
Serum N-Glycosylation RPLC-FD-MS Assay to Assess Colorectal Cancer Surgical Interventions

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. N-Glycan Release and Fluorescent Labeling
2.3. Sialic Acid Derivatization
2.4. Reversed-Phase Liquid Chromatography–Fluorescence Detection–Mass Spectrometry
2.5. Data Processing
2.6. Statistical Analysis
3. Results & Discussion
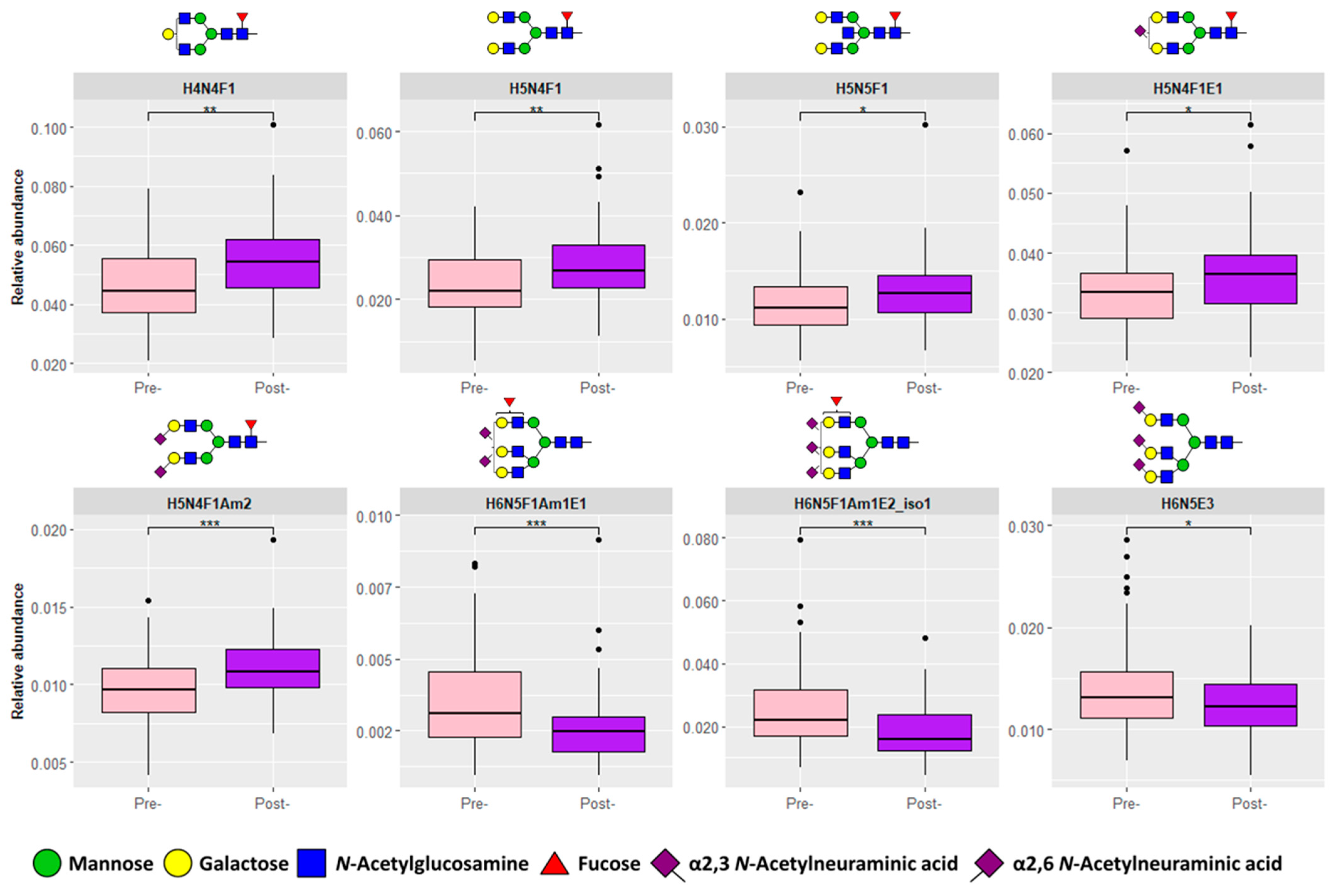
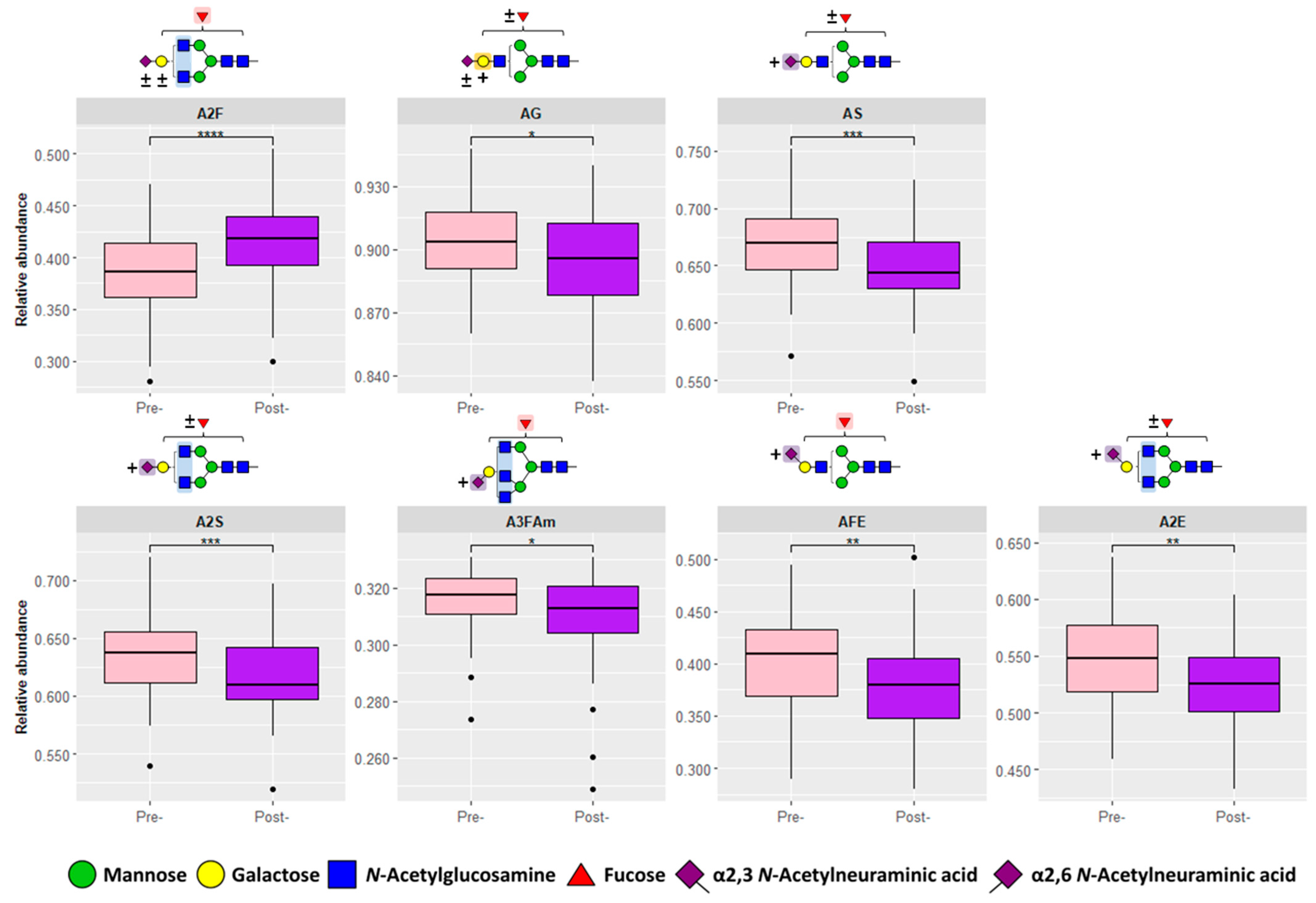
3.1. Serum N-Glycosylation Analysis of Pre- and Post-Operative CRC Samples
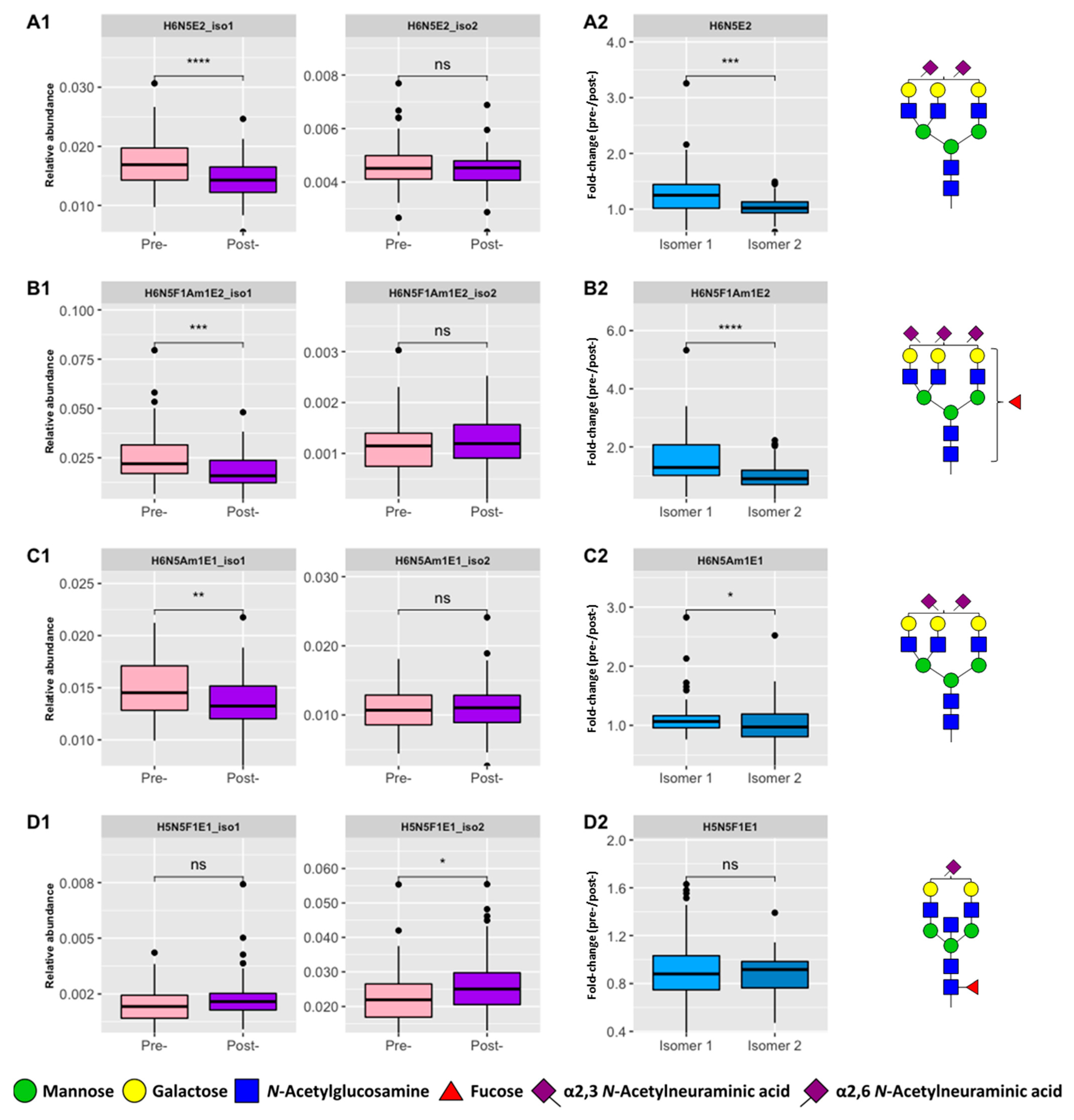
3.2. Positional Isomers in CRC N-Glycome
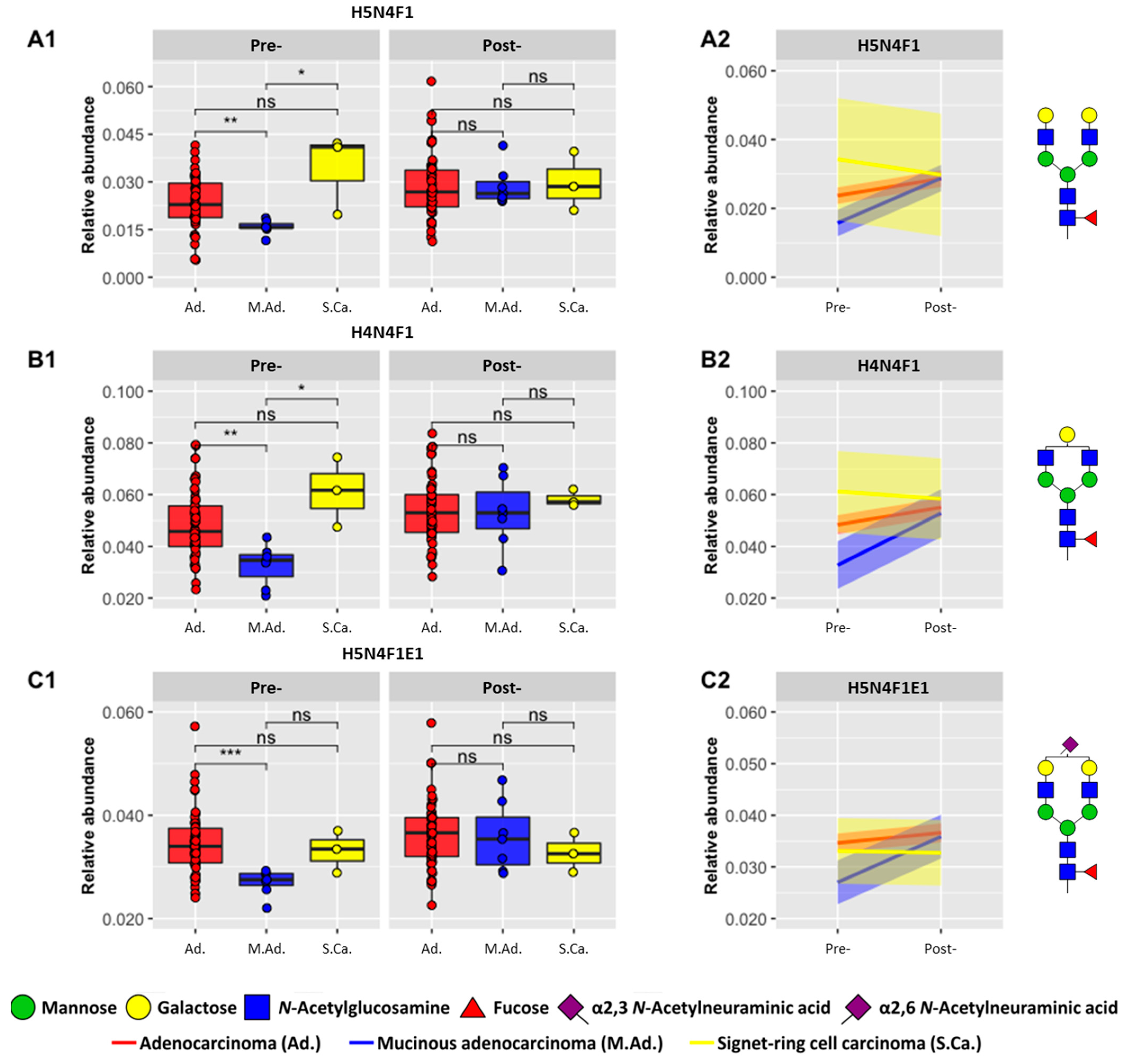
3.3. Investigation of Pre- and Post-Operative Histological Type
3.4. Comparison of Clinical Performance of Three Quantification Approaches
4. Perspectives
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vajaria, B.N.; Patel, P. Glycosylation: A hallmark of cancer? Glycoconj. J. 2017, 34, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Silsirivanit, A. Glycosylation Markers in Cancer. Adv. Clin. Chem. 2019, 89, 189–213. [Google Scholar]
- Ohtsubo, K.; Marth, J.D. Glycosylation in Cellular Mechanisms of Health and Disease. Cell 2006, 126, 855–867. [Google Scholar] [CrossRef]
- Munkley, J.; Elliott, D.J. Hallmarks of glycosylation in cancer. Oncotarget 2016, 7, 35478–35489. [Google Scholar] [CrossRef]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef]
- Boyaval, F.; Dalebout, H.; Van Zeijl, R.; Wang, W.; Fariña-Sarasqueta, A.; Lageveen-Kammeijer, G.S.M.; Boonstra, J.J.; McDonnell, L.A.; Wuhrer, M.; Morreau, H.; et al. High-Mannose N-Glycans as Malignant Progression Markers in Early-Stage Colorectal Cancer. Cancers 2022, 14, 1552. [Google Scholar] [CrossRef] [PubMed]
- Boyaval, F.; van Zeijl, R.; Dalebout, H.; Holst, S.; van Pelt, G.W.; Fariña-Sarasqueta, A.; Mesker, W.E.; Tollenaar, R.; Morreau, H.; Wuhrer, M.; et al. N-Glycomic Signature of Stage II Colorectal Cancer and Its Association with the Tumor Microenvironment. Mol. Cell. Proteom. 2021, 20, 100057. [Google Scholar] [CrossRef] [PubMed]
- Drake, R.R.; Powers, T.W.; Jones, E.E.; Bruner, E.; Mehta, A.S.; Angel, P.M. MALDI Mass Spectrometry Imaging of N-Linked Glycans in Cancer Tissues. Adv. Cancer Res. 2017, 134, 85–116. [Google Scholar] [CrossRef]
- Houvast, R.D.; Vankemmelbeke, M.; Durrant, L.G.; Wuhrer, M.; Baart, V.M.; Kuppen, P.J.K.; de Geus-Oei, L.-F.; Vahrmeijer, A.L.; Sier, C.F.M. Targeting Glycans and Heavily Glycosylated Proteins for Tumor Imaging. Cancers 2020, 12, 3870. [Google Scholar] [CrossRef]
- Holm, M.; Nummela, P.; Heiskanen, A.; Satomaa, T.; Kaprio, T.; Mustonen, H.K.; Ristimäki, A.; Haglund, C. N-glycomic profiling of colorectal cancer according to tumor stage and location. PLoS ONE 2020, 15, e0234989. [Google Scholar] [CrossRef]
- Pan, Y.; Zhang, L.; Zhang, R.; Han, J.; Qin, W.; Gu, Y.; Sha, J.; Xu, X.; Feng, Y.; Ren, Z.; et al. Screening and diagnosis of colorectal cancer and advanced adenoma by Bionic Glycome method and machine learning. Am. J. Cancer Res. 2021, 11, 3002–3020. [Google Scholar]
- Bertok, T.; Bertokova, A.; Jane, E.; Hires, M.; Aguedo, J.; Potocarova, M.; Lukac, L.; Vikartovska, A.; Kasak, P.; Borsig, L.; et al. Identification of Whole-Serum Glycobiomarkers for Colorectal Carcinoma Using Reverse-Phase Lectin Microarray. Front. Oncol. 2021, 11, 735338. [Google Scholar] [CrossRef]
- Crosby, D.; Lyons, N.; Greenwood, E.; Harrison, S.; Hiom, S.; Moffat, J.; Quallo, T.; Samuel, E.; Walker, I. A roadmap for the early detection and diagnosis of cancer. Lancet Oncol. 2020, 21, 1397–1399. [Google Scholar] [CrossRef] [PubMed]
- de Vroome, S.W.; Holst, S.; Girondo, M.R.; van der Burgt, Y.E.; Mesker, W.E.; Tollenaar, R.A.; Wuhrer, M. Serum N-glycome alterations in colorectal cancer associate with survival. Oncotarget 2018, 9, 30610–30623. [Google Scholar] [CrossRef]
- Vreeker, G.C.M.; Nicolardi, S.; Bladergroen, M.R.; van der Plas, C.J.; Mesker, W.E.; Tollenaar, R.A.E.M.; van der Burgt, Y.E.M.; Wuhrer, M. Automated Plasma Glycomics with Linkage-Specific Sialic Acid Esterification and Ultrahigh Resolution MS. Anal. Chem. 2018, 90, 11955–11961. [Google Scholar] [CrossRef] [PubMed]
- Moran, A.B.; Gardner, R.A.; Wuhrer, M.; Lageveen-Kammeijer, G.S.M.; Spencer, D.I.R. Sialic Acid Derivatization of Fluorescently Labeled N-Glycans Allows Linkage Differentiation by Reversed-Phase Liquid Chromatography–Fluorescence Detection–Mass Spectrometry. Anal. Chem. 2022, 94, 6639–6648. [Google Scholar] [CrossRef] [PubMed]
- Reiding, K.R.; Blank, D.; Kuijper, D.M.; Deelder, A.M.; Wuhrer, M. High-Throughput Profiling of Protein N-Glycosylation by MALDI-TOF-MS Employing Linkage-Specific Sialic Acid Esterification. Anal. Chem. 2014, 86, 5784–5793. [Google Scholar] [CrossRef]
- Lageveen-Kammeijer, G.S.M.; de Haan, N.; Mohaupt, P.; Wagt, S.; Filius, M.; Nouta, J.; Falck, D.; Wuhrer, M. Highly sensitive CE-ESI-MS analysis of N-glycans from complex biological samples. Nat. Commun. 2019, 10, 2137. [Google Scholar] [CrossRef]
- Varki, A.; Cummings, R.D.; Aebi, M.; Packer, N.H.; Seeberger, P.H.; Esko, J.D.; Stanley, P.; Hart, G.; Darvill, A.; Kinoshita, T.; et al. Symbol Nomenclature for Graphical Representations of Glycans. Glycobiology 2015, 25, 1323–1324. [Google Scholar] [CrossRef]
- Jansen, B.C.; Hafkenscheid, L.; Bondt, A.; Gardner, R.A.; Hendel, J.L.; Wuhrer, M.; Spencer, D.I.R. HappyTools: A software for high-throughput HPLC data processing and quantitation. PLoS ONE 2018, 13, e0200280. [Google Scholar] [CrossRef]
- European Medicines Agency (EMA). ICH Guidelines Q2(R2) on Validation of Analytical Procedures; European Medicines Agency (EMA): Amsterdam, The Netherlands, 2022; Volume 2. [Google Scholar]
- Jansen, B.C.; Falck, D.; de Haan, N.; Hipgrave Ederveen, A.L.; Razdorov, G.; Lauc, G.; Wuhrer, M. LaCyTools: A Targeted Liquid Chromatography–Mass Spectrometry Data Processing Package for Relative Quantitation of Glycopeptides. J. Proteome Res. 2016, 15, 2198–2210. [Google Scholar] [CrossRef]
- Varki, A.; Esko, J.D.; Colley, K.J. Cellular Organization of Glycosylation. In Essentials of Glycobiology, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2017. [Google Scholar]
- Wehrens, R.; Hageman, J.; Van Eeuwijk, F.; Kooke, R.; Flood, P.; Wijnker, E.; Keurentjes, J.; Lommen, A.; Van Eekelen, H.D.L.M.; Hall, R.; et al. Improved batch correction in untargeted MS-based metabolomics. Metabolomics 2016, 12, 88. [Google Scholar] [CrossRef]
- Takei, D.; Harada, K.; Nouso, K.; Miyahara, K.; Dohi, C.; Matsushita, H.; Kinugasa, H.; Hiraoka, S.; Nishimura, S.; Okada, H. Clinical utility of a serum glycome analysis in patients with colorectal cancer. J. Gastroenterol. Hepatol. 2022, 37, 727–733. [Google Scholar] [CrossRef]
- Zhao, Y.-P.; Ruan, C.-P.; Wang, H.; Hu, Z.-Q.; Fang, M.; Gu, X.; Ji, J.; Zhao, J.-Y.; Gao, C.-F. Identification and assessment of new biomarkers for colorectal cancer with serum N-glycan profiling. Cancer 2012, 118, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Doherty, M.; Theodoratou, E.; Walsh, I.; Adamczyk, B.; Stöckmann, H.; Agakov, F.; Timofeeva, M.; Trbojević-Akmačić, I.; Vučković, F.; Duffy, F.; et al. Plasma N-glycans in colorectal cancer risk. Sci. Rep. 2018, 8, 8655. [Google Scholar] [CrossRef]
- Balog, C.I.A.; Stavenhagen, K.; Fung, W.L.J.; Koeleman, C.A.; McDonnell, L.A.; Verhoeven, A.; Mesker, W.E.; Tollenaar, R.A.E.M.; Deelder, A.M.; Wuhrer, M. N-glycosylation of Colorectal Cancer Tissues: A liquid chromatography and mass spectrometry-based investigation. Mol. Cell. Proteom. 2012, 11, 571–585. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-J.; Lee, M. Increasing the α 2, 6 Sialylation of Glycoproteins May Contribute to Metastatic Spread and Therapeutic Resistance in Colorectal Cancer. Gut Liver 2013, 7, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Nakagoe, T.; Sawai, T.; Tsuji, T.; Jibiki, M.A.; Nanashima, A.; Yamaguchi, H.; Yasutake, T.; Kurosaki, N.; Ayabe, H.; Arisawa, K. Preoperative serum levels of sialyl Lewis(a), sialyl Lewis(x), and sialyl Tn antigens as prognostic markers after curative resection for colorectal cancer. Cancer Detect. Prev. 2001, 25, 299–308. [Google Scholar]
- Miyoshi, E.; Moriwaki, K.; Nakagawa, T. Biological Function of Fucosylation in Cancer Biology. J. Biochem. 2008, 143, 725–729. [Google Scholar] [CrossRef]
- Rebello, O.D.; Nicolardi, S.; Lageveen-Kammeijer, G.S.M.; Nouta, J.; Gardner, R.A.; Mesker, W.E.; Tollenaar, R.A.E.M.; Spencer, D.I.R.; Wuhrer, M.; Falck, D. A Matrix-Assisted Laser Desorption/Ionization—Mass Spectrometry Assay for the Relative Quantitation of Antennary Fucosylated N-Glycans in Human Plasma. Front. Chem. 2020, 8, 138. [Google Scholar] [CrossRef]
- de Hoffmann, E.; Stroobant, V. Mass Spectrometry: Principles and Applications, 3rd ed.; Wiley: Chichester, UK, 2007; ISBN 978-0-470-03310-4. [Google Scholar]
- Keser, T.; Pavić, T.; Lauc, G.; Gornik, O. Comparison of 2-Aminobenzamide, Procainamide and RapiFluor-MS as Derivatizing Agents for High-Throughput HILIC-UPLC-FLR-MS N-glycan Analysis. Front. Chem. 2018, 6, 324. [Google Scholar] [CrossRef] [PubMed]
- Fleming, M.; Ravula, S.; Tatishchev, S.F.; Wang, H.L. Colorectal carcinoma: Pathologic aspects. J. Gastrointest. Oncol. 2012, 3, 153–173. [Google Scholar] [CrossRef] [PubMed]
- Makino, T.; Tsujinaka, T.; Mishima, H.; Ikenaga, M.; Sawamura, T.; Nakamori, S.; Fujitani, K.; Hirao, M.; Kashiwazaki, M.; Masuda, N.; et al. Primary signet-ring cell carcinoma of the colon and rectum: Report of eight cases and review of 154 Japanese cases. Hepatogastroenterology 2006, 53, 845–849. [Google Scholar] [PubMed]
- Chen, J.S.; Hsieh, P.S.; Chiang, J.M.; Yeh, C.Y.; Tsai, W.S.; Tang, R.; Changchien, C.R.; Wu, R.C. Clinical outcome of signet ring cell carcinoma and mucinous adenocarcinoma of the colon. Chang Gung Med. J. 2010, 33, 51–57. [Google Scholar]
- Kang, H.; O'Connell, J.B.; Maggard, M.A.; Sack, J.; Ko, C.Y. A 10-Year Outcomes Evaluation of Mucinous and Signet-Ring Cell Carcinoma of the Colon and Rectum. Dis. Colon Rectum 2005, 48, 1161–1168. [Google Scholar] [CrossRef]
- Leopoldo, S.; Lorena, B.; Cinzia, A.; Gabriella, D.C.; Angela Luciana, B.; Renato, C.; Antonio, M.; Carlo, S.; Cristina, P.; Stefano, C.; et al. Two Subtypes of Mucinous Adenocarcinoma of The Colorectum: Clinicopathological and Genetic Features. Ann. Surg. Oncol. 2008, 15, 1429–1439. [Google Scholar] [CrossRef]
- Compton, C.C.; Fielding, L.P.; Burgart, L.J.; Conley, B.; Cooper, H.S.; Hamilton, S.R.; Hammond, M.E.; Henson, D.E.; Hutter, R.V.; Nagle, R.B.; et al. Prognostic factors in colorectal cancer. College of American Pathologists Consensus Statement 1999. Arch. Pathol. Lab. Med. 2000, 124, 979–994. [Google Scholar] [CrossRef]
- Hanna-Sawires, R.G.; Schiphuis, J.H.; Wuhrer, M.; Vasen, H.F.A.; van Leerdam, M.E.; Bonsing, B.A.; Mesker, W.E.; van der Burgt, Y.E.M.; Tollenaar, R.A.E.M. Clinical Perspective on Proteomic and Glycomic Biomarkers for Diagnosis, Prognosis, and Prediction of Pancreatic Cancer. Int. J. Mol. Sci. 2021, 22, 2655. [Google Scholar] [CrossRef]
- Jackson, S.E.; Chester, J.D. Personalised cancer medicine. Int. J. Cancer 2014, 137, 262–266. [Google Scholar] [CrossRef]
- Stewart, S.L.; Wike, J.M.; Kato, I.; Lewis, D.R.; Michaud, F. A population-based study of colorectal cancer histology in the United States, 1998–2001. Cancer 2006, 107, 1128–1141. [Google Scholar] [CrossRef]
- Mereiter, S.; Balmaña, M.; Campos, D.; Gomes, J.; Reis, C.A. Glycosylation in the Era of Cancer-Targeted Therapy: Where Are We Heading? Cancer Cell 2019, 36, 6–16. [Google Scholar] [CrossRef]
- Lau, K.S.; Partridge, E.A.; Grigorian, A.; Silvescu, C.I.; Reinhold, V.N.; Demetriou, M.; Dennis, J.W. Complex N-Glycan Number and Degree of Branching Cooperate to Regulate Cell Proliferation and Differentiation. Cell 2007, 129, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Hang, Q.; Isaji, T.; Hou, S.; Zhou, Y.; Fukuda, T.; Gu, J. N-Glycosylation of integrin α5 acts as a switch for EGFR-mediated complex formation of integrin α5β1 to α6β4. Sci. Rep. 2016, 6, 33507. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, T.; Ohyama, C.; Hatakeyama, S.; Narita, S.; Habuchi, T.; Koie, T.; Mori, K.; Hidari, K.I.P.J.; Yamaguchi, M.; Suzuki, T.; et al. Measurement of aberrant glycosylation of prostate specific antigen can improve specificity in early detection of prostate cancer. Biochem. Biophys. Res. Commun. 2014, 448, 390–396. [Google Scholar] [CrossRef] [PubMed]
- Vreeker, G.C.M.; Hanna-Sawires, R.G.; Mohammed, Y.; Bladergroen, M.R.; Nicolardi, S.; Dotz, V.; Nouta, J.; Bonsing, B.A.; Mesker, W.E.; van der Burgt, Y.E.M.; et al. Serum N-Glycome analysis reveals pancreatic cancer disease signatures. Cancer Med. 2020, 9, 8519–8529. [Google Scholar] [CrossRef]
- Almeida, A.; Kolarich, D. The promise of protein glycosylation for personalised medicine. Biochim. Biophys. Acta 2016, 1860, 1583–1595. [Google Scholar] [CrossRef]
- de Haan, N.; Wuhrer, M.; Ruhaak, L. Mass spectrometry in clinical glycomics: The path from biomarker identification to clinical implementation. Clin. Mass Spectrom. 2020, 18, 1–12. [Google Scholar] [CrossRef]
- Colhoun, H.O.; Treacy, E.P.; MacMahon, M.; Rudd, P.M.; Fitzgibbon, M.; O'Flaherty, R.; Stepien, K.M. Validation of an automated ultraperformance liquid chromatography IgG N-glycan analytical method applicable to classical galactosaemia. Ann. Clin. Biochem. 2018, 55, 593–603. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre- and Post-Operative CRC Cohort (n = 64) | ||
|---|---|---|
| Female sex, n (%) | 33 (51.6) | |
| Age in years, mean (s.d.) | 64.2 (12.7) | |
| Stage, n (%) | 1 | 13 (20.3) |
| 2 | 31 (48.4) | |
| 3 | 16 (25.0) | |
| 4 | 4 (6.3) | |
| pT, n (%) | pT1 | 7 (10.9) |
| pT2 | 8 (12.5) | |
| pT3 | 42 (65.6) | |
| pT4 | 7 (10.9) | |
| Histological type, n (%) | Signet-ring cell carcinoma | 3 (4.7) |
| Adenocarcinoma | 54 (84.4) | |
| Mucinous adenocarcinoma | 7 (10.9) | |
| Approach | All N-Glycans | Validated N-Glycans | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Count | CRC Alteration | Count | CRC Alteration | |||||||
| Total | Common | Positional Isomers | Same | Other | Total | Common | Significant * | |||
| # | Same | Other | ||||||||
| RPLC-FD-MS | 48 | 39 | 14 | 30 (77%) | 9 (23%) | n/a | 11 | 8 | 8 (100%) | 0 (0%) |
| MALDI-MS | 83 | 0 | 20 | n/a | ||||||
| Peak | RPLC-FD | Direct N-Glycan Traits | RPLC-MS | RPLC-FD-MS | MALDI-MS |
|---|---|---|---|---|---|
| AUC | AUC | AUC | AUC | ||
| 10 | 0.79 | H5N4F1 | 0.70 | 0.79 | 0.79 |
| H4N4F1 | 0.72 | 0.71 | 0.77 | ||
| n.d. | n.d. | H5N4F1S2,61 | 0.70 | 0.80 | 0.76 |
| n.d. | n.d. | Combination | 0.68 | 0.83 | 0.77 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moran, A.B.; Elgood-Hunt, G.; van der Burgt, Y.E.M.; Wuhrer, M.; Mesker, W.E.; Tollenaar, R.A.E.M.; Spencer, D.I.R.; Lageveen-Kammeijer, G.S.M. Serum N-Glycosylation RPLC-FD-MS Assay to Assess Colorectal Cancer Surgical Interventions. Biomolecules 2023, 13, 896. https://doi.org/10.3390/biom13060896
Moran AB, Elgood-Hunt G, van der Burgt YEM, Wuhrer M, Mesker WE, Tollenaar RAEM, Spencer DIR, Lageveen-Kammeijer GSM. Serum N-Glycosylation RPLC-FD-MS Assay to Assess Colorectal Cancer Surgical Interventions. Biomolecules. 2023; 13(6):896. https://doi.org/10.3390/biom13060896
Chicago/Turabian StyleMoran, Alan B., Georgia Elgood-Hunt, Yuri E. M. van der Burgt, Manfred Wuhrer, Wilma E. Mesker, Rob A. E. M. Tollenaar, Daniel I. R. Spencer, and Guinevere S. M. Lageveen-Kammeijer. 2023. "Serum N-Glycosylation RPLC-FD-MS Assay to Assess Colorectal Cancer Surgical Interventions" Biomolecules 13, no. 6: 896. https://doi.org/10.3390/biom13060896
APA StyleMoran, A. B., Elgood-Hunt, G., van der Burgt, Y. E. M., Wuhrer, M., Mesker, W. E., Tollenaar, R. A. E. M., Spencer, D. I. R., & Lageveen-Kammeijer, G. S. M. (2023). Serum N-Glycosylation RPLC-FD-MS Assay to Assess Colorectal Cancer Surgical Interventions. Biomolecules, 13(6), 896. https://doi.org/10.3390/biom13060896