Zika and Flavivirus Shell Disorder: Virulence and Fetal Morbidity




Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. The Correlation between Shell Disorder in Flaviviruses and Zika and Virulence and Fetal Morbidity
3.2. C and M Proteins Play Roles in Virulence and Morbidity
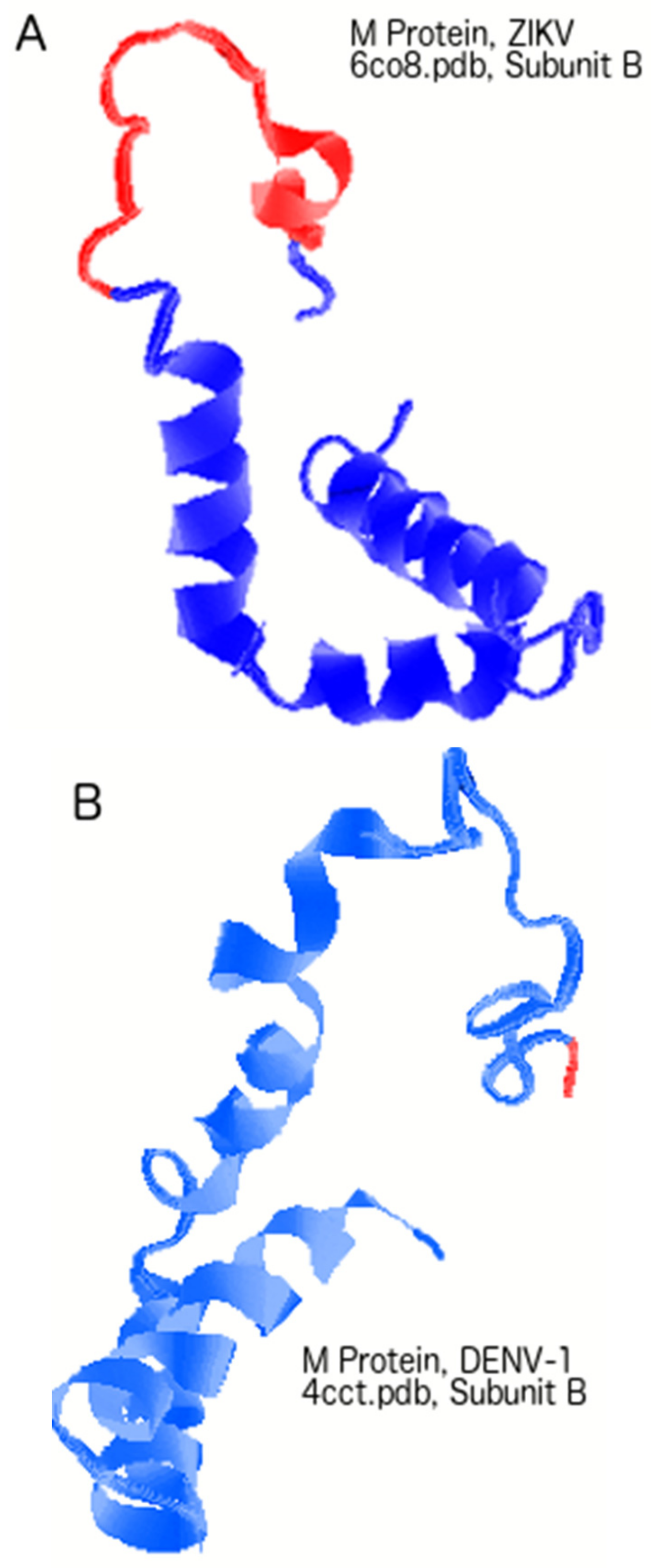
3.3. Regions of Disorder in the M Protein: ZIKV versus YFV and DENV-2
3.4. PONDR VLXT Plot for C Proteins from ZIKV and Other Flaviviruses
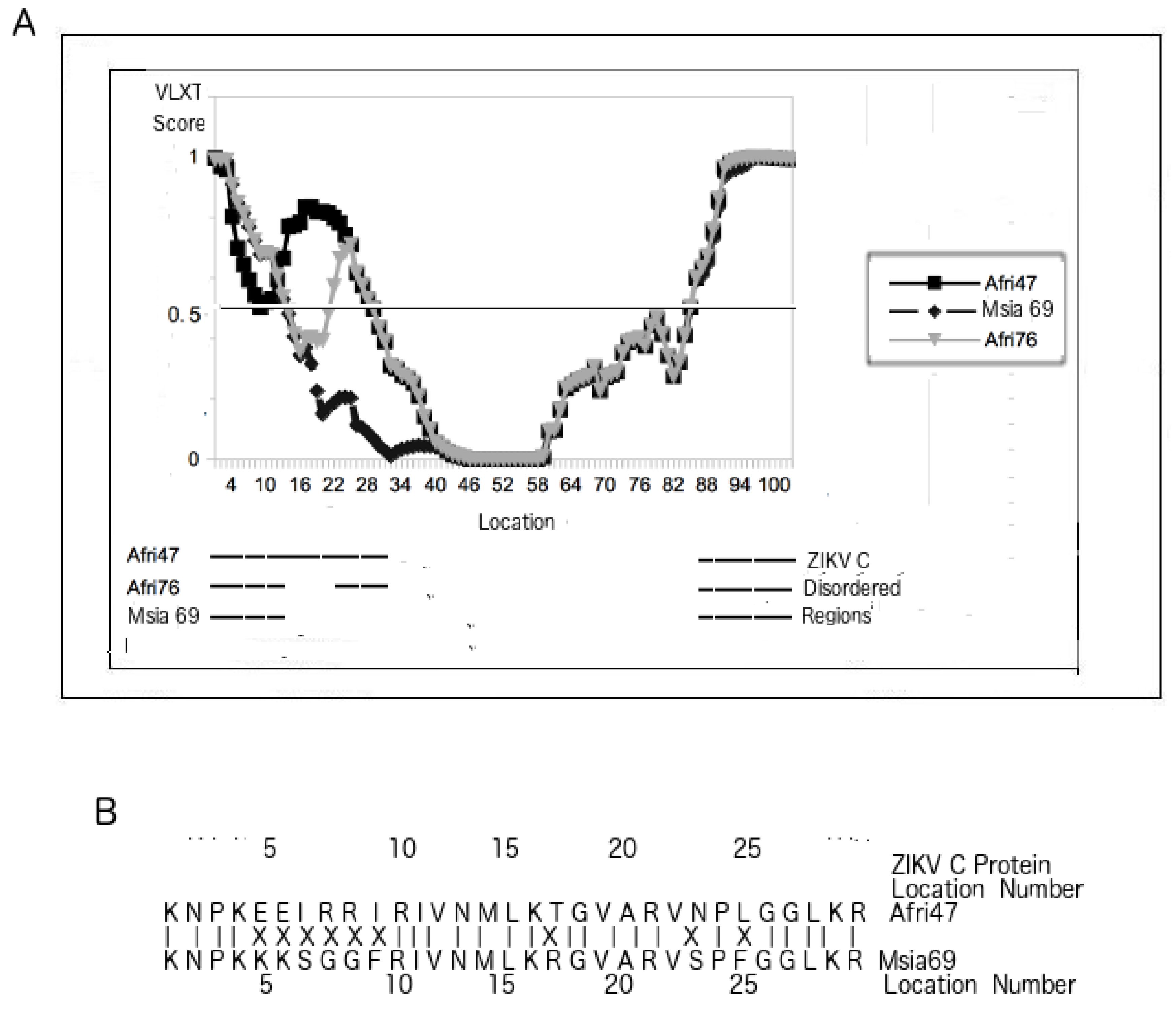
3.5. PONDR VLXT Plot for C Proteins from Different ZIKV Strains
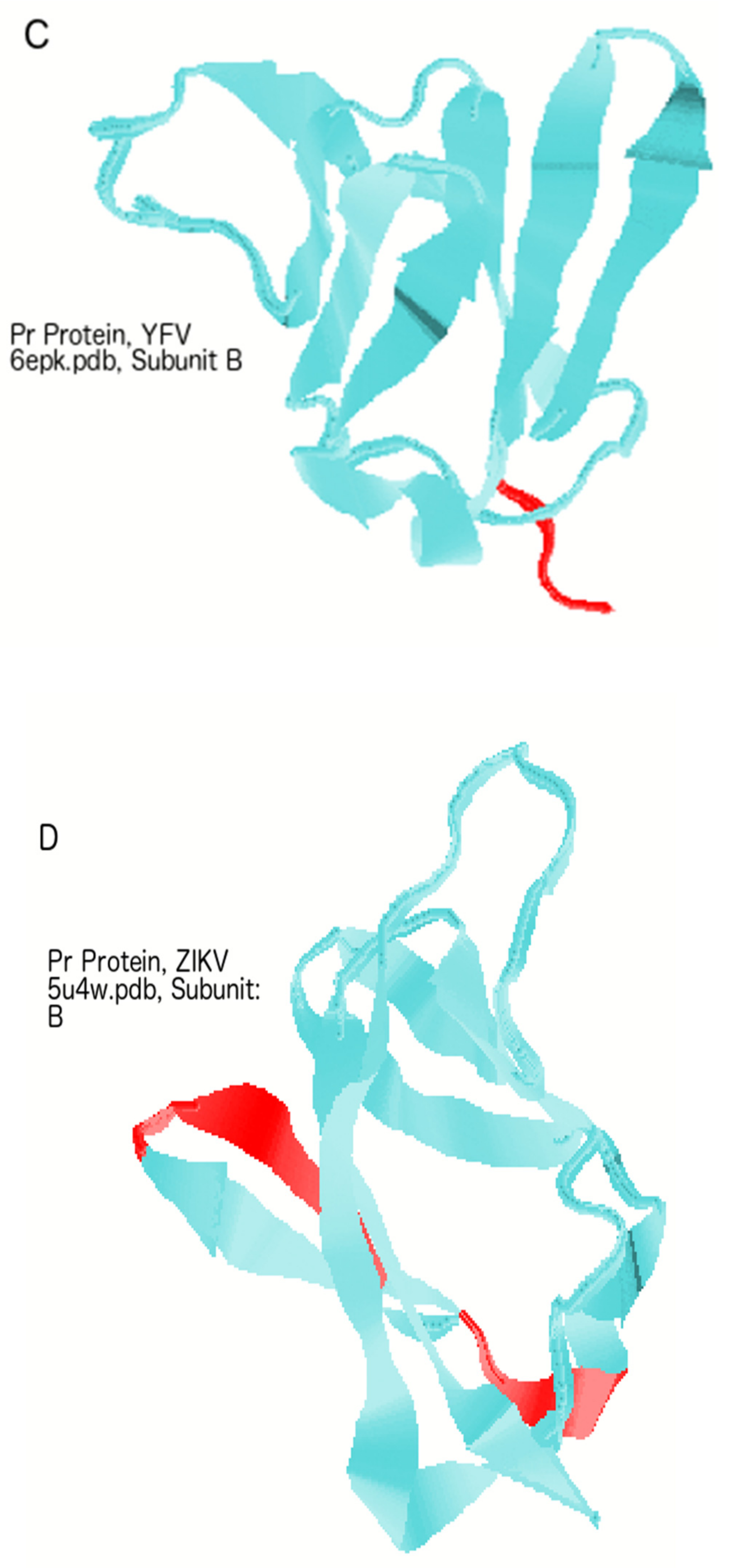
3.6. The Roles of M Versus PrM and Pr in Protecting the Virion at Various Stages
4. Discussion
4.1. Virulence and Disorder from Inner and Outer Shells: Clues from Other Viruses
4.2. A Lesson from HIV: Disordered Outer Shell Provides Better Viral Penetration of Vital Organs
4.3. Virulence and Inner Shell Disorder Involve a Different “Trojan Horse” Strategy in Immune Evasion
4.4. Taking Turns to Protect the Virion from Environmental Insults
4.5. Conflicting Trends Can Be Found among Flaviviruses: Protecting the Virion Versus Immune Evasion
4.6. YFV Uses both Inner and Outer Shell Disorder to Evade Immune Systems, Resulting in High Pathogenicity
4.7. The Shell Disorder Model Predicts Correctly that ZIKV Is Able to Penetrate Vital Organs with Greater Efficiency
4.8. ZIKV C PID Variation Correlates with Fetal Morbidity: A “Trojan Horse”
4.9. Evolution of ZIKV Variation in Fetal Morbidity Is Dependent on the Optimal Viral Load Necessary for More Efficient Transmission Among Its Primary Primate Hosts
4.10. Experimental Evidence for the Role of C and M Proteins in Replication and Immune Evasion: Roles for Disorder
4.11. Clues for Developing a ZIKV Vaccine
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Song, B.-H.; Yun, S.-I.; Woolley, M.; Lee, Y.-M. Zika virus: History, epidemiology, transmission, and clinical presentation. J. Neuroimmunol. 2017, 308, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Goh, G.K. Viral Shapeshifters: Strange Behaviors of HIV and Other Viruses; Simplicity Research Institute: Singapore, 2017. [Google Scholar]
- Petersen, L.R.; Jamieson, D.J.; Powers, A.M.; Honein, M.A. Zika Virus. N. Engl. J. Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Heukelbach, J.; Alencar, C.H.; Kelvin, A.A.; De Oliveira, W.K.; Cavalcanti, L.P.D.G. Zika virus outbreak in Brazil. J. Infect. Dev. Ctries. 2016, 10, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-L.; Tang, R.-B. Why Zika virus infection has become a public health concern? J. Chin. Med. Assoc. 2016, 79, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Hafiz, M.Y.; Mahmood, S.U.; Shoaib, M.; Yusuf, F.H. Concern over Zika virus outbreak: Another alarming global threat. Infect. Drug Resist. 2016, 9, 149–151. [Google Scholar] [CrossRef]
- Udenze, D.; Trus, I.; Berube, N.; Gerdts, V.; Karniychuk, U. The African strain of Zika virus causes more severe in utero infection than Asian strain in a porcine fetal transmission model. Emerg. Microbes Infect. 2019, 8, 1098–1107. [Google Scholar] [CrossRef]
- Gurung, S.; Reuter, N.; Preno, A.; Dubaut, J.; Nadeau, H.; Hyatt, K.; Singleton, K.; Martin, A.; Parks, W.T.; Papin, J.F.; et al. Zika virus infection at mid-gestation results in fetal cerebral cortical injury and fetal death in the olive baboon. PLoS Pathog. 2019, 15, e1007507. [Google Scholar] [CrossRef]
- Pena, L.J.; Guarines, K.M.; Silva, A.J.D.; Leal, L.R.S.; Félix, D.M.; Silva, A.; De Oliveira, S.A.; Ayres, C.F.J.; Júnior, A.S.; De Freitas, A.C. In vitro and in vivo models for studying Zika virus biology. J. Gen. Virol. 2018, 99, 1529–1550. [Google Scholar] [CrossRef]
- Gorman, M.J.; Caine, E.A.; Zaitsev, K.; Begley, M.C.; Weger-Lucarelli, J.; Uccellini, M.B.; Tripathi, S.; Morrison, J.; Yount, B.L.; Dinnon, K.H., 3rd; et al. An Immunocompetent Mouse Model of Zika Virus Infection. Cell Host Microbe 2018, 23, 672–685.e6. [Google Scholar] [CrossRef] [Green Version]
- Schreur, P.J.W.; Van Keulen, L.; Anjema, D.; Kant, J.; Kortekaas, J. Microencephaly in fetal piglets following in utero inoculation of Zika virus. Emerg. Microbes Infect. 2018, 7, 1–11. [Google Scholar] [CrossRef]
- Hirsch, A.J.; Roberts, V.H.J.; Grigsby, P.L.; Haese, N.; Schabel, M.C.; Wang, X.; Lo, J.O.; Liu, Z.; Kroenke, C.D.; Smith, J.L.; et al. Zika virus infection in pregnant rhesus macaques causes placental dysfunction and immunopathology. Nat. Commun. 2018, 9, 263. [Google Scholar] [CrossRef] [PubMed]
- Winkler, C.W.; Peterson, K.E. Using immunocompromised mice to identify mechanisms of Zika virus transmission and pathogenesis. Immunology 2018, 153, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, G.K.-M.; Dunker, A.K.; Foster, J.A.; Uversky, V.N. HIV Vaccine Mystery and Viral Shell Disorder. Biomolecules 2019, 9, 178. [Google Scholar] [CrossRef] [PubMed]
- Acheson, N.H. Fundamentals of Molecular Virology; Wiley: Hoboken, NJ, USA, 2007. [Google Scholar]
- Sirohi, D.; Kuhn, R.J. Zika Virus Structure, Maturation, and Receptors. J. Infect. Dis. 2017, 216, S935–S944. [Google Scholar] [CrossRef] [Green Version]
- Prasad, V.M.; Miller, A.S.; Klose, T.; Sirohi, D.; Buda, G.; Jiang, W.; Kuhn, R.J.; Rossmann, M.G. Structure of the immature Zika virus at 9 Å resolution. Nat. Struct. Mol. Boil. 2017, 24, 184–186. [Google Scholar] [CrossRef]
- Dunker, A.K.; Brown, C.J.; Lawson, J.D.; Iakoucheva, L.M.; Obradović, Z. Intrinsic Disorder and Protein Function†. Biochemistry 2002, 41, 6573–6582. [Google Scholar] [CrossRef]
- Wright, P.E.; Dyson, H.J. Intrinsically unstructured proteins: Re-assessing the protein structure-function paradigm. J. Mol. Boil. 1999, 293, 321–331. [Google Scholar] [CrossRef]
- Uversky, V.N.; Gillespie, J.R.; Fink, A.L. Why are “natively unfolded” proteins unstructured under physiologic conditions? Proteins: Struct. Funct. Bioinform. 2000, 41, 415–427. [Google Scholar] [CrossRef]
- Dunker, A.K.; Lawson, J.D.; Brown, C.J.; Williams, R.M.; Romero, P.; Oh, J.S.; Oldfield, C.J.; Campen, A.M.; Ratliff, C.M.; Hipps, K.W.; et al. Intrinsically disordered protein. J. Mol. Graph. Model. 2001, 19, 26–59. [Google Scholar] [CrossRef] [Green Version]
- Tompa, P. Intrinsically unstructured proteins. Trends Biochem. Sci. 2002, 27, 527–533. [Google Scholar] [CrossRef]
- Giri, R.; Kumar, D.; Sharma, N.; Uversky, V.N. Intrinsically Disordered Side of the Zika Virus Proteome. Front. Microbiol. 2016, 6, 144. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.M.; Uversky, V.N.; Giri, R. Molecular Recognition Features in Zika Virus Proteome. J. Mol. Boil. 2018, 430, 2372–2388. [Google Scholar] [CrossRef] [PubMed]
- Goh, G.K.-M.; Dunker, A.K.; Uversky, V.N. Correlating Flavivirus virulence and levels of intrinsic disorder in shell proteins: Protective roles vs. immune evasion. Mol. BioSyst. 2016, 12, 1881–1891. [Google Scholar] [CrossRef] [PubMed]
- Xue, B.; Blocquel, D.; Habchi, J.; Uversky, A.V.; Kurgan, L.; Uversky, V.N.; Longhi, S. Structural Disorder in Viral Proteins. Chem. Rev. 2014, 114, 6880–6911. [Google Scholar] [CrossRef] [PubMed]
- Xue, B.; Dunker, A.K.; Uversky, V.N. Orderly order in protein intrinsic disorder distribution: Disorder in 3500 proteomes from viruses and the three domains of life. J. Biomol. Struct. Dyn. 2012, 30, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Xue, B.; Ganti, K.; Rabionet, A.; Banks, L.; Uversky, V.N. Disordered interactome of human papillomavirus. Curr. Pharm. Des. 2014, 20, 1274–1292. [Google Scholar] [CrossRef] [PubMed]
- Uversky, V.N.; Roman, A.; Oldfield, C.J.; Dunker, A.K. Protein Intrinsic Disorder and Human Papillomaviruses: Increased Amount of Disorder in E6 and E7 Oncoproteins from High Risk HPVs. J. Proteome Res. 2006, 5, 1829–1842. [Google Scholar] [CrossRef]
- Goh, G.K.-M.; Dunker, A.K.; Uversky, V.N. Protein intrinsic disorder and influenza virulence: The 1918 H1N1 and H5N1 viruses. Virol. J. 2009, 6, 69. [Google Scholar] [CrossRef]
- Xue, B.; Mizianty, M.J.; Kurgan, L.; Uversky, V.N. Protein intrinsic disorder as a flexible armor and a weapon of HIV-1. Cell Mol. Life Sci. 2012, 69, 1211–1259. [Google Scholar] [CrossRef]
- Dolan, P.T.; Roth, A.P.; Xue, B.; Sun, R.; Dunker, A.K.; Uversky, V.N.; LaCount, D.J. Intrinsic disorder mediates hepatitis C virus core-host cell protein interactions. Protein Sci. 2015, 24, 221–235. [Google Scholar] [CrossRef]
- Fan, X.; Xue, B.; Dolan, P.T.; LaCount, D.J.; Kurgan, L.; Uversky, V.N. The intrinsic disorder status of the human hepatitis C virus proteome. Mol. BioSyst. 2014, 10, 1345–1363. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Badierah, R.A.; Almehdar, H.A.; Redwan, E.M.; Kurgan, L.; Uversky, V.N. Unstructural biology of the dengue virus proteins. FEBS J. 2015, 282, 3368–3394. [Google Scholar] [CrossRef] [PubMed]
- Whelan, J.N.; Reddy, K.D.; Uversky, V.N.; Teng, M.N. Functional correlations of respiratory syncytial virus proteins to intrinsic disorder. Mol. BioSyst. 2016, 12, 1507–1526. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kumar, A.; Yadav, R.; Uversky, V.N.; Giri, R. Deciphering the dark proteome of Chikungunya virus. Sci. Rep. 2018, 8, 5822. [Google Scholar] [CrossRef]
- Redwan, E.M.; AlJaddawi, A.A.; Uversky, V.N. Structural disorder in the proteome and interactome of Alkhurma virus (ALKV). Cell Mol. Life Sci. 2019, 76, 577–608. [Google Scholar] [CrossRef]
- Shao, Q.; Herrlinger, S.; Zhu, Y.-N.; Yang, M.; Goodfellow, F.; Stice, S.L.; Qi, X.-P.; Brindley, M.A.; Chen, J.-F. The African Zika virus MR-766 is more virulent and causes more severe brain damage than current Asian lineage and dengue virus. Development 2017, 144, 4114–4124. [Google Scholar] [CrossRef]
- Beaver, J.T.; Lelutiu, N.; Habib, R.; Skountzou, I. Evolution of Two Major Zika Virus Lineages: Implications for Pathology, Immune Response, and Vaccine Development. Front. Immunol. 2018, 9, 9. [Google Scholar] [CrossRef]
- Goh, G.K.-M.; Dunker, A.K.; Uversky, V.N. Protein intrinsic disorder toolbox for comparative analysis of viral proteins. BMC Genom. 2008, 9, S4. [Google Scholar] [CrossRef]
- Garner, E.; Romero, P.; Dunker, A.K.; Brown, C.; Obradovic, Z. Predicting Binding Regions within Disordered Proteins. Genome Inform Ser Workshop Genome Inform. 1999, 10, 41–50. [Google Scholar]
- Romero, P.; Obradovic, Z.; Kissinger, C.; Villafranca, J.E.; Dunker, A.K. Identifying disordered regions in proteins from amino acid sequence. In Proceedings of the IEEE International Conference on Neural Networks, Houston, TX, USA, 12 June 1997; pp. 90–95. [Google Scholar]
- Cheng, Y.; Oldfield, C.J.; Meng, J.; Romero, P.; Uversky, V.N.; Dunker, A.K. Mining α-Helix-Forming Molecular Recognition Features with Cross Species Sequence Alignments†. Biochemistry 2007, 46, 13468–13477. [Google Scholar] [CrossRef]
- Oldfield, C.J.; Cheng, Y.; Cortese, M.S.; Romero, P.; Uversky, V.N.; Dunker, A.K. Coupled Folding and Binding with α-Helix-Forming Molecular Recognition Elements†. Biochemistry 2005, 44, 12454–12470. [Google Scholar] [CrossRef] [PubMed]
- Goh, G.K.; Foster, J.A.; Dunker, A.K.; Uversky, V.N. Nipah virulence and v shell disorder. In SCON Summit on Vaccine and HIVAIDS Conferences, Las Vegas, NVUSA; 12 November 2019; in press. [Google Scholar]
- Goh, G.K.-M.; Dunker, A.K.; Uversky, V.N. A comparative analysis of viral matrix proteins using disorder predictors. Virol. J. 2008, 5, 126. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, A.A.; Heinemann, M.B.; Leite, R.C.; Reis, J.K.P. A comparative analysis of envelope and tegument proteins of suid herpesvirus 1, bovine herpesvirus 1 and bovine herpesvirus 5. Arch. Virol. 2010, 155, 1687–1692. [Google Scholar] [CrossRef] [PubMed]
- Xue, B.; Williams, R.W.; Oldfield, C.J.; Goh, G.K.-M.; Dunker, A.K.; Uversky, V.N. Viral disorder or disordered viruses: Do viral proteins possess unique features? Protein Pept. Lett. 2010, 17, 932–951. [Google Scholar] [CrossRef] [PubMed]
- Goh, G.K.-M.; Dunker, A.K.; Uversky, V.N. Understanding Viral Transmission Behavior via Protein Intrinsic Disorder Prediction: Coronaviruses. J. Pathog. 2012, 2012, 1–13. [Google Scholar] [CrossRef]
- Goh, G.K.-M.; Dunker, A.K.; Uversky, V. Prediction of Intrinsic Disorder in MERS-CoV/HCoV-EMC Supports a High Oral-Fecal Transmission. PLoS Curr. 2013, 5, 5. [Google Scholar] [CrossRef]
- Goh, G.K.-M.; Dunker, A.K.; Uversky, V.N. Detection of links between Ebola nucleocapsid and virulence using disorder analysis. Mol. BioSyst. 2015, 11, 2337–2344. [Google Scholar] [CrossRef]
- Goh, G.K.-M.; Dunker, A.K.; Uversky, V.N. Shell disorder, immune evasion and transmission behaviors among human and animal retroviruses. Mol. BioSyst. 2015, 11, 2312–2323. [Google Scholar] [CrossRef]
- Romero, P.; Obradovic, Z.; Li, X.; Garner, E.C.; Brown, C.J.; Dunker, A.K. Sequence complexity of disordered protein. Proteins Struct. Funct. Bioinform. 2001, 42, 38–48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; ISBN 3-900051-07-0. [Google Scholar]
- Zhang, Z.; Sun, M.; Deng, J.; Yu, J.; Yang, X.; Zhao, W.; Chen, G.; Wang, P. Zika Virus Induced More Severe Inflammatory Response Than Dengue Virus in Chicken Embryonic Livers. Front. Microbiol. 2019, 10, 1127. [Google Scholar] [CrossRef]
- Gong, Z.; Xu, X.; Han, G.-Z. The Diversification of Zika Virus: Are There Two Distinct Lineages? Genome Boil. Evol. 2017, 9, 2940–2945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Quijano, F.A.; Waldman, E.A. Factors Associated with Dengue Mortality in Latin America and the Caribbean, 1995–2009: An Ecological Study. Am. J. Trop. Med. Hyg. 2012, 86, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Halstead, S.B. Flavivirus vaccine. In Molecular Virology and Control of Flavivirus; Shi, P., Ed.; Caister: Wymanham, UK, 2012; p. 185. [Google Scholar]
- Mickiene, A.; Lindquist, L.; Laiskonis, A. Tick-borne encephalitis—Clinical course and outcome. In Progress in Encephalitis Research; Ebert, R.A., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2005; pp. 1–30. [Google Scholar]
- McKenzie, J.; Pinger, R.R.; Kotecki, J. Epidemiology: The study of disease, injury and death in the community. In An Introduction to Community Health, 8th ed.; Jones and Bartlett Publishers: Sudbury MA, USA, 2008; p. 70. [Google Scholar]
- Rodenhuis-Zybert, I.A.; Van Der Schaar, H.M.; Voorham, J.M.D.S.; Van Der Ende-Metselaar, H.; Lei, H.-Y.; Wilschut, J.; Smit, J.M. Immature Dengue Virus: A Veiled Pathogen? PLoS Pathog. 2010, 6, e1000718. [Google Scholar] [CrossRef] [PubMed]
- Zybert, I.A.; Van Der Ende-Metselaar, H.; Wilschut, J.; Smit, J.M. Functional importance of dengue virus maturation: Infectious properties of immature virions. J. Gen. Virol. 2008, 89, 3047–3051. [Google Scholar] [CrossRef] [PubMed]
- Goudsmit, J. Viral Sex: The Nature of AIDS; Oxford University Press: New York, NY, USA, 1997. [Google Scholar]
- Bhuvanakantham, R.; Chong, M.-K.; Ng, M.-L. Specific interaction of capsid protein and importin-α/β influences West Nile virus production. Biochem. Biophys. Res. Commun. 2009, 389, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Colpitts, T.M.; Barthel, S.; Wang, P.; Fikrig, E. Dengue Virus Capsid Protein Binds Core Histones and Inhibits Nucleosome Formation in Human Liver Cells. PLoS ONE 2011, 6, e24365. [Google Scholar] [CrossRef]
- Bhuvanakantham, R.; Ng, M.-L. West Nile virus and dengue virus capsid protein negates the antiviral activity of human Sec3 protein through the proteasome pathway. Cell. Microbiol. 2013, 15, 1688–1706. [Google Scholar] [CrossRef]
- Li, L.; Lok, S.-M.; Yu, I.-M.; Zhang, Y.; Kuhn, R.J.; Chen, J.; Rossmann, M.G. The Flavivirus Precursor Membrane-Envelope Protein Complex: Structure and Maturation. Science 2008, 319, 1830–1834. [Google Scholar] [CrossRef] [Green Version]
- Wei, D.; Xu, J.; Liu, X.X.-Y.; Chen, Z.-N.; Bian, H. Fighting Cancer with Viruses: Oncolytic Virus Therapy in China. Hum. Gene Ther. 2018, 29, 151–159. [Google Scholar] [CrossRef]
- Chernajovsky, Y.; Layward, L.; Lemoine, N. Fighting cancer with oncolytic viruses. BMJ 2006, 332, 170–172. [Google Scholar] [CrossRef] [Green Version]
- Chaurasiya, S.; Chen, N.G.; Warner, S.G. Oncolytic Virotherapy versus Cancer Stem Cells: A Review of Approaches and Mechanisms. Cancers 2018, 10, 124. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Lineage | C PID d | M PID d | UniProt ID |
|---|---|---|---|---|
| Senegal 2001 Uganda 1947 (MR-766) a,b | African strains | 45.0 ± 0.1 | 28.0 ± 0.1 | W8QFH2 Q32ZE1 a,b |
| Uganda 1947 (MR-766) a,c Central African Rep 1976 | African strains | 37.0 ± 0.1 | 28.0 ± 0.1 | A0A2S1KKZ9 a,c W0G5P7 |
| Malaysia 1969 Colombia 2015 Brazil 2015 | Asian strains | 31.0 ± 0.1 | 29.0 ± 0.1 | H8XX11 A0A127AM58 A0A0X9QZM7 |
| Micronesia 2007 | Asian strain | 36.0 ± 0.1 | 29.0 ± 0.1 | B3U3M3 |
| Virus | CFR a,b,c | C PID a,b | M PID a,c | UniProt ID |
|---|---|---|---|---|
| YFV | >50% | 74 ± 1d | 35 ± 1d | P03314, Q6DV88, Q1X881 |
| TBEV-Fe | 20–40% | 68 ± 3 | 8.0 ± 0.1 | G5CP55, P07720, K4P8A |
| TBEV-Si | 2–3% | 63 ± 1 | 6.0 ± 0.1 | G8FGD9, G8FGD9 |
| TBEV-Eu | 1–2% | 62 ± 2 | 6.0 ± 0.1 | Q01299, P14336 |
| DENV1-4 | 1.4 ± 0.8% | 47 ± 5 | 11 ± 5 | P14337, P09866, Q58Ht7 |
| WNV | ~0.4% | 49 ± 4 | 9.0 ± 0.1 | Q9EMB5, Q53AP1, P06935 |
| ZIKV | <0.2% | 35 ± 6 | 28.7 ± 0.5 | P14336, Q32ZE1, H8XX11 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goh, G.K.-M.; Dunker, A.K.; Foster, J.A.; Uversky, V.N. Zika and Flavivirus Shell Disorder: Virulence and Fetal Morbidity. Biomolecules 2019, 9, 710. https://doi.org/10.3390/biom9110710
Goh GK-M, Dunker AK, Foster JA, Uversky VN. Zika and Flavivirus Shell Disorder: Virulence and Fetal Morbidity. Biomolecules. 2019; 9(11):710. https://doi.org/10.3390/biom9110710
Chicago/Turabian StyleGoh, Gerard Kian-Meng, A. Keith Dunker, James A. Foster, and Vladimir N. Uversky. 2019. "Zika and Flavivirus Shell Disorder: Virulence and Fetal Morbidity" Biomolecules 9, no. 11: 710. https://doi.org/10.3390/biom9110710
APA StyleGoh, G. K. -M., Dunker, A. K., Foster, J. A., & Uversky, V. N. (2019). Zika and Flavivirus Shell Disorder: Virulence and Fetal Morbidity. Biomolecules, 9(11), 710. https://doi.org/10.3390/biom9110710