
Infection, Inflammation, and Immunity in Sepsis


Abstract
:1. Introduction
- An alteration in mental status
- A decrease in systolic blood pressure of less than 100 mm Hg
- A respiration rate greater than 22 breaths/min
2. TLRs and Eicosanoids in Sepsis
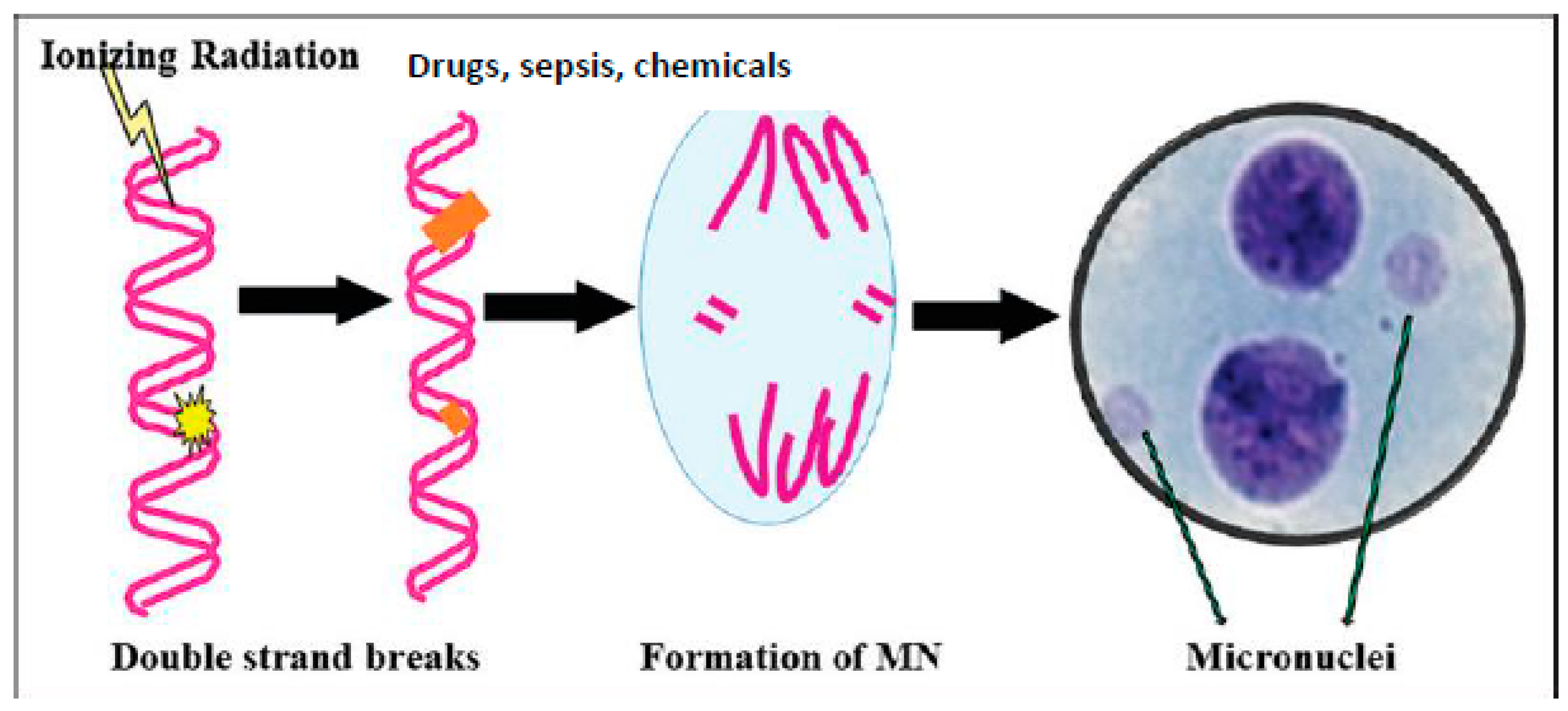
3. The cGAS-STING Pathway, MN Cells, and Sepsis
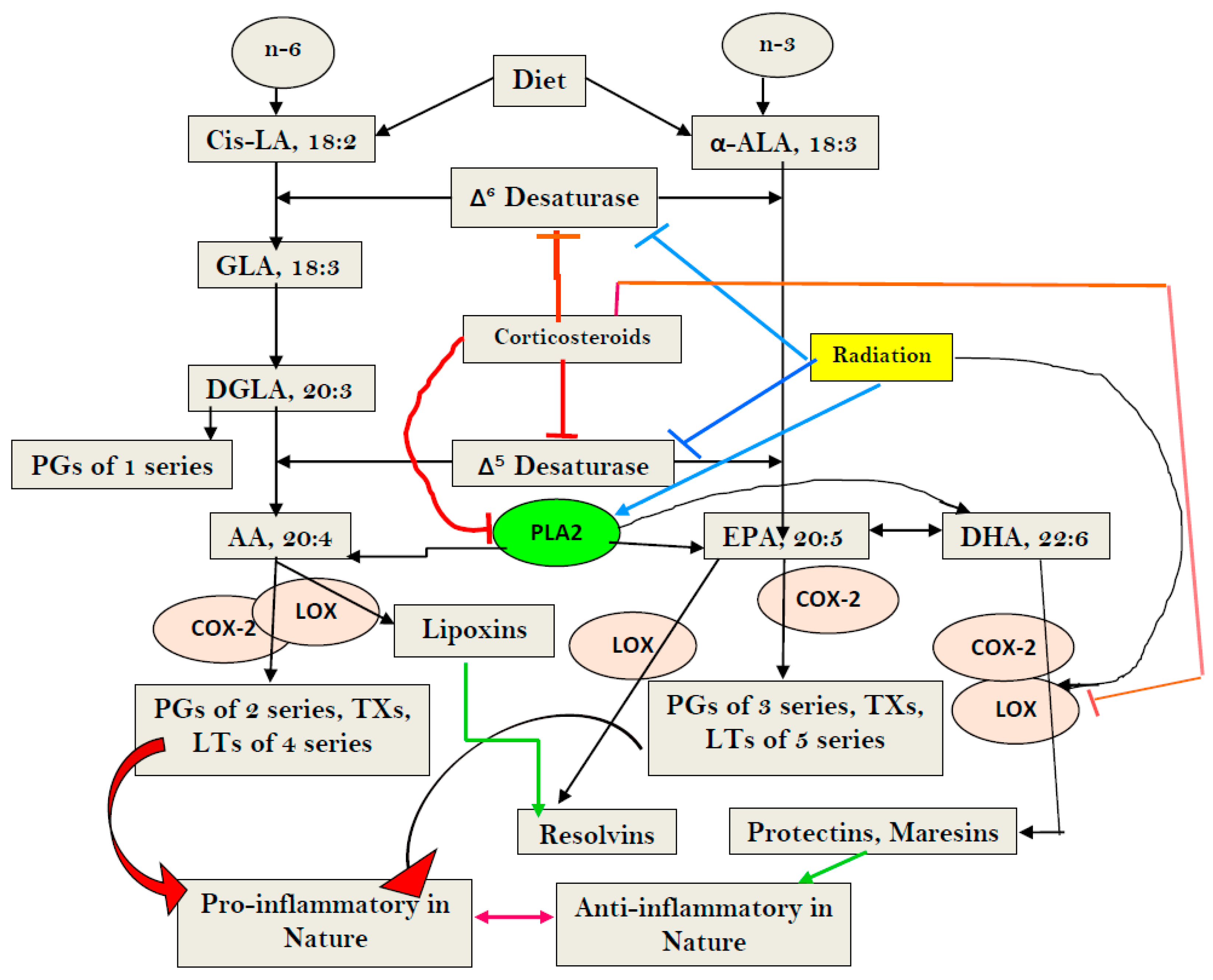
4. Metabolism of Essential Fatty Acids (EFAs)
5. The cGAS-STING System, MN Cells, and EFAs and their Metabolites in Inflammation and Sepsis
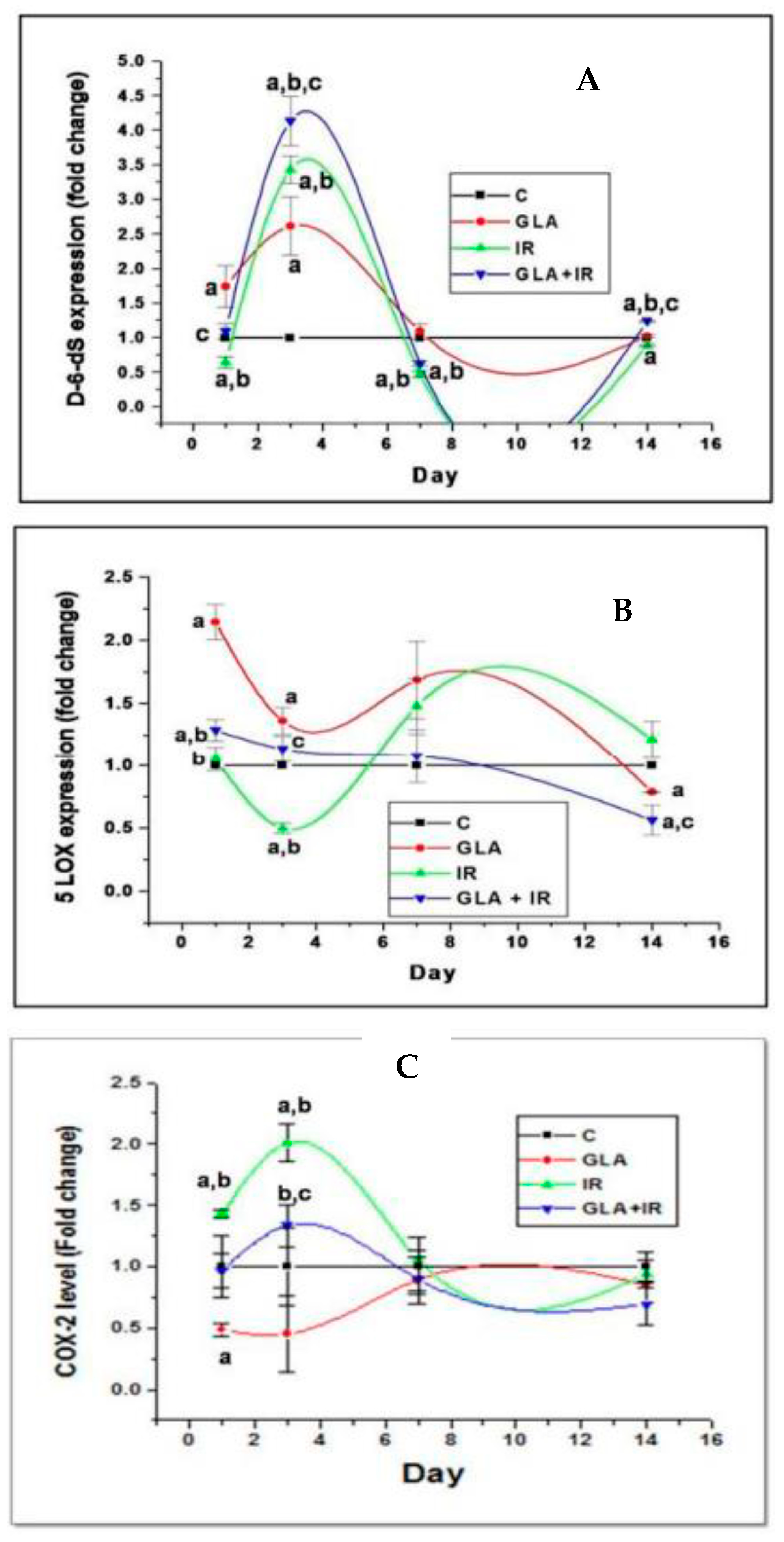
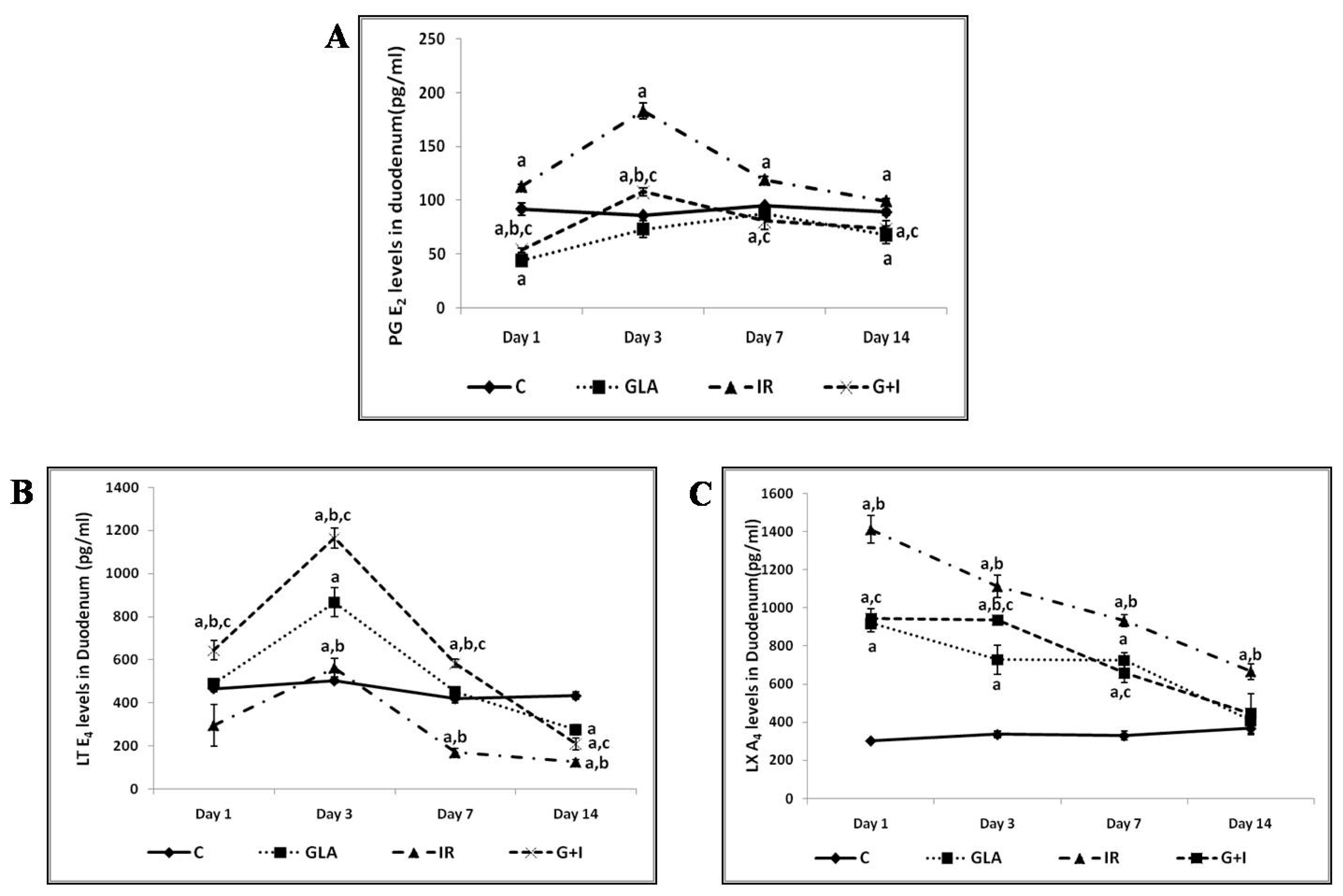
6. Radiation-Induced DNA Damage, Activation of the cGAS-STING System, and EFAs
7. Conclusions and Therapeutic Implications
Funding
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Toti, P.; De Felice, C.; Occhini, R.; Schuerfeld, K.; Stumpo, M.; Epistolato, M.C.; Vatti, R.; Buonocore, G. Spleen depletion in neonatal sepsis and chorioamnionitis. Am. J. Clin. Pathol. 2004, 122, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Monneret, G. How to identify systemic sepsis-induced immunoparalysis. Adv. Sepsis 2005, 4, 42–49. [Google Scholar]
- Monneret, G.; Venet, F.; Pachot, A.; Lepape, A. Monitoring Immune Dysfunctions in the Septic Patient: A New Skin for the Old Ceremony. Mol. Med. 2008, 14, 64–78. [Google Scholar] [CrossRef] [PubMed]
- Arbour, N.C.; Lorenz, E.; Schutte, B.C.; Zabner, J.; Kline, J.N.; Jones, M.; Frees, K.; Watt, J.L.; Schwartz, D.A. TLR4 mutations are associated with endotoxin hyporesponsiveness in humans. Nat. Genet. 2000, 25, 187–191. [Google Scholar] [CrossRef]
- Schwartz, D.A. The role of TLR4 in endotoxin responsiveness in humans. J. Endotoxin Res. 2001, 7, 389–493. [Google Scholar] [CrossRef]
- Albert Vega, C.; Karakike, E.; Bartolo, F.; Mouton, W.; Cerrato, E.; Brengel-Pesce, K.; Giamarellos-Bourboulis, E.J.; Mallet, F.; Trouillet-Assant, S. Differential response induced by LPS and MPLA in immunocompetent and septic individuals. Clin. Immunol. 2021, 226, 108714. [Google Scholar] [CrossRef]
- Holmes, C.L.; Russell, J.A.; Walley, K.R. Genetic polymorphisms in sepsis and septic shock: Role in prognosis and potential for therapy. Chest 2003, 124, 1103–1115. [Google Scholar] [CrossRef]
- Imahara, S.D.; Jelacic, S.; Junker, C.E.; O’Keefe, G.E. The TLR4 +896 polymorphism is not associated with lipopolysaccharide hypo-responsiveness in leukocytes. Genes Immun. 2005, 6, 37–43. [Google Scholar] [CrossRef]
- Shishido, T.; Nozaki, N.; Takahashi, H.; Arimoto, T.; Niizeki, T.; Koyama, Y.; Abe, J.-I.; Takeishi, Y.; Kubota, I. Central role of endogenous Toll-like receptor-2 activation in regulating inflammation, reactive oxygen species production, and subsequent neointimal formation after vascular injury. Biochem. Biophys. Res. Commun. 2006, 345, 1446–1453. [Google Scholar] [CrossRef]
- Norris, P.C.; Reichart, D.; Dumlao, D.S.; Glass, C.K.; Dennis, E.A. Specificity of eicosanoid production depends on the TLR-4-stimulated macrophage phenotype. J. Leukoc. Biol. 2011, 90, 563–574. [Google Scholar] [CrossRef]
- Lefebvre, J.S.; Marleau, S.; Milot, V.; Lévesque, T.; Picard, S.; Flamand, N.; Borgeat, P. Toll-like receptor ligands induce polymorphonuclear leukocyte migration: Key roles for leukotriene B4 and platelet-activating factor. FASEB J. 2010, 24, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Arachidonic Acid as Mechanotransducer of Renin Cell Baroreceptor. Nutrients 2022, 14, 749. [Google Scholar] [CrossRef] [PubMed]
- Lomakin, A.J.; Cattin, C.J.; Cuvelier, D.; Alraies, Z.; Molina, M.; Nader, G.P.F.; Srivastava, N.; Sáez, P.J.; Garcia-Arcos, J.M.; Zhitnyak, I.Y.; et al. The nucleus acts as a ruler tailoring cell responses to spatial constraints. Science 2020, 370, eaba2894. [Google Scholar] [CrossRef] [PubMed]
- Venturini, V.; Pezzano, F.; Castro, F.C.; Häkkinen, H.-M.; Jiménez-Delgado, S.; Colomer-Rosell, M.; Marro, M.; Tolosa-Ramon, Q.; Paz-López, S.; Valverde, M.A.; et al. The nucleus measures shape changes for cellular proprioception to control dynamic cell behavior. Science 2020, 370, eaba2644. [Google Scholar] [CrossRef]
- Shen, Z.; Niethammer, P. A cellular sense of space and pressure. Science 2020, 370, 295–296. [Google Scholar] [CrossRef]
- Enyedi, B.; Kala, S.; Nikolich-Zugich, T.; Niethammer, P. Tissue damage detection by osmotic surveillance. Nature 2013, 15, 1123–1130. [Google Scholar] [CrossRef]
- Sapirstein, A.; Bonventre, J.V. Specific physiological roles of cytosolic phospholipase A2 as defined by gene knockouts. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2000, 1488, 139–148. [Google Scholar] [CrossRef]
- Font-Nieves, M.; Sans-Fons, M.G.; Gorina, R.; Bonfill-Teixidor, E.; SalasPerdomo, A.; Marquez-Kisinousky, L. Induction of COX-2 Enzyme and Down-regulation of COX-1 Expression by Lipopolysaccharide (LPS) Control Prostaglandin E2 Production in Astrocytes. J. Biol. Chem. 2012, 287, 6454–6468. [Google Scholar] [CrossRef]
- Gorina, R.; Font-Nieves, M.; Márquez-Kisinousky, L.; Santalucia, T.; Planas, A.M. Astrocyte TLR4 activation induces a proinflammatory environment through the interplay between MyD88-dependent NFκB signaling, MAPK, and Jak1/Stat1 pathways. Glia 2011, 59, 242–255. [Google Scholar] [CrossRef]
- Balistreri, C.R.; Caruso, C.; Listì, F.; Colonna-Romano, G.; Lio, D.; Candore, G. LPS-mediated production of pro/anti-inflammatory cytokines and eicosanoids in whole blood samples: Biological effects of +896A/G TLR4 polymorphism in a Sicilian population of healthy subjects. Mech. Ageing Dev. 2011, 132, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Jilling, T.; Li, D.; Caplan, M.S. Polyunsaturated Fatty Acid Supplementation Alters Proinflammatory Gene Expression and Reduces the Incidence of Necrotizing Enterocolitis in a Neonatal Rat Model. Pediatr. Res. 2007, 61, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Caplan, M.S.; Russell, T.; Xiao, Y.; Amer, M.; Kaup, S.; Jilling, T. Effect of Polyunsaturated Fatty Acid (PUFA) Supplementation on Intestinal Inflammation and Necrotizing Enterocolitis (NEC) in a Neonatal Rat Model. Pediatr. Res. 2001, 49, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Duffield, J.S.; Hong, S.; Vaidya, V.S.; Lu, Y.; Fredman, G.; Serhan, C.N.; Bonventre, J.V. Resolvin D Series and Protectin D1 Mitigate Acute Kidney Injury. J. Immunol. 2006, 177, 5902–5911. [Google Scholar] [CrossRef]
- Hassan, I.R.; Gronert, K. Acute changes in dietary omega-3 and omega-6 polyunsaturated fatty acids have a pronounced impact on survival following ischemic renal injury and formation of renoprotective docosahexaenoic acid-derived protectin D1. J. Immunol. 2009, 182, 3223–3322. [Google Scholar] [CrossRef]
- Spite, M.; Norling, L.V.; Summers, L.; Yang, R.; Cooper, D.; Petasis, N.A.; Flower, R.J.; Perretti, M.; Serhan, C.N. Resolvin D2 is a potent regulator of leukocytes and controls microbial sepsis. Nature 2009, 461, 1287–1292. [Google Scholar] [CrossRef]
- Das, U. HLA-DR expression, cytokines and bioactive lipids in sepsis. Arch. Med. Sci. 2014, 10, 325–335. [Google Scholar] [CrossRef]
- Das, U.N. Pro- and anti-inflammatory bioactive lipids imbalance contributes to the pathobiology of autoimmune diseases. Eur. J. Clin. Nutr. 2022, 77, 637–651. [Google Scholar] [CrossRef]
- Pinheiro da Silva, F.; Nizet, V. Cell death during sepsis: Integration of disintegration in the inflammatory response to overwhelming infection. Apoptosis 2009, 14, 509–521. [Google Scholar] [CrossRef]
- Timmermans, K.; Kox, M.; Scheffer, G.J.; Pickkers, P. Faculty Opinions recommendation of Plasma Nuclear and Mitochondrial DNA Levels, and Markers of Inflammation, Shock, and Organ Damage in Patients with Septic Shock. Shock 2016, 45, 607–612. [Google Scholar] [CrossRef]
- Di Caro, V.; Walko, T.D., III; Bola, R.A.; Hong, J.D.; Pang, D.; Hsue, V.; Au, A.K.; Halstead, E.S.; Carcillo, J.A.; Clark, R.S.B.; et al. Plasma mitochondrial DNA—A novel DAMP in pediatric sepsis. Shock 2016, 45, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Heipertz, E.L.; Harper, J.; Walker, W.E. STING and TRIF contribute to mouse sepsis, depending on severity of the disease model. Shock 2017, 47, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.-X.; Kang, R.; Tang, D.-L. STING1 in sepsis: Mechanisms, functions, and implications. Chin. J. Traumatol. 2022, 25, 1–10. [Google Scholar] [CrossRef]
- Zhang, Q.; Raoof, M.; Chen, Y.; Sumi, Y.; Sursal, T.; Junger, W.; Brohi, K.; Itagaki, K.; Hauser, C.J. Circulating mitochondrial DAMPs cause inflammatory responses to injury. Nature 2010, 464, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, N.; Tsuji, T.; Ohashi, N.; Kato, A.; Fujigaki, Y.; Yasuda, H. Role of mitochondrial DNA in septic AKI via toll-like receptor 9. J. Am. Soc. Nephrol. 2016, 27, 2009–2020. [Google Scholar] [CrossRef] [PubMed]
- Ablasser, A.; Goldeck, M.; Cavlar, T.; Deimling, T.; Witte, G.; Röhl, I.; Hopfner, K.-P.; Ludwig, J.; Hornung, V. cGAS produces a 2′-5′-linked cyclic dinucleotide second messenger that activates STING. Nature 2013, 498, 380–384. [Google Scholar] [CrossRef]
- Civril, F.; Deimling, T.; Mann, C.C.d.O.; Ablasser, A.; Moldt, M.; Witte, G.; Hornung, V.; Hopfner, K.-P. Structural mechanism of cytosolic DNA sensing by cGAS. Nature 2013, 498, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Ablasser, A.; Schmid-Burgk, J.L.; Hemmerling, I.; Horvath, G.L.; Schmidt, T.; Latz, E.; Hornung, V. Cell intrinsic immunity spreads to bystander cells via the intercellular transfer of cGAMP. Nature 2013, 503, 530–534. [Google Scholar] [CrossRef]
- Schoggins, J.W.; MacDuff, D.A.; Imanaka, N.; Gainey, M.D.; Shrestha, B.; Eitson, J.L.; Mar, K.B.; Richardson, R.B.; Ratushny, A.V.; Litvak, V.; et al. Pan-viral specificity of IFN-induced genes reveals new roles for cGAS in innate immunity. Nature 2014, 505, 691–695. [Google Scholar] [CrossRef]
- Harding, S.M.; Benci, J.L.; Irianto, J.; Discher, D.E.; Minn, A.J.; Greenberg, R.A. Mitotic progression following DNA damage enables pattern recognition within micronuclei. Nature 2017, 548, 466–470. [Google Scholar] [CrossRef]
- Das, U.N. Molecular pathobiology of scleritis and its therapeutic implications. Int. J. Ophthalmol. 2020, 13, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Karaman, A.; Kadi, M.; Kara, F. Sister chromatid exchange and micronucleus studies in patients with Behçet’s disease. J. Cutan. Pathol. 2009, 36, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Hamurcu, Z.; Donmez-Altuntas, H.; Borlu, M.; Demirtas, H.; Ascioslu, O. Micronucleus frequency in the oral mucosa and lymphocytes of patients with Behcet’s disease. Clin. Exp. Dermatol. 2005, 30, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Sommer, S.; Buraczewska, I.; Kruszewski, M. Micronucleus Assay: The State of Art, and Future Directions. Int. J. Mol. Sci. 2020, 21, 1534. [Google Scholar] [CrossRef] [PubMed]
- Franzke, B.; Schwingshackl, L.; Wagner, K.-H. Chromosomal damage measured by the cytokinesis block micronucleus cytome assay in diabetes and obesity—A systematic review and meta-analysis. Mutat. Res. Mol. Mech. Mutagen. 2020, 786, 108343. [Google Scholar] [CrossRef]
- Torres-Bugarín, O.; Romero, N.M.; Ibarra, M.L.R.; Flores-García, A.; Aburto, P.V.; Zavala-Cerna, M.G. Genotoxic Effect in Autoimmune Diseases Evaluated by the Micronucleus Test Assay: Our Experience and Literature Review. BioMed Res. Int. 2015, 2015, 194031. [Google Scholar] [CrossRef]
- Das, U.N. Molecular Biochemical Aspects of Cancer; Humana Press: New York, NY, USA, 2020. [Google Scholar]
- Nakanishi, A.; Tsukamoto, I. n-3 polyunsaturated fatty acids stimulate osteoclastogenesis through PPARγ-mediated enhancement of c-Fos expression, and suppress osteoclastogenesis through PPARγ-dependent inhibition of NFkB activation. J. Nutr. Biochem. 2015, 26, 1317–1327. [Google Scholar] [CrossRef]
- Das, U.N. “Cell Membrane Theory of Senescence” and the Role of Bioactive Lipids in Aging, and Aging Associated Diseases and Their Therapeutic Implications. Biomolecules 2021, 11, 241. [Google Scholar] [CrossRef]
- Das, U.N. Bioactive Lipids in Age-Related Disorders. Adv. Exp. Med. Biol. 2020, 1260, 33–83. [Google Scholar] [CrossRef]
- Yin, Y.; Chen, F.; Wang, W.; Wang, H.; Zhang, X. Resolvin D1 inhibits inflammatory response in STZ-induced diabetic retinopathy rats: Possible involvement of NLRP3 inflammasome and NF-κB signaling pathway. Mol. Vis. 2017, 23, 242–250. [Google Scholar]
- Das, U.N.; Devi, G.R.; Rao, K.P.; Rao, M.S. Prostaglandins and their precursors can modify genetic damage induced by gamma-radiation and benzo(a)pyrene. Prostaglandins 1985, 29, 911–919. [Google Scholar] [CrossRef]
- Koratkar, R.; Das, U.N.; Sagar, P.S.; Ramesh, G.; Padma, M.; Kumar, G.S.; Vijay, K.; Madhavi, N. Prostacyclin is a potent anti-mutagen. Prostaglandins Leukot. Essent. Fat. Acids 1993, 48, 175–184. [Google Scholar] [CrossRef]
- Ramadevi, G.; Das, U.; Rao, K.; Rao, M. Prostaglandins and mutagenesis: Prevention and/or reversibility of genetic damage induced by benzo (a) pyrene in the bone marrow cells of mice by prostaglandin El. Prostaglandins Leukot. Med. 1984, 15, 287–292. [Google Scholar] [CrossRef]
- Ramadevi, G.; Das, U.; Rao, K.; Rao, M. Prostaglandins and mutagenesis: Modification of phenytoin induced genetic damage by prostaglandins in lymphocyte cultures. Prostaglandins Leukot. Med. 1984, 15, 109–113. [Google Scholar] [CrossRef]
- Das, U.N.; Rao, K.P. Effect of γ-linolenic acid and prostaglandins E1 on gamma-radiation and chemical-induced genetic damage to the bone marrow cells of mice. Prostaglandins Leukot. Essent. Fat. Acids 2006, 74, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Ponnala, S.; Rao, K.P.; Chaudhury, J.R.; Ahmed, J.; Rao, B.R.; Kanjilal, S.; Hasan, Q.; Das, U.N. Effect of polyunsaturated fatty acids on diphenyl hydantoin-induced genetic damage invitro and in vivo. Prostaglandins Leukot. Essent. Fat. Acids 2009, 80, 43–50. [Google Scholar] [CrossRef]
- Das, U. Tumoricidal action of cis-unsaturated fatty acids and their relationship to free radicals and lipid peroxidation. Cancer Lett. 1991, 56, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N.; Huang, Y.-S.; Bēgin, M.E.; Ells, G.; Horrobin, D.F. Uptake and distribution of cis-unsaturated fatty acids and their effect on free radical generation in normal and tumor cells in vitro. Free. Radic. Biol. Med. 1987, 3, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Essential fatty acids enhance free radical generation and lipid peroxidation to induce apoptosis of tumor cells. Clin. Lipidol. 2011, 6, 463–489. [Google Scholar] [CrossRef]
- Viswanathan, V.S.; Ryan, M.J.; Dhruv, H.D.; Gill, S.; Eichhoff, O.M.; Seashore-Ludlow, B.; Kaffenberger, S.D.; Eaton, J.K.; Shimada, K.; Aguirre, A.J.; et al. Dependency of a therapy-resistant state of cancer cells on a lipid peroxidase pathway. Nature 2017, 547, 453–457. [Google Scholar] [CrossRef]
- Hangauer, M.J.; Viswanathan, V.S.; Ryan, M.J.; Bole, D.; Eaton, J.K.; Matov, A.; Galeas, J.; Dhruv, H.D.; Berens, M.E.; Schreiber, S.L.; et al. Drug-tolerant persister cancer cells are vulnerable to GPX4 inhibition. Nature 2017, 551, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Polavarapu, S.; Mani, A.M.; Gundala, N.K.V.; Hari, A.D.; Bathina, S.; Das, U.N. Effect of Polyunsaturated Fatty Acids and Their Metabolites on Bleomycin-Induced Cytotoxic Action on Human Neuroblastoma Cells In Vitro. PLoS ONE 2014, 9, e114766. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ma, Y.; Ma, J.; Yang, L.; Song, Q.; Wang, H.; Lv, G. Glutathione Peroxidase 4 as a Therapeutic Target for Anti-Colorectal Cancer Drug-Tolerant Persister Cells. Front. Oncol. 2022, 12, 913669. [Google Scholar] [CrossRef] [PubMed]
- Croasdell, A.; Lacy, S.H.; Thatcher, T.H.; Sime, P.J.; Phipps, R.P. Resolvin D1 Dampens Pulmonary Inflammation and Promotes Clearance of Nontypeable Haemophilus influenzae. J. Immunol. 2016, 196, 2742–2752. [Google Scholar] [CrossRef]
- Bathina, S.; Gundala, N.K.; Rhenghachar, P.; Polavarapu, S.; Hari, A.D.; Sadananda, M.; Das, U.N. Resolvin D1 Ameliorates Nicotinamide-streptozotocin-induced Type 2 Diabetes Mellitus by its Anti-inflammatory Action and Modulating PI3K/Akt/mTOR Pathway in the Brain. Arch. Med. Res. 2020, 51, 492–503. [Google Scholar] [CrossRef]
- Kang, J.X.; Weylandt, K.H. Modulation of Inflammatory Cytokines by Omega-3 Fatty Acids. In Lipids in Health and Disease; Quinn, P.J., Wang, X., Eds.; Subcellular Biochemistry Series; Springer: Dordrecht, The Netherlands, 2008; Volume 49, pp. 133–143. [Google Scholar] [CrossRef]
- Gundala, N.K.V.; Naidu, V.G.M.; Das, U.N. Arachidonic acid and lipoxin A4 attenuate alloxan-induced cytotoxicity to RIN5F cells in vitro and type 1 diabetes mellitus in vivo. Biofactors 2017, 43, 251–271. [Google Scholar] [CrossRef]
- Rengachar, P.; Polavarapu, S.; Das, U.N. Insights in diabetes: Molecular mechanisms-Protectin DX, an anti-inflammatory and a stimulator of inflammation resolution metabolite of docosahexaenoic acid, protects against the development of streptozotocin-induced type 1 and type 2 diabetes mellitus in male Swiss albino mice. Front. Endocrinol. 2023, 13, 1053879. [Google Scholar] [CrossRef]
- Bahar, I.; Elay, G.; Başkol, G.; Sungur, M.; Donmez-Altuntas, H. Increased DNA damage and increased apoptosis and necrosis in patients with severe sepsis and septic shock. J. Crit. Care 2018, 43, 271–275. [Google Scholar] [CrossRef]
- Ramos-Remus, C.; Dorazco-Barragan, G.; Aceves-Avila, F.J.; Alcaraz-Lopez, F.; Fuentes-Ramirez, F.; Michel-Diaz, J.; Torres-Bugarin, O.; Ventura-Aguilar, A.; Zuñiga-González, G. Genotoxicity assessment using micronuclei assay in rheumatoid arthritis patients. Clin. Exp. Rheumatol. 2002, 20, 208–212. [Google Scholar]
- Al-Rawi, Z.S.; Gorial, F.I.; Tawfiq, R.F.; Mohammed, A.K.; Al-Naaimi, A.S.; Al’Aadhmi, M.A.; Hayyawi, A.A. Brief Report: A Novel Application of Buccal Micronucleus Cytome Assay in Systemic Lupus Erythematosus: A Case-Control Study. Arthritis Rheumatol. 2014, 66, 2837–2841. [Google Scholar] [CrossRef]
- Baig, A.; Avlasevich, S.L.; Torous, D.K.; Bemis, J.C.; Saubermann, L.J.; Lovell, D.P.; MacGregor, J.T.; Dertinger, S.D. Assessment of systemic genetic damage in pediatric inflammatory bowel disease. Environ. Mol. Mutagen. 2020, 61, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Constanzo, J.; Faget, J.; Urssino, C.; Badle, C.; Pouget, J.-P. Radiation-Induced Immunity and Toxicities: The Versatility of the cGAS-STING Pathway. Front. Immunol. 2012, 12, 680503. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R.; Heo, J.I.; Park, J.W.; Kang, C.M.; Kim, K.S. Radiation-induced lipoprotein-associated phospholipase A2 increases lysophosphatidylcholine and induces endothelial cell damage. Toxicology 2021, 458, 152841. [Google Scholar] [CrossRef]
- Yang, H.J.; Youn, H.; Seong, K.M.; Yun, Y.J.; Kim, W.; Kim, Y.H.; Lee, J.Y.; Kim, C.S.; Jin, Y.-W.; Youn, B. Psoralidin, a dual inhibitor of COX-2 and 5-LOX, regulates ionizing radiation (IR)-induced pulmonary inflammation. Biochem. Pharmacol. 2011, 82, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Du, S.S.; Chen, G.W.; Yang, P.; Chen, Y.X.; Hu, Y.; Zhao, Q.Q.; Zhang, Y.; Liu, R.; Zheng, D.X.; Zhou, J.; et al. Radiation Therapy Promotes Hepatocellular Carcinoma Immune Cloaking via PD-L1 Upregulation Induced by cGAS-STING Activation. Int. J. Radiat. Oncol. Biol. Phys. 2022, 112, 1243–1255. [Google Scholar] [CrossRef]
- Kanno, T.; Nakajima, T.; Yokoyama, S.; Asou, H.K.; Sasamoto, S.; Kamii, Y.; Hayashizaki, K.; Ouchi, Y.; Onodera, T.; Takahashi, Y.; et al. SCD2-mediated monounsaturated fatty acid metabolism regulates cGAS-STING-dependent type I IFN responses in CD4+ T cells. Commun. Biol. 2021, 4, 820. [Google Scholar] [CrossRef]
- Vila, I.K.; Chamma, H.; Steer, A.; Saccas, M.; Taffoni, C.; Turtoi, E.; Reinert, L.S.; Hussain, S.; Marines, J.; Jin, L.; et al. STING orchestrates the crosstalk between polyunsaturated fatty acid metabolism and inflammatory responses. Cell Metab. 2022, 34, 125–139. [Google Scholar] [CrossRef]
- Prabha, P.S.; Das, U.; Ramesh, G.; Kumar, K.V.; Kamalakar, V. Free radical generation, lipid peroxidation and essential fatty acids in patients with septicemia. Prostaglandins Leukot. Essent. Fat. Acids 1991, 42, 61–65. [Google Scholar] [CrossRef]
- Rengachar, P.; Bhatt, A.N.; Polavarapu, S.; Veeramani, S.; Krishnan, A.; Sadananda, M.; Das, U.N. Gamma-Linolenic Acid (GLA) Protects against Ionizing Radiation-Induced Damage: An In Vitro and In Vivo Study. Biomolecules 2022, 12, 797. [Google Scholar] [CrossRef]
- Tateishi, N.; Kakutani, S.; Kawashima, H.; Shibata, H.; Morita, I. Dietary supplementation of arachidonic acid increases arachidonic acid and lipoxin A4 contents in colon, but does not affect severity or prostaglandin E2 content in murine colitis model. Lipids Health Dis. 2014, 13, 30. [Google Scholar] [CrossRef]
- Tateishi, N.; Kaneda, Y.; Kakutani, S.; Kawashima, H.; Shibata, H.; Morita, I. Dietary supplementation with arachidonic acid increases arachidonic acid content in paw, but does not affect arthritis severity or prostaglandin E2 content in rat adjuvant-induced arthritis model. Lipids Health Dis. 2015, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Duvall, M.G.; Barnig, C.; Cernadas, M.; Ricklefs, I.; Krishnamoorthy, N.; Grossman, N.L.; Bhakta, N.R.; Fahy, J.V.; Bleecker, E.R.; Castro, M.; et al. National Heart, Lung, and Blood Institute’s Severe Asthma Research Program-3 Investigators. Natural killer cell-mediated inflammation resolution is disabled in severe asthma. Sci. Immunol. 2017, 2, eaam5446. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Is sepsis a pro-resolution deficiency disorder? Med. Hypotheses 2013, 80, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Polyunsaturated fatty acids and sepsis. Nutrition 2019, 65, 39–43. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
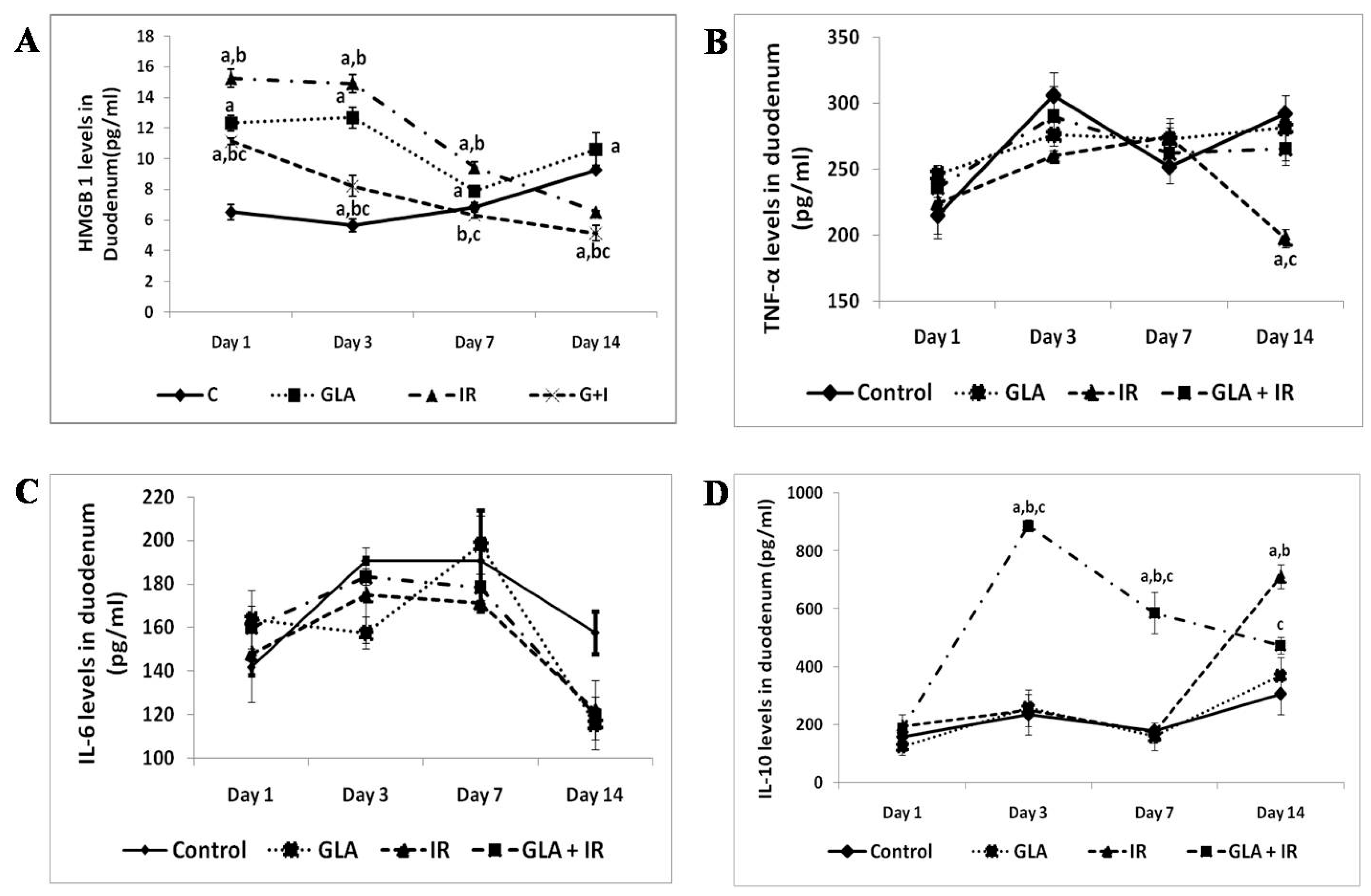{kind=link}
| Fatty Acid | Control (n = 10) | Septicemia (n = 14) |
|---|---|---|
| 16:0 | 24.8 ± 3.4 | 26.95 ± 1.1 |
| 18:0 | 23.3 ± 4.1 | 24.58 ± 6.0 |
| 18:1 n-9 | 13.1 ± 2.3 | 16.5 ± 3.3 * |
| 18:2 n-6 (LA) | 17.7 ± 3.1 | 16.3 ± 3.3 |
| 18:3 n-6 (GLA) | 0.13 ± 0.09 | 0.04 ± 0.05 * |
| 20:3 n-6 (DGLA) | 3.2 ± 0.79 | 0.46 ± 0.54 * |
| 20:4 n-6 (AA) | 8.8 ± 2.0 | 5.8 ± 1.6 * |
| 18:3 n-3 (ALA) | 0.27 ± 0.12 | 0.16 ± 0.11 * |
| 20:5 n-3 (EPA) | 0.25 ± 0.26 | 0.01 ± 0.01 * |
| 22:6 n-3 (DHA) | 1.43 ± 0.43 | 1.2 ± 1.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Das, U.N. Infection, Inflammation, and Immunity in Sepsis. Biomolecules 2023, 13, 1332. https://doi.org/10.3390/biom13091332
Das UN. Infection, Inflammation, and Immunity in Sepsis. Biomolecules. 2023; 13(9):1332. https://doi.org/10.3390/biom13091332
Chicago/Turabian StyleDas, Undurti N. 2023. "Infection, Inflammation, and Immunity in Sepsis" Biomolecules 13, no. 9: 1332. https://doi.org/10.3390/biom13091332
APA StyleDas, U. N. (2023). Infection, Inflammation, and Immunity in Sepsis. Biomolecules, 13(9), 1332. https://doi.org/10.3390/biom13091332