

Characterization of the Abracl-Expressing Cell Populations in the Embryonic Mammalian Telencephalon

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Tissue Preparation, Fixation and Sectioning
2.3. Primary Cultures
2.4. Immunofluorescence
2.5. Microscope Imaging and Processing
3. Results
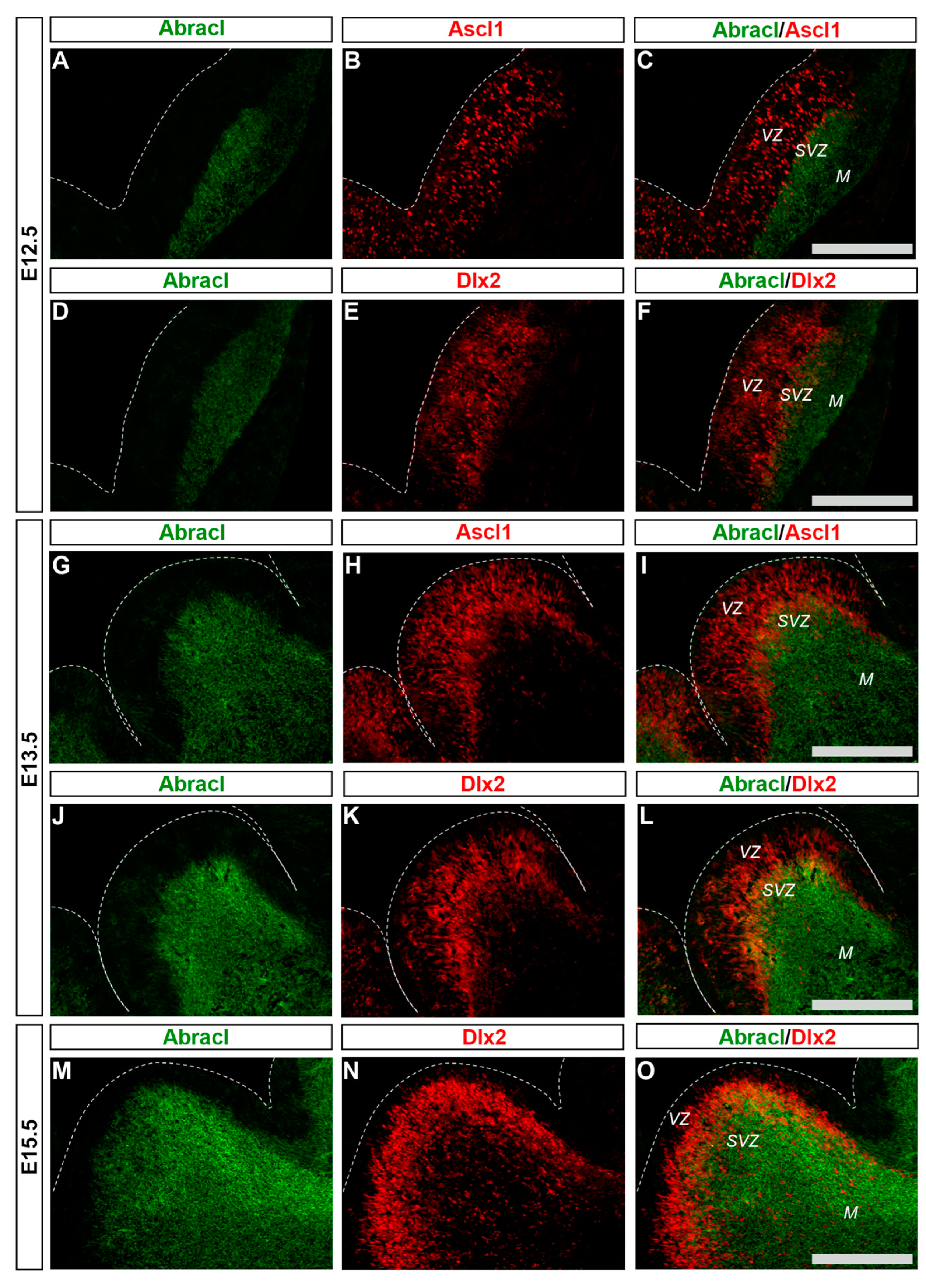
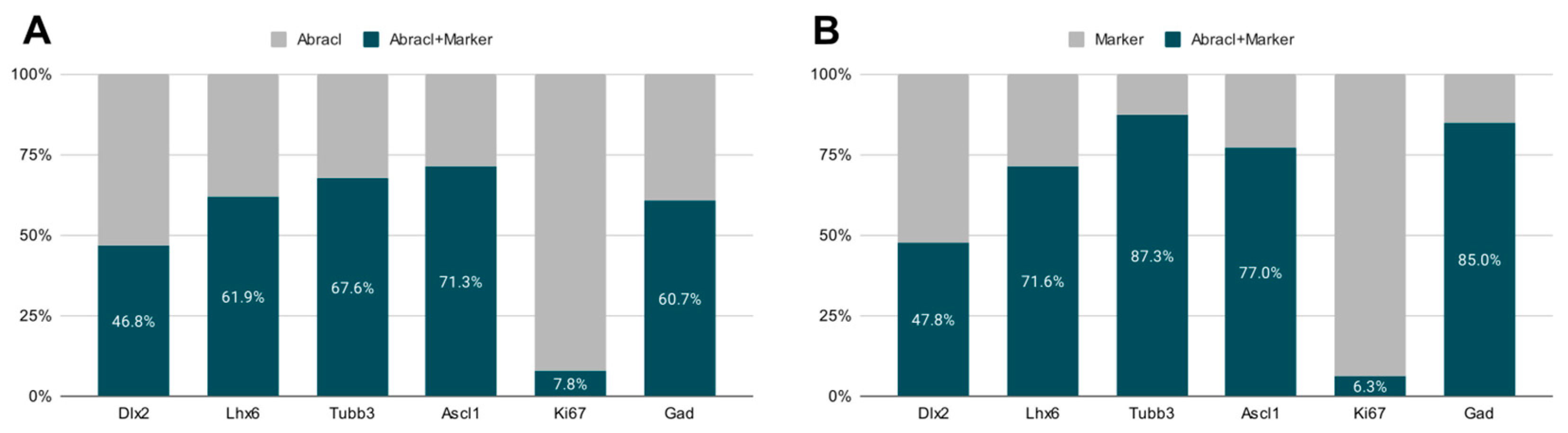
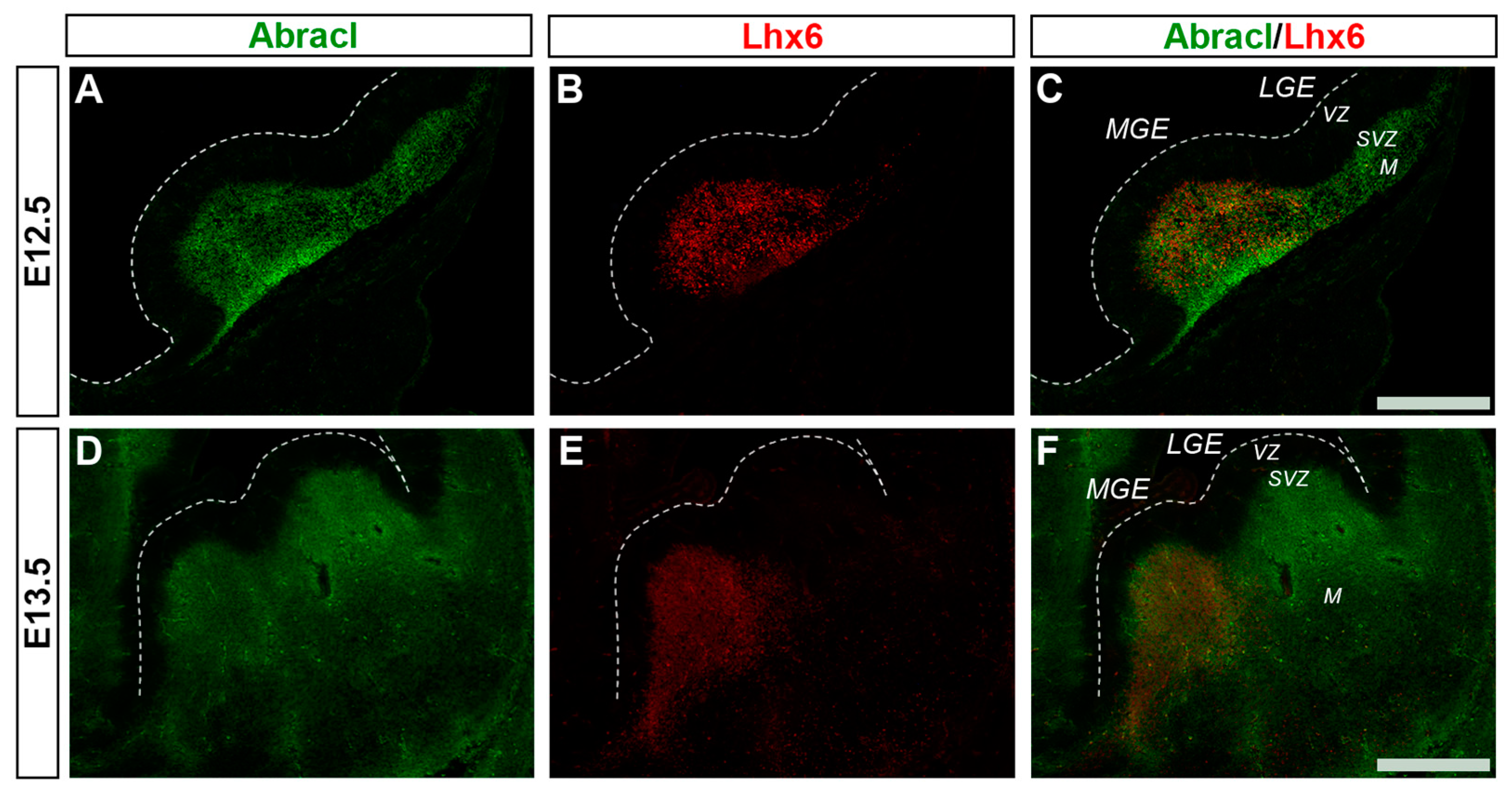
3.1. In the Subpallium, Abracl Is Expressed by Proliferating and Post-Mitotic Cells
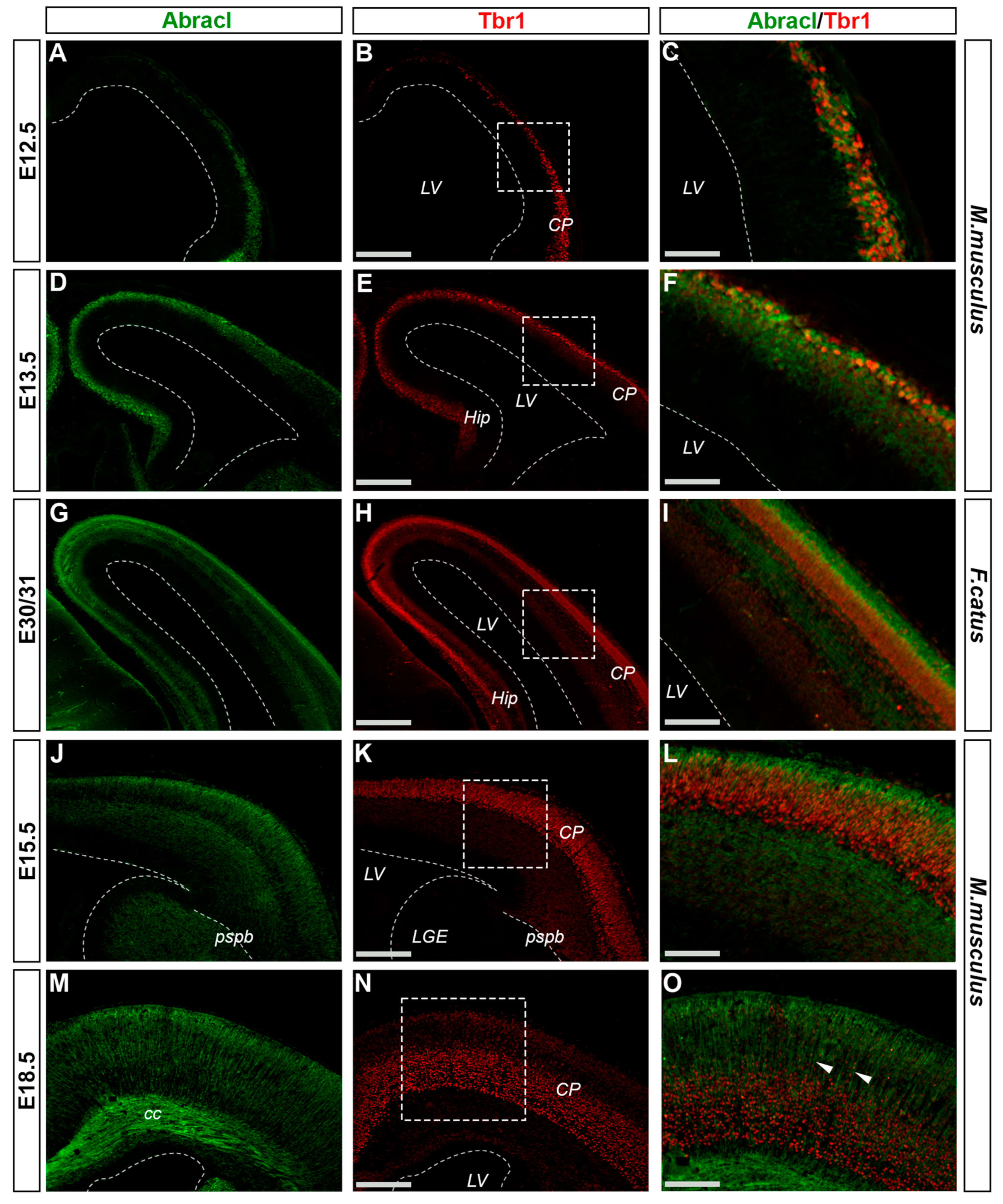
3.2. In the Pallium, Abracl Is Expressed Only by Post-Mitotic Cells
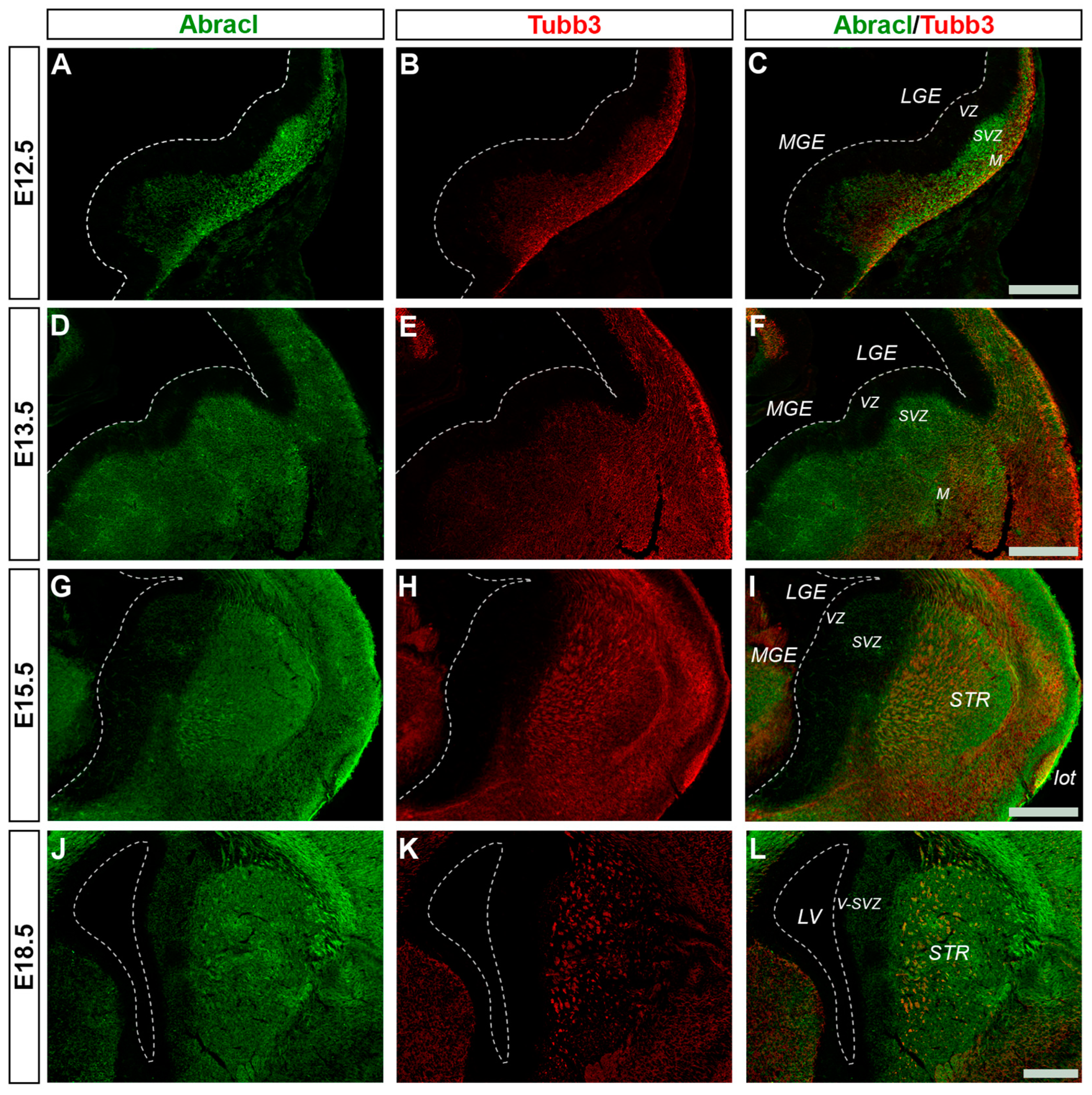
3.3. Abracl Is Expressed in Major Telencephalic Fiber Tracts
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lin, J.; Zhou, T.; Wang, J. Solution Structure of the Human HSPC280 Protein: Solution Structure of HSPC280. Protein Sci. 2011, 20, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Pang, T.-L.; Chen, F.-C.; Weng, Y.-L.; Liao, H.-C.; Yi, Y.-H.; Ho, C.-L.; Lin, C.-H.; Chen, M.-Y. Costars, a Dictyostelium Protein Similar to the C-Terminal Domain of STARS, Regulates the Actin Cytoskeleton and Motility. J. Cell Sci. 2010, 123, 3745–3755. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Chen, P.; Li, S. miR-145-5p Inhibits the Proliferation, Migration and Invasion of Esophageal Carcinoma Cells by Targeting ABRACL. Biomed Res. Int. 2021, 2021, 6692544. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, B.-Y.; Chen, C.-H.; Chi, H.-Y.; Yen, P.-R.; Yu, Y.-Z.; Lin, C.-H.; Pang, T.-L.; Lin, W.-C.; Li, M.-L.; Yeh, Y.-C.; et al. Human Costars Family Protein ABRACL Modulates Actin Dynamics and Cell Migration and Associates with Tumorigenic Growth. Int. J. Mol. Sci. 2021, 22, 2037. [Google Scholar] [CrossRef]
- Li, J.; Chen, H. Actin-Binding Rho Activating C-Terminal like (ABRACL) Transcriptionally Regulated by MYB Proto-Oncogene like 2 (MYBL2) Promotes the Proliferation, Invasion, Migration and Epithelial-Mesenchymal Transition of Breast Cancer Cells. Bioengineered 2022, 13, 9019–9031. [Google Scholar] [CrossRef]
- Ura, B.; Monasta, L.; Arrigoni, G.; Franchin, C.; Radillo, O.; Peterlunger, I.; Ricci, G.; Scrimin, F. A Proteomic Approach for the Identification of Biomarkers in Endometrial Cancer Uterine Aspirate. Oncotarget 2017, 8, 109536–109545. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, T.; Xie, F.; Wang, L.; Liang, Z.; Li, D.; Liang, Y.; Zhao, K.; Qi, X.; Yang, X.; et al. Evaluating the Biological Functions of the Prognostic Genes Identified by the Pathology Atlas in Bladder Cancer. Oncol. Rep. 2020, 45, 191–201. [Google Scholar] [CrossRef]
- Wang, D.; Liu, H.; Ren, C.; Wang, L. High Expression of ABRACL Is Associated with Tumorigenesis and Affects Clinical Outcome in Gastric Cancer. Genet. Test. Mol. Biomark. 2019, 23, 91–97. [Google Scholar] [CrossRef]
- Gupta, G.D.; Coyaud, É.; Gonçalves, J.; Mojarad, B.A.; Liu, Y.; Wu, Q.; Gheiratmand, L.; Comartin, D.; Tkach, J.M.; Cheung, S.W.T.; et al. A Dynamic Protein Interaction Landscape of the Human Centrosome-Cilium Interface. Cell 2015, 163, 1484–1499. [Google Scholar] [CrossRef]
- Monaghan, J.R.; Walker, J.A.; Page, R.B.; Putta, S.; Beachy, C.K.; Voss, S.R. Early Gene Expression during Natural Spinal Cord Regeneration in the Salamander Ambystoma Mexicanum: Gene Expression and Spinal Cord Regeneration. J. Neurochem. 2006, 101, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Stylianopoulou, E.; Kalamakis, G.; Pitsiani, M.; Fysekis, I.; Ypsilantis, P.; Simopoulos, C.; Skavdis, G.; Grigoriou, M.E. HSPC280, a Winged Helix Protein Expressed in the Subventricular Zone of the Developing Ganglionic Eminences, Inhibits Neuronal Differentiation. Histochem. Cell Biol. 2016, 145, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Loo, L.; Simon, J.M.; Xing, L.; McCoy, E.S.; Niehaus, J.K.; Guo, J.; Anton, E.S.; Zylka, M.J. Single-Cell Transcriptomic Analysis of Mouse Neocortical Development. Nat. Commun. 2019, 10, 134. [Google Scholar] [CrossRef]
- Lee, D.R.; Rhodes, C.; Mitra, A.; Zhang, Y.; Maric, D.; Dale, R.K.; Petros, T.J. Transcriptional Heterogeneity of Ventricular Zone Cells in the Ganglionic Eminences of the Mouse Forebrain. eLife 2022, 11, e71864. [Google Scholar] [CrossRef] [PubMed]
- Siskos, N.; Ververidis, C.; Skavdis, G.; Grigoriou, M.E. Genoarchitectonic Compartmentalization of the Embryonic Telencephalon: Insights from the Domestic Cat. Front. Neuroanat. 2021, 15, 785541. [Google Scholar] [CrossRef] [PubMed]
- Knospe, C. Periods and stages of the prenatal development of the domestic cat. Anat. Histol. Embryol. 2002, 31, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Evans, H.E.; Sack, W.O. Prenatal development of domestic and laboratory mammals: Growth curves, external features and selected references. Anat. Histol. Embryol. 1973, 2, 11–45. [Google Scholar] [CrossRef] [PubMed]
- Glatzle, M.; Hoops, M.; Kauffold, J.; Seeger, J.; Fietz, S.A. Development of deep and upper neuronal layers in the domestic cat, sheep and pig neocortex. J. Vet. Med. Ser. C Anat. Histol. Embryol. 2017, 46, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Luskin, M.B.; Shatz, C.J. Neurogenesis of the cat’s primary visual cortex. J. Comp. Neurol. 1985, 242, 611–631. [Google Scholar] [CrossRef]
- Luskin, M.B.; Shatz, C.J. Studies of the earliest generated cells of the cat’s visual cortex: Cogeneration of subplate and marginal zones. J. Neurosci. 1985, 5, 1062–1075. [Google Scholar] [CrossRef]
- Workman, A.D.; Charvet, C.J.; Clancy, B.; Darlington, R.B.; Finlay, B.L. Modeling transformations of neurodevelopmental sequences across mammalian species. J. Neurosci. 2013, 33, 7368–7383. [Google Scholar] [CrossRef] [PubMed]
- Chytoudis-Peroudis, C.C.; Siskos, N.; Kalyviotis, K.; Fysekis, I.; Ypsilantis, P.; Simopoulos, C.; Skavdis, G.; Grigoriou, M.E. Spatial Distribution of the Full-Length Members of the Grg Family during Embryonic Neurogenesis Reveals a “Grg-Mediated Repression Map” in the Mouse Telencephalon. PLoS ONE 2018, 13, e0209369. [Google Scholar] [CrossRef]
- Lunde, A.; Glover, J.C. A Versatile Toolbox for Semi-Automatic Cell-by-Cell Object-Based Colocalization Analysis. Sci. Rep. 2020, 10, 19027. [Google Scholar] [CrossRef] [PubMed]
- Casarosa, S.; Fode, C.; Guillemot, F. Mash1 Regulates Neurogenesis in the Ventral Telencephalon. Development 1999, 126, 525–534. [Google Scholar] [CrossRef]
- Eisenstat, D.D.; Liu, J.K.; Mione, M.; Zhong, W.; Yu, G.; Anderson, S.A.; Ghattas, I.; Puelles, L.; Rubenstein, J.L. DLX-1, DLX-2, and DLX-5 Expression Define Distinct Stages of Basal Forebrain Differentiation. J. Comp. Neurol. 1999, 414, 217–237. [Google Scholar] [CrossRef]
- Lavdas, A.A.; Grigoriou, M.; Pachnis, V.; Parnavelas, J.G. The Medial Ganglionic Eminence Gives Rise to a Population of Early Neurons in the Developing Cerebral Cortex. J. Neurosci. 1999, 19, 7881–7888. [Google Scholar] [CrossRef] [PubMed]
- Liodis, P.; Denaxa, M.; Grigoriou, M.; Akufo-Addo, C.; Yanagawa, Y.; Pachnis, V. Lhx6 Activity Is Required for the Normal Migration and Specification of Cortical Interneuron Subtypes. J. Neurosci. 2007, 27, 3078–3089. [Google Scholar] [CrossRef] [PubMed]
- Menezes, J.R.; Luskin, M.B. Expression of Neuron-Specific Tubulin Defines a Novel Population in the Proliferative Layers of the Developing Telencephalon. J. Neurosci. 1994, 14, 5399–5416. [Google Scholar] [CrossRef] [PubMed]
- Molnár, Z.; Métin, C.; Stoykova, A.; Tarabykin, V.; Price, D.J.; Francis, F.; Meyer, G.; Dehay, C.; Kennedy, H. Comparative Aspects of Cerebral Cortical Development. Eur. J. Neurosci. 2006, 23, 921–934. [Google Scholar] [CrossRef]
- Guillemot, F. Cell Fate Specification in the Mammalian Telencephalon. Prog. Neurobiol. 2007, 83, 37–52. [Google Scholar] [CrossRef]
- Urban, N.; Guillemot, F. Neurogenesis in the Embryonic and Adult Brain: Same Regulators, Different Roles. Front. Cell. Neurosci. 2014, 8, 396. [Google Scholar] [CrossRef]
- Hardwick, L.J.A.; Philpott, A. Nervous Decision-Making: To Divide or Differentiate. Trends Genet. 2014, 30, 254–261. [Google Scholar] [CrossRef]
- Villalba, A.; Götz, M.; Borrell, V. The Regulation of Cortical Neurogenesis. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2021; Volume 142, pp. 1–66. ISBN 978-0-12-815281-2. [Google Scholar]
- Smart, I.H. A Pilot Study of Cell Production by the Ganglionic Eminences of the Developing Mouse Brain. J. Anat. 1976, 121, 71–84. [Google Scholar]
- Fentress, J.C.; Stanfield, B.B.; Cowan, W.M. Observations on the Development of the Striatum in Mice and Rats. Anat. Embryol. 1981, 163, 275–298. [Google Scholar] [CrossRef] [PubMed]
- Pilz, G.-A.; Shitamukai, A.; Reillo, I.; Pacary, E.; Schwausch, J.; Stahl, R.; Ninkovic, J.; Snippert, H.J.; Clevers, H.; Godinho, L.; et al. Amplification of Progenitors in the Mammalian Telencephalon Includes a New Radial Glial Cell Type. Nat. Commun. 2013, 4, 2125. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.E. Cell Cycle Regulation and Interneuron Production. Dev. Neurobiol. 2011, 71, 2–9. [Google Scholar] [CrossRef]
- Yun, K.; Potter, S.; Rubenstein, J.L.R. Gsh2 and Pax6 Play Complementary Roles in Dorsoventral Patterning of the Mammalian Telencephalon. Development 2001, 128, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Grigoriou, M.; Tucker, A.S.; Sharpe, P.T.; Pachnis, V. Expression and Regulation of Lhx6 and Lhx7, a Novel Subfamily of LIM Homeodomain Encoding Genes, Suggests a Role in Mammalian Head Development. Development 1998, 125, 2063–2074. [Google Scholar] [CrossRef]
- Katarova, Z.; Sekerková, G.; Prodan, S.; Mugnaini, E.; Szabó, G. Domain-Restricted Expression of Two Glutamic Acid Decarboxylase Genes in Midgestation Mouse Embryos. J. Comp. Neurol. 2000, 424, 607–627. [Google Scholar] [CrossRef]
- Turrero García, M.; Harwell, C.C. Radial Glia in the Ventral Telencephalon. FEBS Lett. 2017, 591, 3942–3959. [Google Scholar] [CrossRef]
- Mathieson, T.; Franken, H.; Kosinski, J.; Kurzawa, N.; Zinn, N.; Sweetman, G.; Poeckel, D.; Ratnu, V.S.; Schramm, M.; Becher, I.; et al. Systematic Analysis of Protein Turnover in Primary Cells. Nat. Commun. 2018, 9, 689. [Google Scholar] [CrossRef]
- Hasenpusch-Theil, K.; Theil, T. The Multifaceted Roles of Primary Cilia in the Development of the Cerebral Cortex. Front. Cell Dev. Biol. 2021, 9, 630161. [Google Scholar] [CrossRef]
- Doetsch, F.; Caillé, I.; Lim, D.A.; García-Verdugo, J.M.; Alvarez-Buylla, A. Subventricular Zone Astrocytes Are Neural Stem Cells in the Adult Mammalian Brain. Cell 1999, 97, 703–716. [Google Scholar] [CrossRef] [PubMed]
- Glickstein, S.B.; Moore, H.; Slowinska, B.; Racchumi, J.; Suh, M.; Chuhma, N.; Ross, M.E. Selective Cortical Interneuron and GABA Deficits in Cyclin D2-Null Mice. Development 2007, 134, 4083–4093. [Google Scholar] [CrossRef]
- Glickstein, S.B.; Monaghan, J.A.; Koeller, H.B.; Jones, T.K.; Ross, M.E. Cyclin D2 Is Critical for Intermediate Progenitor Cell Proliferation in the Embryonic Cortex. J. Neurosci. 2009, 29, 9614–9624. [Google Scholar] [CrossRef]
- Noctor, S.C.; Martínez-Cerdeño, V.; Ivic, L.; Kriegstein, A.R. Cortical Neurons Arise in Symmetric and Asymmetric Division Zones and Migrate through Specific Phases. Nat. Neurosci. 2004, 7, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Martynoga, B.; Drechsel, D.; Guillemot, F. Molecular Control of Neurogenesis: A View from the Mammalian Cerebral Cortex. Cold Spring Harb. Perspect. Biol. 2012, 4, a008359. [Google Scholar] [CrossRef]
- Stoufflet, J.; Caillé, I. The Primary Cilium and Neuronal Migration. Cells 2022, 11, 3384. [Google Scholar] [CrossRef] [PubMed]
- Spaulding, E.L.; Burgess, R.W. Accumulating Evidence for Axonal Translation in Neuronal Homeostasis. Front. Neurosci. 2017, 11, 312. [Google Scholar] [CrossRef]
- Glock, C.; Biever, A.; Tushev, G.; Nassim-Assir, B.; Kao, A.; Bartnik, I.; Tom Dieck, S.; Schuman, E.M. The Translatome of Neuronal Cell Bodies, Dendrites, and Axons. Proc. Natl. Acad. Sci. USA 2021, 118, e2113929118. [Google Scholar] [CrossRef] [PubMed]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibody | Company | Catalog Number | Dilution |
|---|---|---|---|
| Rabbit anti-Abracl | [11] | - | 1/100 |
| Rabbit anti-Abracl | ATLAS Antibodies | HPA030217 | 1/100 |
| Rat anti-Ki67 | Invitrogen | 14-5698-80 | 1/500 |
| Mouse anti-Tubb3 | Millipore | MAB1637 | 1/100 (1/1000 in cells) |
| Mouse anti-Dlx2 | Santa Cruz antibodies | sc-393879 | 1/100 |
| Mouse anti-Ascl1 | Santa Cruz antibodies | sc-374104 | 1/100 |
| Mouse anti-Gad65/67 | Santa Cruz antibodies | sc-365180 | 1/100 |
| Mouse anti-Tbr1 | Proteintech | 66564-1-Ig | 1/100 |
| Mouse anti-Lhx6 | Santa Cruz antibodies | sc-271433 | 1/100 |
| Secondary Antibody | Company | Catalog Number | Dilution |
|---|---|---|---|
| Goat anti-rabbit IgG CF488A | Biotium | 20015 | 1/2000 |
| Donkey anti-rat IgG CF568 | Biotium | 20092-1 | 1/500 |
| Donkey anti-mouse IgG CF568 | Biotium | 20105 | 1/500 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Troumpoukis, D.; Vasileiou, A.R.; Siskos, N.; Stylianopoulou, E.; Ypsilantis, P.; Skavdis, G.; Grigoriou, M.E. Characterization of the Abracl-Expressing Cell Populations in the Embryonic Mammalian Telencephalon. Biomolecules 2023, 13, 1337. https://doi.org/10.3390/biom13091337
Troumpoukis D, Vasileiou AR, Siskos N, Stylianopoulou E, Ypsilantis P, Skavdis G, Grigoriou ME. Characterization of the Abracl-Expressing Cell Populations in the Embryonic Mammalian Telencephalon. Biomolecules. 2023; 13(9):1337. https://doi.org/10.3390/biom13091337
Chicago/Turabian StyleTroumpoukis, Dimitrios, Andreas Rafail Vasileiou, Nikistratos Siskos, Electra Stylianopoulou, Petros Ypsilantis, George Skavdis, and Maria E. Grigoriou. 2023. "Characterization of the Abracl-Expressing Cell Populations in the Embryonic Mammalian Telencephalon" Biomolecules 13, no. 9: 1337. https://doi.org/10.3390/biom13091337
APA StyleTroumpoukis, D., Vasileiou, A. R., Siskos, N., Stylianopoulou, E., Ypsilantis, P., Skavdis, G., & Grigoriou, M. E. (2023). Characterization of the Abracl-Expressing Cell Populations in the Embryonic Mammalian Telencephalon. Biomolecules, 13(9), 1337. https://doi.org/10.3390/biom13091337