bonapace, S.; Lunardi, G.; Canali, G.; Dugo, C.; Vinco, G.; Calabria, S.; Barbieri, E.; Laaksonen, R.; Bonnet, F.; et al. Associations between specific plasma ceramides and severity of coronary-artery stenosis assessed by coronary angiography. Diabetes Metab. 2020, 46, 150–157. [Google Scholar] [CrossRef]
- Spijkers, L.J.; van den Akker, R.F.; Janssen, B.J.; Debets, J.J.; De Mey, J.G.; Stroes, E.S.; van den Born, B.J.; Wijesinghe, D.S.; Chalfant, C.E.; MacAleese, L.; et al. Hypertension is associated with marked alterations in sphingolipid biology: A potential role for ceramide. PLoS ONE 2011, 6, e21817. [Google Scholar] [CrossRef]
- Li, N.; Zhang, F. Implication of sphingosin-1-phosphate in cardiovascular regulation. Front. Biosci. 2016, 21, 1296–1313. [Google Scholar] [CrossRef]
- Feuerborn, R.; Becker, S.; Potì, F.; Nagel, P.; Brodde, M.; Schmidt, H.; Christoffersen, C.; Ceglarek, U.; Burkhardt, R.; Nofer, J.R. High density lipoprotein (hdl)-associated sphingosine 1-phosphate (s1p) inhibits macrophage apoptosis by stimulating stat3 activity and survivin expression. Atherosclerosis 2017, 257, 29–37. [Google Scholar] [CrossRef]
- Li, S.; Chen, J.; Fang, X.; Xia, X. Sphingosine-1-phosphate activates the akt pathway to inhibit chemotherapy induced human granulosa cell apoptosis. Gynecol. Endocrinol. 2017, 33, 476–479. [Google Scholar] [CrossRef]
- Venkataraman, K.; Thangada, S.; Michaud, J.; Oo, M.L.; Ai, Y.; Lee, Y.M.; Wu, M.; Parikh, N.S.; Khan, F.; Proia, R.L.; et al. Extracellular export of sphingosine kinase-1a contributes to the vascular s1p gradient. Biochem. J. 2006, 397, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Nagahashi, M.; Yamada, A.; Aoyagi, T.; Allegood, J.; Wakai, T.; Spiegel, S.; Takabe, K. Sphingosine-1-phosphate in the lymphatic fluid determined by novel methods. Heliyon 2016, 2, e00219. [Google Scholar] [CrossRef] [PubMed]
- Le Stunff, H.; Milstien, S.; Spiegel, S. Generation and metabolism of bioactive sphingosine-1-phosphate. J. Cell. Biochem. 2004, 92, 882–899. [Google Scholar] [CrossRef] [PubMed]
- Tolksdorf, C.; Moritz, E.; Wolf, R.; Meyer, U.; Marx, S.; Bien-Moller, S.; Garscha, U.; Jedlitschky, G.; Rauch, B.H. Platelet-derived s1p and its relevance for the communication with immune cells in multiple human diseases. Int. J. Mol. Sci. 2022, 23, 10278. [Google Scholar] [CrossRef]
- Venkataraman, K.; Lee, Y.M.; Michaud, J.; Thangada, S.; Ai, Y.; Bonkovsky, H.L.; Parikh, N.S.; Habrukowich, C.; Hla, T. Vascular endothelium as a contributor of plasma sphingosine 1-phosphate. Circ. Res. 2008, 102, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Książek, M.; Chacińska, M.; Chabowski, A.; Baranowski, M. Sources, metabolism, and regulation of circulating sphingosine-1-phosphate. J. Lipid Res. 2015, 56, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Hisano, Y.; Kobayashi, N.; Yamaguchi, A.; Nishi, T. Mouse spns2 functions as a sphingosine-1-phosphate transporter in vascular endothelial cells. PLoS ONE 2012, 7, e38941. [Google Scholar] [CrossRef]
- Mendoza, A.; Bréart, B.; Ramos-Perez, W.D.; Pitt, L.A.; Gobert, M.; Sunkara, M.; Lafaille, J.J.; Morris, A.J.; Schwab, S.R. The transporter spns2 is required for secretion of lymph but not plasma sphingosine-1-phosphate. Cell Rep. 2012, 2, 1104–1110. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.M.; Ishizu, A.N.; Foo, J.C.; Toh, X.R.; Zhang, F.; Whee, D.M.; Torta, F.; Cazenave-Gassiot, A.; Matsumura, T.; Kim, S.; et al. Mfsd2b is essential for the sphingosine-1-phosphate export in erythrocytes and platelets. Nature 2017, 550, 524–528. [Google Scholar] [CrossRef] [PubMed]
- Takabe, K.; Kim, R.H.; Allegood, J.C.; Mitra, P.; Ramachandran, S.; Nagahashi, M.; Harikumar, K.B.; Hait, N.C.; Milstien, S.; Spiegel, S. Estradiol induces export of sphingosine 1-phosphate from breast cancer cells via abcc1 and abcg2. J. Biol. Chem. 2010, 285, 10477–10486. [Google Scholar] [CrossRef] [PubMed]
- Vogt, K.; Mahajan-Thakur, S.; Wolf, R.; Bröderdorf, S.; Vogel, C.; Böhm, A.; Ritter, C.A.; Gräler, M.; Oswald, S.; Greinacher, A.; et al. Release of platelet-derived sphingosine-1-phosphate involves multidrug resistance protein 4 (mrp4/abcc4) and is inhibited by statins. Thromb. Haemost. 2018, 118, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Le Stunff, H.; Peterson, C.; Liu, H.; Milstien, S.; Spiegel, S. Sphingosine-1-phosphate and lipid phosphohydrolases. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2002, 1582, 8–17. [Google Scholar] [CrossRef]
- Harris, C.M.; Mittelstadt, S.; Banfor, P.; Bousquet, P.; Duignan, D.B.; Gintant, G.; Hart, M.; Kim, Y.; Segreti, J. Sphingosine-1-phosphate (s1p) lyase inhibition causes increased cardiac s1p levels and bradycardia in rats. J. Pharmacol. Exp. Ther. 2016, 359, 151–158. [Google Scholar] [CrossRef]
- Giussani, P.; Maceyka, M.; Le Stunff, H.; Mikami, A.; Lepine, S.; Wang, E.; Kelly, S.; Merrill, A.H., Jr.; Milstien, S.; Spiegel, S. Sphingosine-1-phosphate phosphohydrolase regulates endoplasmic reticulum-to-golgi trafficking of ceramide. Mol. Cell. Biol. 2006, 26, 5055–5069. [Google Scholar] [CrossRef]
- Pyne, S.; Long, J.S.; Ktistakis, N.T.; Pyne, N.J. Lipid phosphate phosphatases and lipid phosphate signalling. Biochem. Soc. Trans. 2005, 33, 1370–1374. [Google Scholar] [CrossRef]
- Pyne, S.; Lee, S.C.; Long, J.; Pyne, N.J. Role of sphingosine kinases and lipid phosphate phosphatases in regulating spatial sphingosine 1-phosphate signalling in health and disease. Cell. Signal. 2009, 21, 14–21. [Google Scholar] [CrossRef]
- Chun, J.; Giovannoni, G.; Hunter, S.F. Sphingosine 1-phosphate receptor modulator therapy for multiple sclerosis: Differential downstream receptor signalling and clinical profile effects. Drugs 2021, 81, 207–231. [Google Scholar] [CrossRef]
- Shao, X.; Liu, K.; Fan, Y.; Ding, Z.; Chen, M.; Zhu, M.; Weinstein, L.S.; Li, H.; Li, H. Galphas relays sphingosine-1-phosphate receptor 1 signaling to stabilize vascular endothelial-cadherin at endothelial junctions to control mouse embryonic vascular integrity. J. Genet. Genom. 2015, 42, 613–624. [Google Scholar] [CrossRef]
- Landeen, L.K.; Dederko, D.A.; Kondo, C.S.; Hu, B.S.; Aroonsakool, N.; Haga, J.H.; Giles, W.R. Mechanisms of the negative inotropic effects of sphingosine-1-phosphate on adult mouse ventricular myocytes. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H736–H749. [Google Scholar] [CrossRef]
- Means, C.K.; Miyamoto, S.; Chun, J.; Brown, J.H. S1p1 receptor localization confers selectivity for gi-mediated camp and contractile responses. J. Biol. Chem. 2008, 283, 11954–11963. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; He, L.; Gan, B.; Ti, R.; Xiao, Q.; Yang, X.; Hu, H.; Zhu, L.; Wang, S.; Ren, R. Structural insights into sphingosine-1-phosphate receptor activation. Proc. Natl. Acad. Sci. USA 2022, 119, e2117716119. [Google Scholar] [CrossRef]
- Xu, Z.; Ikuta, T.; Kawakami, K.; Kise, R.; Qian, Y.; Xia, R.; Sun, M.X.; Zhang, A.; Guo, C.; Cai, X.H.; et al. Structural basis of sphingosine-1-phosphate receptor 1 activation and biased agonism. Nat. Chem. Biol. 2022, 18, 281–288. [Google Scholar] [CrossRef]
- Liu, S.; Paknejad, N.; Zhu, L.; Kihara, Y.; Ray, M.; Chun, J.; Liu, W.; Hite, R.K.; Huang, X.Y. Differential activation mechanisms of lipid gpcrs by lysophosphatidic acid and sphingosine 1-phosphate. Nat. Commun. 2022, 13, 731. [Google Scholar] [CrossRef]
- Chen, H.; Chen, K.; Huang, W.; Staudt, L.M.; Cyster, J.G.; Li, X. Structure of s1pr2–heterotrimeric g13 complex. Sci. Adv. 2022, 8, eabn0067. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Shiimura, Y.; Asada, H.; Hirata, K.; Luo, F.; Nango, E.; Tanaka, N.; Toyomoto, M.; Inoue, A.; Aoki, J.; et al. Endogenous agonist–bound s1pr3 structure reveals determinants of g protein–subtype bias. Sci. Adv. 2021, 7, eabf5325. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Jia, G.; Wu, C.; Wang, W.; Cheng, L.; Li, Q.; Li, Z.; Luo, K.; Yang, S.; Yan, W.; et al. Structures of signaling complexes of lipid receptors s1pr1 and s1pr5 reveal mechanisms of activation and drug recognition. Cell Res. 2021, 31, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Lyapina, E.; Marin, E.; Gusach, A.; Orekhov, P.; Gerasimov, A.; Luginina, A.; Vakhrameev, D.; Ergasheva, M.; Kovaleva, M.; Khusainov, G.; et al. Structural basis for receptor selectivity and inverse agonism in s1p5 receptors. Nat. Commun. 2022, 13, 4736. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Shen, J.; Jiang, W.; Yang, Y.; Liu, X.; Zeng, Y. Sphingosine 1-phosphate and its regulatory role in vascular endothelial cells. Histol. Histopathol. 2022, 37, 213–225. [Google Scholar]
- Anwar, M.; Mehta, D. Post-translational modifications of s1pr1 and endothelial barrier regulation. Biochim. Biophys. Acta (BBA)-Mol. Cell. Biol. Lipids 2020, 1865, 158760. [Google Scholar] [CrossRef]
- Du, J.; Zeng, C.; Li, Q.; Chen, B.; Liu, H.; Huang, X.; Huang, Q. Lps and tnf-alpha induce expression of sphingosine-1-phosphate receptor-2 in human microvascular endothelial cells. Pathol. Res. Pract. 2012, 208, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Akhter, M.Z.; Chandra Joshi, J.; Balaji Ragunathrao, V.A.; Maienschein-Cline, M.; Proia, R.L.; Malik, A.B.; Mehta, D. Programming to s1pr1(+) endothelial cells promotes restoration of vascular integrity. Circ. Res. 2021, 129, 221–236. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.S.; Nierhaus, A.; Poppe, A.; Greiwe, G.; Gräler, M.H.; Daum, G. Sphingosine-1-phosphate: A potential biomarker and therapeutic target for endothelial dysfunction and sepsis? Shock 2017, 47, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Peng, H.; Chen, S.; Liu, Y.; Xiang, H.; Chen, R.; Chen, W.; Zhao, S.; Chen, P.; Lu, H. S1pr2 antagonist protects endothelial cells against high glucose-induced mitochondrial apoptosis through the akt/gsk-3β signaling pathway. Biochem. Biophys. Res. Commun. 2017, 490, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Singleton, P.A.; Letsiou, E.; Zhao, J.; Belvitch, P.; Sammani, S.; Chiang, E.T.; Moreno-Vinasco, L.; Wade, M.S.; Zhou, T.; et al. Sphingosine-1-phosphate receptor-3 is a novel biomarker in acute lung injury. Am. J. Respir. Cell Mol. Biol. 2012, 47, 628–636. [Google Scholar] [CrossRef]
- Zeng, Y.; Adamson, R.H.; Curry, F.R.; Tarbell, J.M. Sphingosine-1-phosphate protects endothelial glycocalyx by inhibiting syndecan-1 shedding. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H363–H372. [Google Scholar] [CrossRef]
- Zeng, Y.; Liu, X.H.; Tarbell, J.; Fu, B. Sphingosine 1-phosphate induced synthesis of glycocalyx on endothelial cells. Exp. Cell Res. 2015, 339, 90–95. [Google Scholar] [CrossRef]
- Zhang, L.; Zeng, M.; Fan, J.; Tarbell, J.M.; Curry, F.R.; Fu, B.M. Sphingosine-1-phosphate maintains normal vascular permeability by preserving endothelial surface glycocalyx in intact microvessels. Microcirculation 2016, 23, 301–310. [Google Scholar] [CrossRef]
- Mensah, S.A.; Cheng, M.J.; Homayoni, H.; Plouffe, B.D.; Coury, A.J.; Ebong, E.E. Regeneration of glycocalyx by heparan sulfate and sphingosine 1-phosphate restores inter-endothelial communication. PLoS ONE 2017, 12, e0186116. [Google Scholar] [CrossRef]
- Alves, N.G.; Trujillo, A.N.; Breslin, J.W.; Yuan, S.Y. Sphingosine-1-phosphate reduces hemorrhagic shock and resuscitation-induced microvascular leakage by protecting endothelial mitochondrial integrity. Shock 2019, 52, 423–433. [Google Scholar] [CrossRef]
- Diebel, M.E.; Diebel, L.N.; Liberati, D.M. Protective effects of plasma products on the endothelial-glycocalyx barrier following trauma-hemorrhagic shock: Is sphingosine-1 phosphate responsible? J. Trauma. Acute Care Surg. 2019, 87, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Gaengel, K.; Niaudet, C.; Hagikura, K.; Lavina, B.; Muhl, L.; Hofmann, J.J.; Ebarasi, L.; Nystrom, S.; Rymo, S.; Chen, L.L.; et al. The sphingosine-1-phosphate receptor s1pr1 restricts sprouting angiogenesis by regulating the interplay between ve-cadherin and vegfr2. Dev. Cell 2012, 23, 587–599. [Google Scholar] [CrossRef] [PubMed]
- Balaji Ragunathrao, V.A.; Anwar, M.; Akhter, M.Z.; Chavez, A.; Mao, Y.; Natarajan, V.; Lakshmikanthan, S.; Chrzanowska-Wodnicka, M.; Dudek, A.Z.; Claesson-Welsh, L.; et al. Sphingosine-1-phosphate receptor 1 activity promotes tumor growth by amplifying vegf-vegfr2 angiogenic signaling. Cell Rep. 2019, 29, 3472–3487. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.; Hagemann, N.; Sun, L.; Wu, J.; Doeppner, T.R.; Dai, Y.; Hermann, D.M. High-density lipoprotein (hdl) promotes angiogenesis via s1p3-dependent vegfr2 activation. Angiogenesis 2018, 21, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zheng, Y.; Wang, F.; Zhong, J.; Zhao, T.; Xie, Q.; Zhu, T.; Ma, F.; Tang, Q.; Zhou, B.; et al. Mfsd2a and spns2 are essential for sphingosine-1-phosphate transport in the formation and maintenance of the blood-brain barrier. Sci. Adv. 2020, 6, eaay8627. [Google Scholar] [CrossRef] [PubMed]
- Jeya Paul, J.; Weigel, C.; Müller, T.; Heller, R.; Spiegel, S.; Gräler, M.H. Inflammatory conditions disrupt constitutive endothelial cell barrier stabilization by alleviating autonomous secretion of sphingosine 1-phosphate. Cells 2020, 9, 928. [Google Scholar] [CrossRef]
- Igarashi, J.; Michel, T. Sphingosine-1-phosphate and modulation of vascular tone. Cardiovasc. Res. 2009, 82, 212–220. [Google Scholar] [CrossRef]
- Lee, D.Y.; Won, K.J.; Lee, K.P.; Jung, S.H.; Baek, S.; Chung, H.W.; Choi, W.S.; Lee, H.M.; Lee, B.H.; Jeon, B.H.; et al. Angiotensin ii facilitates neointimal formation by increasing vascular smooth muscle cell migration: Involvement of ape/ref-1-mediated overexpression of sphingosine-1-phosphate receptor 1. Toxicol. Appl. Pharmacol. 2018, 347, 45–53. [Google Scholar] [CrossRef]
- Kerage, D.; Gombos, R.B.; Wang, S.; Brown, M.; Hemmings, D.G. Sphingosine 1-phosphate-induced nitric oxide production simultaneously controls endothelial barrier function and vascular tone in resistance arteries. Vasc. Pharmacol. 2021, 140, 106874. [Google Scholar] [CrossRef]
- Jozefczuk, E.; Guzik, T.J.; Siedlinski, M. Significance of sphingosine-1-phosphate in cardiovascular physiology and pathology. Pharmacol. Res. 2020, 156, 104793. [Google Scholar] [CrossRef]
- Mascall, K.S.; Small, G.R.; Gibson, G.; Nixon, G.F. Sphingosine-1-phosphate-induced release of timp-2 from vascular smooth muscle cells inhibits angiogenesis. J. Cell Sci. 2012, 125, 2267–2275. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Xia, M.; Wang, Z.; Li, P.L.; Li, N. A novel lipid natriuretic factor in the renal medulla: Sphingosine-1-phosphate. Am. J. Physiol. Ren. Physiol. 2011, 301, F35–F41. [Google Scholar] [CrossRef] [PubMed]
- Huwiler, A.; Zangemeister-Wittke, U. The sphingosine 1-phosphate receptor modulator fingolimod as a therapeutic agent: Recent findings and new perspectives. Pharmacol. Ther. 2018, 185, 34–49. [Google Scholar] [CrossRef] [PubMed]
- Cantalupo, A.; Gargiulo, A.; Dautaj, E.; Liu, C.; Zhang, Y.; Hla, T.; Di Lorenzo, A. S1pr1 (sphingosine-1-phosphate receptor 1) signaling regulates blood flow and pressure. Hypertension 2017, 70, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Benechet, A.P.; Menon, M.; Xu, D.; Samji, T.; Maher, L.; Murooka, T.T.; Mempel, T.R.; Sheridan, B.S.; Lemoine, F.M.; Khanna, K.M. T cell-intrinsic s1pr1 regulates endogenous effector t-cell egress dynamics from lymph nodes during infection. Proc. Natl. Acad. Sci. USA 2016, 113, 2182–2187. [Google Scholar] [CrossRef]
- Means, C.K.; Brown, J.H. Sphingosine-1-phosphate receptor signalling in the heart. Cardiovasc. Res. 2009, 82, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Keul, P.; van Borren, M.M.; Ghanem, A.; Müller, F.U.; Baartscheer, A.; Verkerk, A.O.; Stümpel, F.; Schulte, J.S.; Hamdani, N.; Linke, W.A.; et al. Sphingosine-1-phosphate receptor 1 regulates cardiac function by modulating Ca2+ sensitivity and Na+/H+ exchange and mediates protection by ischemic preconditioning. J. Am. Heart Assoc. 2016, 5, e003393. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.; Lee, S.W.; Oh, J.; Lee, S.; Jang, I.J.; Yu, K.S. Dose-dependent reduction of lymphocyte count and heart rate after multiple administration of lc51-0255, a novel sphingosine-1-phosphate receptor 1 modulator, in healthy subjects. Front. Pharmacol. 2022, 13, 930615. [Google Scholar] [CrossRef]
- Constantinescu, V.; Haase, R.; Akgun, K.; Ziemssen, T. S1p receptor modulators and the cardiovascular autonomic nervous system in multiple sclerosis: A narrative review. Ther. Adv. Neurol. Disord. 2022, 15, 17562864221133163. [Google Scholar] [CrossRef]
- Guo, J.; MacDonell, K.L.; Giles, W.R. Effects of sphingosine 1-phosphate on pacemaker activity in rabbit sino-atrial node cells. Pflug. Arch. 1999, 438, 642–648. [Google Scholar] [CrossRef]
- Potì, F.; Simoni, M.; Nofer, J.R. Atheroprotective role of high-density lipoprotein (hdl)-associated sphingosine-1-phosphate (s1p). Cardiovasc. Res. 2014, 103, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Keul, P.; Polzin, A.; Kaiser, K.; Gräler, M.; Dannenberg, L.; Daum, G.; Heusch, G.; Levkau, B. Potent anti-inflammatory properties of hdl in vascular smooth muscle cells mediated by hdl-s1p and their impairment in coronary artery disease due to lower hdl-s1p: A new aspect of hdl dysfunction and its therapy. FASEB J. 2019, 33, 1482–1495. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, L.; Qian, A.S.; Tahir, U.; Yu, P.; Trigatti, B.L. Sphingosine-1-phosphate receptor 1, expressed in myeloid cells, slows diet-induced atherosclerosis and protects against macrophage apoptosis in ldlr ko mice. Int. J. Mol. Sci. 2017, 18, 2721. [Google Scholar] [CrossRef]
- Potì, F.; Ceglarek, U.; Burkhardt, R.; Simoni, M.; Nofer, J.R. Ski-II—A sphingosine kinase 1 inhibitor—Exacerbates atherosclerosis in low-density lipoprotein receptor-deficient (ldl-r−/−) mice on high cholesterol diet. Atherosclerosis 2015, 240, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Bot, M.; Van Veldhoven, P.P.; de Jager, S.C.; Johnson, J.; Nijstad, N.; Van Santbrink, P.J.; Westra, M.M.; Van Der Hoeven, G.; Gijbels, M.J.; Müller-Tidow, C.; et al. Hematopoietic sphingosine 1-phosphate lyase deficiency decreases atherosclerotic lesion development in ldl-receptor deficient mice. PLoS ONE 2013, 8, e63360. [Google Scholar] [CrossRef]
- Sattler, K.J.; Elbasan, S.; Keul, P.; Elter-Schulz, M.; Bode, C.; Gräler, M.H.; Bröcker-Preuss, M.; Budde, T.; Erbel, R.; Heusch, G.; et al. Sphingosine 1-phosphate levels in plasma and hdl are altered in coronary artery disease. Basic Res. Cardiol. 2010, 105, 821–832. [Google Scholar] [CrossRef]
- Sattler, K.; Gräler, M.; Keul, P.; Weske, S.; Reimann, C.M.; Jindrová, H.; Kleinbongard, P.; Sabbadini, R.; Bröcker-Preuss, M.; Erbel, R.; et al. Defects of high-density lipoproteins in coronary artery disease caused by low sphingosine-1-phosphate content: Correction by sphingosine-1-phosphate-loading. J. Am. Coll. Cardiol. 2015, 66, 1470–1485. [Google Scholar] [CrossRef]
- Imeri, F.; Blanchard, O.; Jenni, A.; Schwalm, S.; Wünsche, C.; Zivkovic, A.; Stark, H.; Pfeilschifter, J.; Huwiler, A. Fty720 and two novel butterfly derivatives exert a general anti-inflammatory potential by reducing immune cell adhesion to endothelial cells through activation of s1p3 and phosphoinositide 3-kinase. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 388, 1283–1292. [Google Scholar] [CrossRef]
- Kurano, M.; Yatomi, Y. Sphingosine 1-phosphate and atherosclerosis. J. Atheroscler. Thromb. 2018, 25, 16–26. [Google Scholar] [CrossRef]
- Therond, P.; Chapman, M.J. Sphingosine-1-phosphate: Metabolism, transport, atheroprotection and effect of statin treatment. Curr. Opin. Lipidol. 2022, 33, 199–207. [Google Scholar] [CrossRef]
- Ma, Q.; Reiter, R.J.; Chen, Y. Role of melatonin in controlling angiogenesis under physiological and pathological conditions. Angiogenesis 2020, 23, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qiu, Z.; Dong, W.; Yang, Z.; Wang, J.; Xu, H.; Sun, T.; Huang, Z.; Jin, J. S1pr1 induces metabolic reprogramming of ceramide in vascular endothelial cells, affecting hepatocellular carcinoma angiogenesis and progression. Cell Death Dis. 2022, 13, 768. [Google Scholar] [CrossRef] [PubMed]
- LaMontagne, K.; Littlewood-Evans, A.; Schnell, C.; O’Reilly, T.; Wyder, L.; Sanchez, T.; Probst, B.; Butler, J.; Wood, A.; Liau, G.; et al. Antagonism of sphingosine-1-phosphate receptors by fty720 inhibits angiogenesis and tumor vascularization. Cancer Res. 2006, 66, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Kuang, Y.; Li, Q.; Duan, Y.; Liu, X.; Yue, J.; Chen, X.; Liu, J.; Zhang, Y.; Zhang, L. Endothelial s1pr2 regulates post-ischemic angiogenesis via akt/enos signaling pathway. Theranostics 2022, 12, 5172–5188. [Google Scholar] [CrossRef]
- Wilkerson, J.L.; Basu, S.K.; Mandal, N.A. Angiogenesis model of cornea to understand the role of sphingosine 1-phosphate. Methods Mol. Biol. 2023, 2625, 231–240. [Google Scholar]
- Wilson, P.C.; Fitzgibbon, W.R.; Garrett, S.M.; Jaffa, A.A.; Luttrell, L.M.; Brands, M.W.; El-Shewy, H.M. Inhibition of sphingosine kinase 1 ameliorates angiotensin ii-induced hypertension and inhibits transmembrane calcium entry via store-operated calcium channel. Mol. Endocrinol. 2015, 29, 896–908. [Google Scholar] [CrossRef]
- Li, J.J.; Zhao, X.Y.; Wang, Y.; Xu, R.; Di, X.H.; Zhang, Y.; Yang, H.; Han, M.Z.; Bai, R.Y.; Xie, L.; et al. Endothelial k(ca)3.1 and k(ca)2.3 mediate s1p (sphingosine-1-phosphate)-dependent vasodilation and blood pressure homeostasis. Arter. Thromb. Vasc. Biol. 2023, 43, 726–738. [Google Scholar] [CrossRef]
- Werth, S.; Müller-Fielitz, H.; Raasch, W. Obesity-stimulated aldosterone release is not related to an s1p-dependent mechanism. J. Endocrinol. 2017, 235, 251–265. [Google Scholar] [CrossRef]
- Swendeman, S.L.; Xiong, Y.; Cantalupo, A.; Yuan, H.; Burg, N.; Hisano, Y.; Cartier, A.; Liu, C.H.; Engelbrecht, E.; Blaho, V.; et al. An engineered s1p chaperone attenuates hypertension and ischemic injury. Sci. Signal. 2017, 10, eaal2722. [Google Scholar] [CrossRef]
- Kerage, D.; Brindley, D.N.; Hemmings, D.G. Review: Novel insights into the regulation of vascular tone by sphingosine 1-phosphate. Placenta 2014, 35, S86–S92. [Google Scholar] [CrossRef]
- Kennedy, S.; Kane, K.A.; Pyne, N.J.; Pyne, S. Targeting sphingosine-1-phosphate signalling for cardioprotection. Curr. Opin. Pharmacol. 2009, 9, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Theilmeier, G.; Schmidt, C.; Herrmann, J.; Keul, P.; Schafers, M.; Herrgott, I.; Mersmann, J.; Larmann, J.; Hermann, S.; Stypmann, J.; et al. High-density lipoproteins and their constituent, sphingosine-1-phosphate, directly protect the heart against ischemia/reperfusion injury in vivo via the s1p3 lysophospholipid receptor. Circulation 2006, 114, 1403–1409. [Google Scholar] [CrossRef] [PubMed]
- Nitzsche, A.; Poittevin, M.; Benarab, A.; Bonnin, P.; Faraco, G.; Uchida, H.; Favre, J.; Garcia-Bonilla, L.; Garcia, M.C.L.; Leger, P.L.; et al. Endothelial s1p1 signaling counteracts infarct expansion in ischemic stroke. Circ. Res. 2021, 128, 363–382. [Google Scholar] [CrossRef]
- Yung, B.S.; Brand, C.S.; Xiang, S.Y.; Gray, C.B.; Means, C.K.; Rosen, H.; Chun, J.; Purcell, N.H.; Brown, J.H.; Miyamoto, S. Selective coupling of the s1p3 receptor subtype to s1p-mediated rhoa activation and cardioprotection. J. Mol. Cell. Cardiol. 2017, 103, 1–10. [Google Scholar] [CrossRef]
- Deshpande, G.P.; Imamdin, A.; Lecour, S.; Opie, L.H. Sphingosine-1-phosphate (s1p) activates stat3 to protect against de novo acute heart failure (ahf). Life Sci. 2018, 196, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xu, J.; Li, X.; Dai, J.; Wang, L. Inhibition of sphk1/s1p signaling pathway alleviates fibrosis and inflammation of rat myocardium after myocardial infarction. Comput. Math. Methods Med. 2022, 2022, 5985375. [Google Scholar] [CrossRef]
- Gupta, S.; Ge, Y.; Singh, A.; Gräni, C.; Kwong, R.Y. Multimodality imaging assessment of myocardial fibrosis. JACC Cardiovasc. Imaging 2021, 14, 2457–2469. [Google Scholar] [CrossRef]
- Cannavo, A.; Rengo, G.; Liccardo, D.; Pagano, G.; Zincarelli, C.; De Angelis, M.C.; Puglia, R.; Di Pietro, E.; Rabinowitz, J.E.; Barone, M.V.; et al. Β1-adrenergic receptor and sphingosine-1-phosphate receptor 1 (s1pr1) reciprocal downregulation influences cardiac hypertrophic response and progression to heart failure: Protective role of s1pr1 cardiac gene therapy. Circulation 2013, 128, 1612–1622. [Google Scholar] [CrossRef]
- Ohkura, S.I.; Usui, S.; Takashima, S.I.; Takuwa, N.; Yoshioka, K.; Okamoto, Y.; Inagaki, Y.; Sugimoto, N.; Kitano, T.; Takamura, M.; et al. Augmented sphingosine 1 phosphate receptor-1 signaling in cardiac fibroblasts induces cardiac hypertrophy and fibrosis through angiotensin ii and interleukin-6. PLoS ONE 2017, 12, e0182329. [Google Scholar] [CrossRef]
- Chen, R.; Cai, X.; Liu, J.; Bai, B.; Li, X. Sphingosine 1-phosphate promotes mesenchymal stem cell-mediated cardioprotection against myocardial infarction via erk1/2-mmp-9 and akt signaling axis. Life Sci. 2018, 215, 31–42. [Google Scholar] [CrossRef]
- Deng, S.; Zhou, X.; Ge, Z.; Song, Y.; Wang, H.; Liu, X.; Zhang, D. Exosomes from adipose-derived mesenchymal stem cells ameliorate cardiac damage after myocardial infarction by activating s1p/sk1/s1pr1 signaling and promoting macrophage m2 polarization. Int. J. Biochem. Cell. Biol. 2019, 114, 105564. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, J.; Shu, Z.; Chen, S.; Xiang, H.; Lu, H. The role of sphingosine 1-phosphate and its receptors in cardiovascular diseases. J. Cell. Mol. Med. 2020, 24, 10290–10301. [Google Scholar] [CrossRef] [PubMed]
- Gellings Lowe, N.; Swaney, J.S.; Moreno, K.M.; Sabbadini, R.A. Sphingosine-1-phosphate and sphingosine kinase are critical for transforming growth factor-beta-stimulated collagen production by cardiac fibroblasts. Cardiovasc. Res. 2009, 82, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Visentin, B.; Vekich, J.A.; Sibbald, B.J.; Cavalli, A.L.; Moreno, K.M.; Matteo, R.G.; Garland, W.A.; Lu, Y.; Yu, S.; Hall, H.S.; et al. Validation of an anti-sphingosine-1-phosphate antibody as a potential therapeutic in reducing growth, invasion, and angiogenesis in multiple tumor lineages. Cancer Cell 2006, 9, 225–238. [Google Scholar] [CrossRef]
- Sah, V.P.; Seasholtz, T.M.; Sagi, S.A.; Brown, J.H. The role of rho in g protein-coupled receptor signal transduction. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 459–489. [Google Scholar] [CrossRef]
- Hattori, T.; Shimokawa, H.; Higashi, M.; Hiroki, J.; Mukai, Y.; Tsutsui, H.; Kaibuchi, K.; Takeshita, A. Long-term inhibition of rho-kinase suppresses left ventricular remodeling after myocardial infarction in mice. Circulation 2004, 109, 2234–2239. [Google Scholar] [CrossRef]
- Rikitake, Y.; Oyama, N.; Wang, C.Y.; Noma, K.; Satoh, M.; Kim, H.H.; Liao, J.K. Decreased perivascular fibrosis but not cardiac hypertrophy in rock1+/− haploinsufficient mice. Circulation 2005, 112, 2959–2965. [Google Scholar] [CrossRef]
- Lijnen, P.; Petrov, V. Induction of cardiac fibrosis by aldosterone. J. Mol. Cell. Cardiol. 2000, 32, 865–879. [Google Scholar] [CrossRef]
- Yang, C.C.; Hsiao, L.D.; Shih, Y.F.; Su, M.H.; Yang, C.M. Sphingosine 1-phosphate-upregulated cox-2/pge(2) system contributes to human cardiac fibroblast apoptosis: Involvement of mmp-9-dependent transactivation of egfr cascade. Oxidative Med. Cell. Longev. 2022, 2022, 7664290. [Google Scholar]
- Kuang, Y.; Li, X.; Liu, X.; Wei, L.; Chen, X.; Liu, J.; Zhuang, T.; Pi, J.; Wang, Y.; Zhu, C.; et al. Vascular endothelial s1pr1 ameliorates adverse cardiac remodelling via stimulating reparative macrophage proliferation after myocardial infarction. Cardiovasc. Res. 2021, 117, 585–599. [Google Scholar] [CrossRef]
- Li, Q.; Zhou, C.; Zhao, K.; Duan, Y.; Yue, J.; Liu, X.; Wu, J.; Deng, S. Lymphatic endothelial sphingosine 1-phosphate receptor 1 enhances macrophage clearance via lymphatic system following myocardial infarction. Front. Cardiovasc. Med. 2022, 9, 872102. [Google Scholar] [CrossRef] [PubMed]
- Takuwa, N.; Ohkura, S.; Takashima, S.; Ohtani, K.; Okamoto, Y.; Tanaka, T.; Hirano, K.; Usui, S.; Wang, F.; Du, W.; et al. S1p3-mediated cardiac fibrosis in sphingosine kinase 1 transgenic mice involves reactive oxygen species. Cardiovasc. Res. 2010, 85, 484–493. [Google Scholar] [CrossRef] [PubMed]
- Keller, C.D.; Rivera Gil, P.; Tolle, M.; van der Giet, M.; Chun, J.; Radeke, H.H.; Schafer-Korting, M.; Kleuser, B. Immunomodulator fty720 induces myofibroblast differentiation via the lysophospholipid receptor s1p3 and smad3 signaling. Am. J. Pathol. 2007, 170, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Polzin, A.; Piayda, K.; Keul, P.; Dannenberg, L.; Mohring, A.; Gräler, M.; Zeus, T.; Kelm, M.; Levkau, B. Plasma sphingosine-1-phosphate concentrations are associated with systolic heart failure in patients with ischemic heart disease. J. Mol. Cell. Cardiol. 2017, 110, 35–37. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cell Type | Function | Role of S1PRs | Associated Diseases |
|---|---|---|---|
| Endothelial cell | Barrier maintenance, angiogenesis, lymphocyte trafficking, vascular tone, and inflammation | S1PR1: protect or rescue endothelial barrier function; promote endothelium-dependent vasorelaxation | Atherosclerosis, angiogenesis, and hypertension |
| S1PR2: disrupt adhesion junctions; increase paracellular permeability | |||
| S1PR3: mediate cell proliferation and vascular permeability; a potential biomarker in acute lung injury (ALI); promote endothelium-dependent vasorelaxation | |||
| Vascular smooth muscle cell | Angiogenesis, inflammation, and vascular tone | S1PR1-3: regulate VSMC proliferation, migration and VSMC-dependent vascular tone | |
| S1PR2/3: mediate vasoconstriction and anti-inflammation | |||
| Renal medullary epithelial cell | Water and sodium homeostasis | S1PR1: increase urine flow and sodium excretion; regulate blood pressure through effects on blood volume | |
| Cardiomyocyte | Affect cardiac muscle contraction, cardiomyocyte viability, and heart rhythm | S1PR1: produce the negative inotropic effects by inhibiting L-type calcium channel current, activating inwardly rectifying K+ current and shortening action potential duration (APD) | Myocardial ischemia and reinfusion (I/R) injury, cardiac hypertrophy, myocardial infarction, fibrosis and heart failure |
| S1PR2/3: lead to negative inotropic response by inhibiting isoproterenol-stimulated cAMP accumulation | |||
| Fibroblast | Regulation of cardiac fibroblast proliferation, migration, differentiation, and survival | S1PR1: regulate of cardiac fibrosis | |
| S1PR2: mediate fibroblast transformation and collagen production | |||
| S1PR3: mediate myofibroblast differentiation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, N.; Li, J.-Y.; Zeng, B.; Chen, G.-L. Sphingosine-1-Phosphate Signaling in Cardiovascular Diseases. Biomolecules 2023, 13, 818. https://doi.org/10.3390/biom13050818
Wang N, Li J-Y, Zeng B, Chen G-L. Sphingosine-1-Phosphate Signaling in Cardiovascular Diseases. Biomolecules. 2023; 13(5):818. https://doi.org/10.3390/biom13050818
Chicago/Turabian StyleWang, Na, Jing-Yi Li, Bo Zeng, and Gui-Lan Chen. 2023. "Sphingosine-1-Phosphate Signaling in Cardiovascular Diseases" Biomolecules 13, no. 5: 818. https://doi.org/10.3390/biom13050818
APA StyleWang, N., Li, J. -Y., Zeng, B., & Chen, G. -L. (2023). Sphingosine-1-Phosphate Signaling in Cardiovascular Diseases. Biomolecules, 13(5), 818. https://doi.org/10.3390/biom13050818