Predicting Angiogenesis by Endothelial Progenitor Cells Relying on In-Vitro Function Assays and VEGFR-2 Expression Levels

Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Culturing of Endothelial Progenitor Cells from Human Peripheral Blood
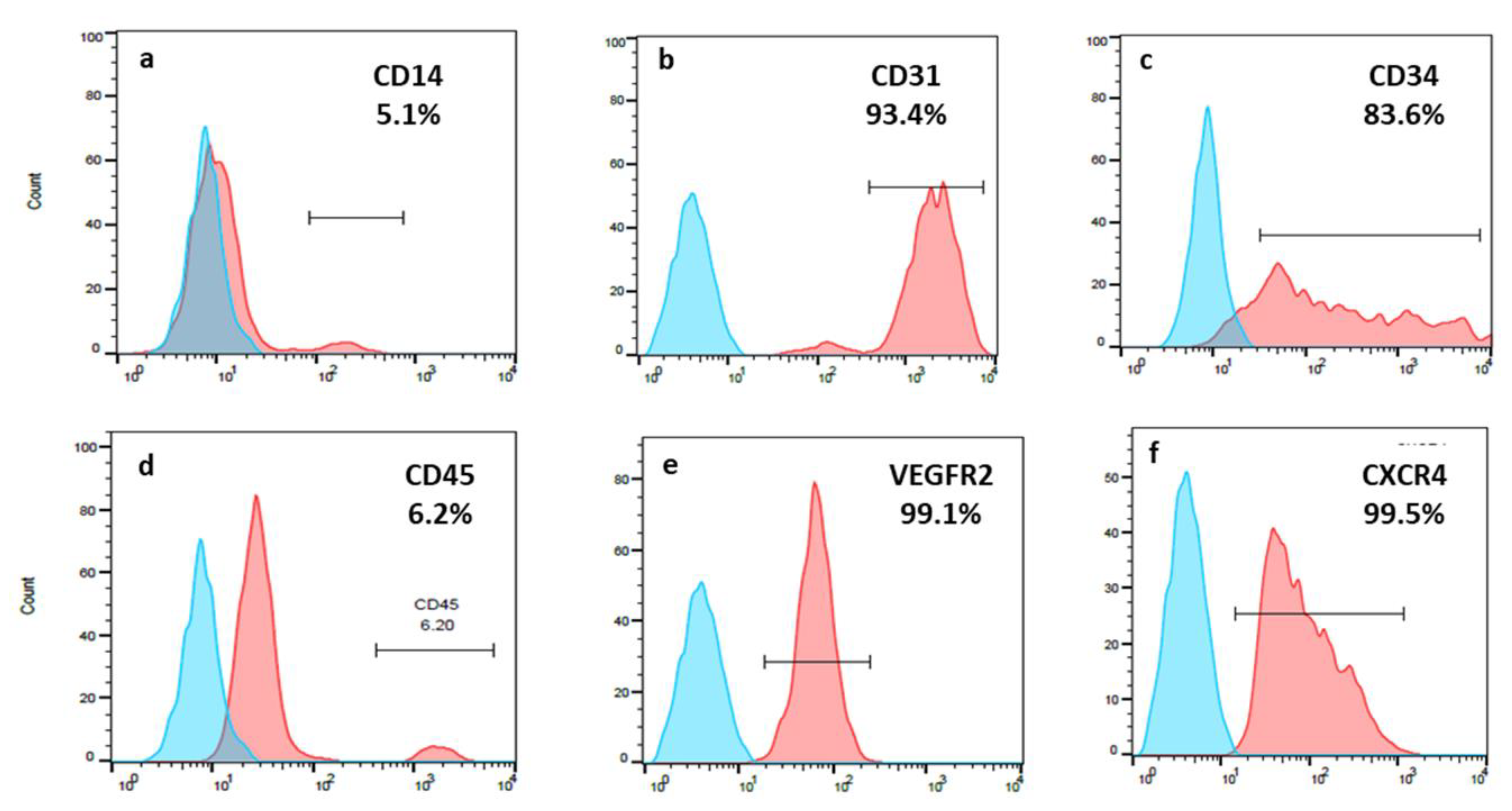
2.2. Characterization of Human EPCs
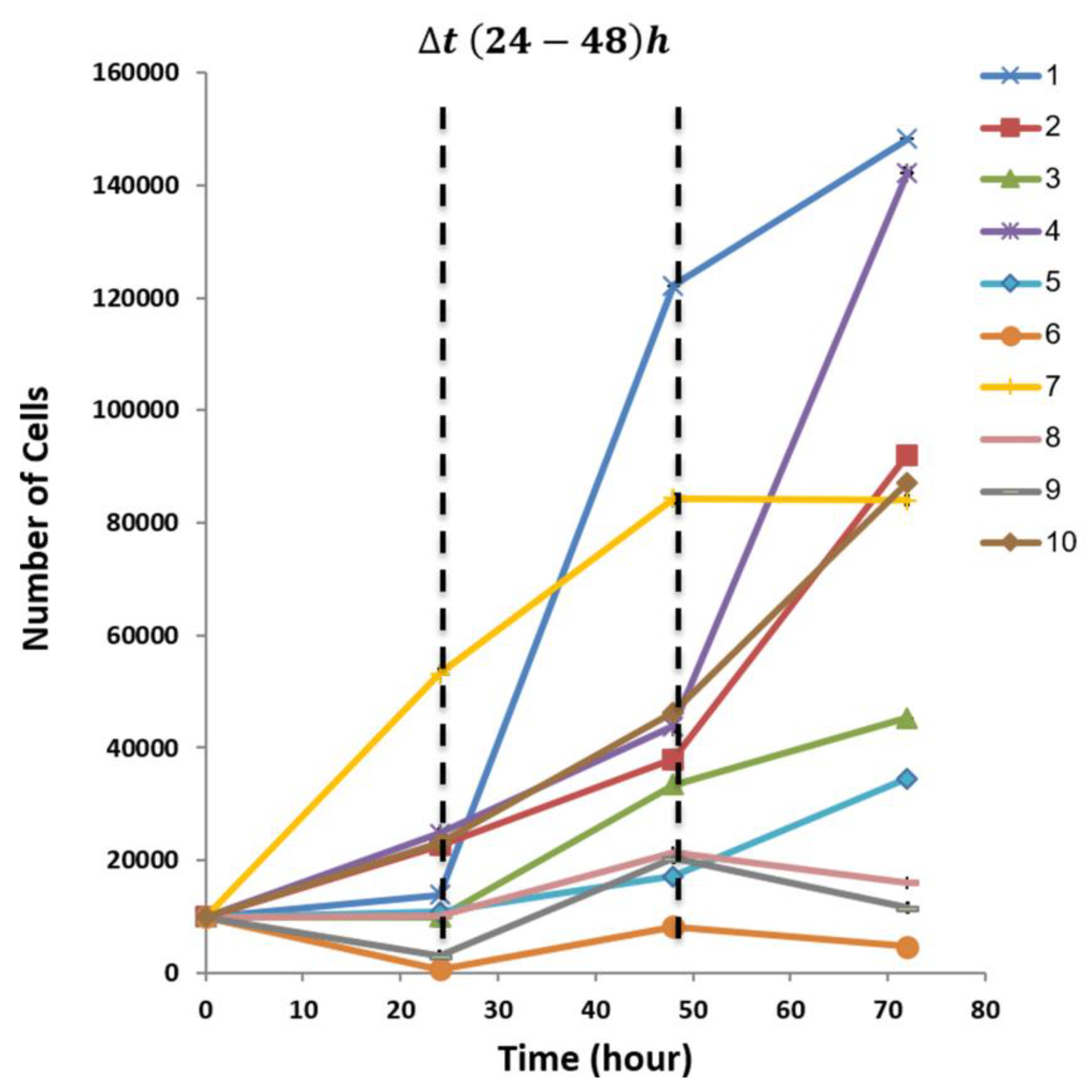
2.3. Cell Proliferation Assay XTT
2.4. EPC Conditioned Medium (EPC-CM) Preparation
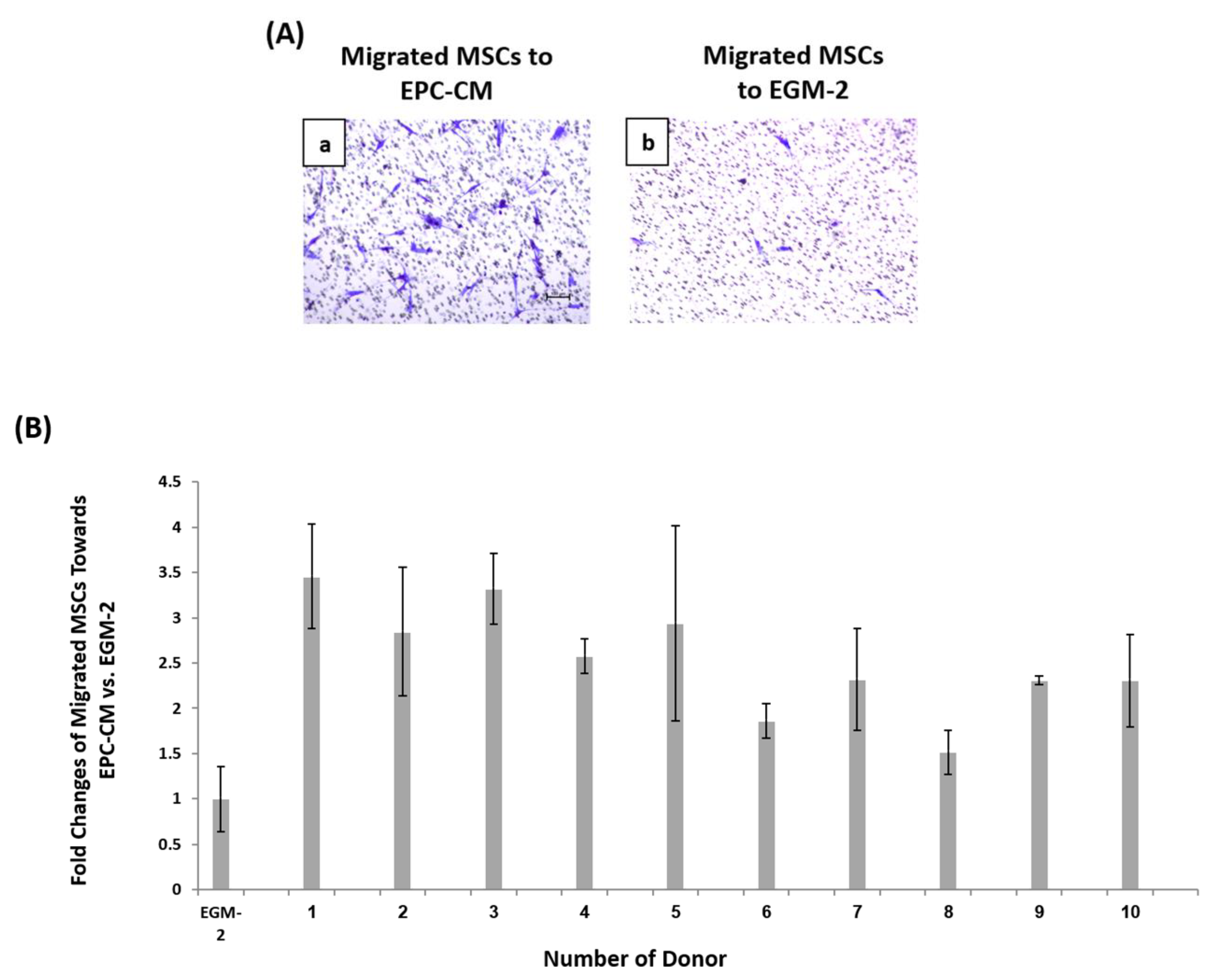
2.5. Migration Assay
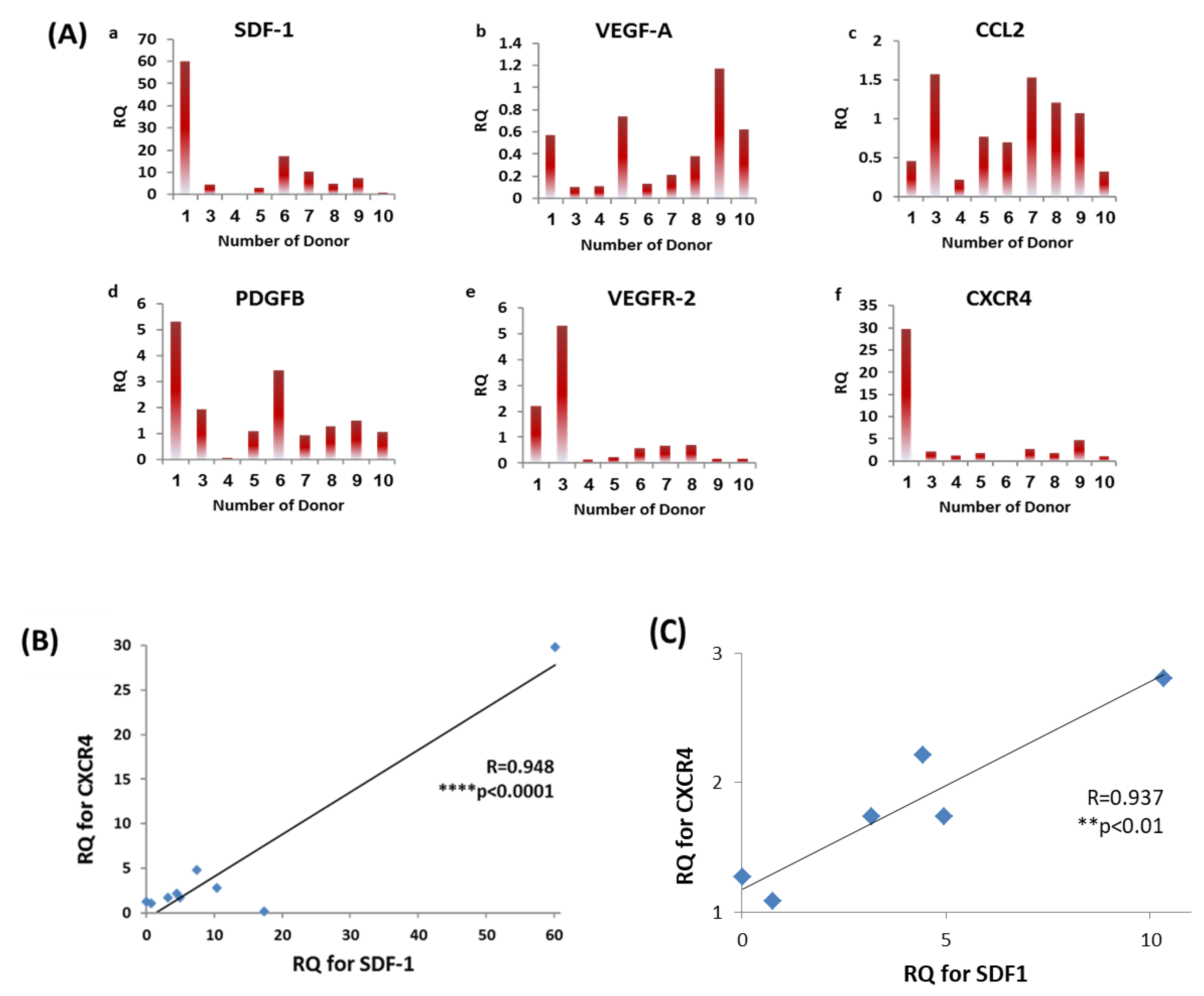
2.6. RNA Extraction and Real-Time PCR
2.7. Human EPCs Transplantation in Ectopic Subcutaneous Bone Model
2.8. Histological Preparation
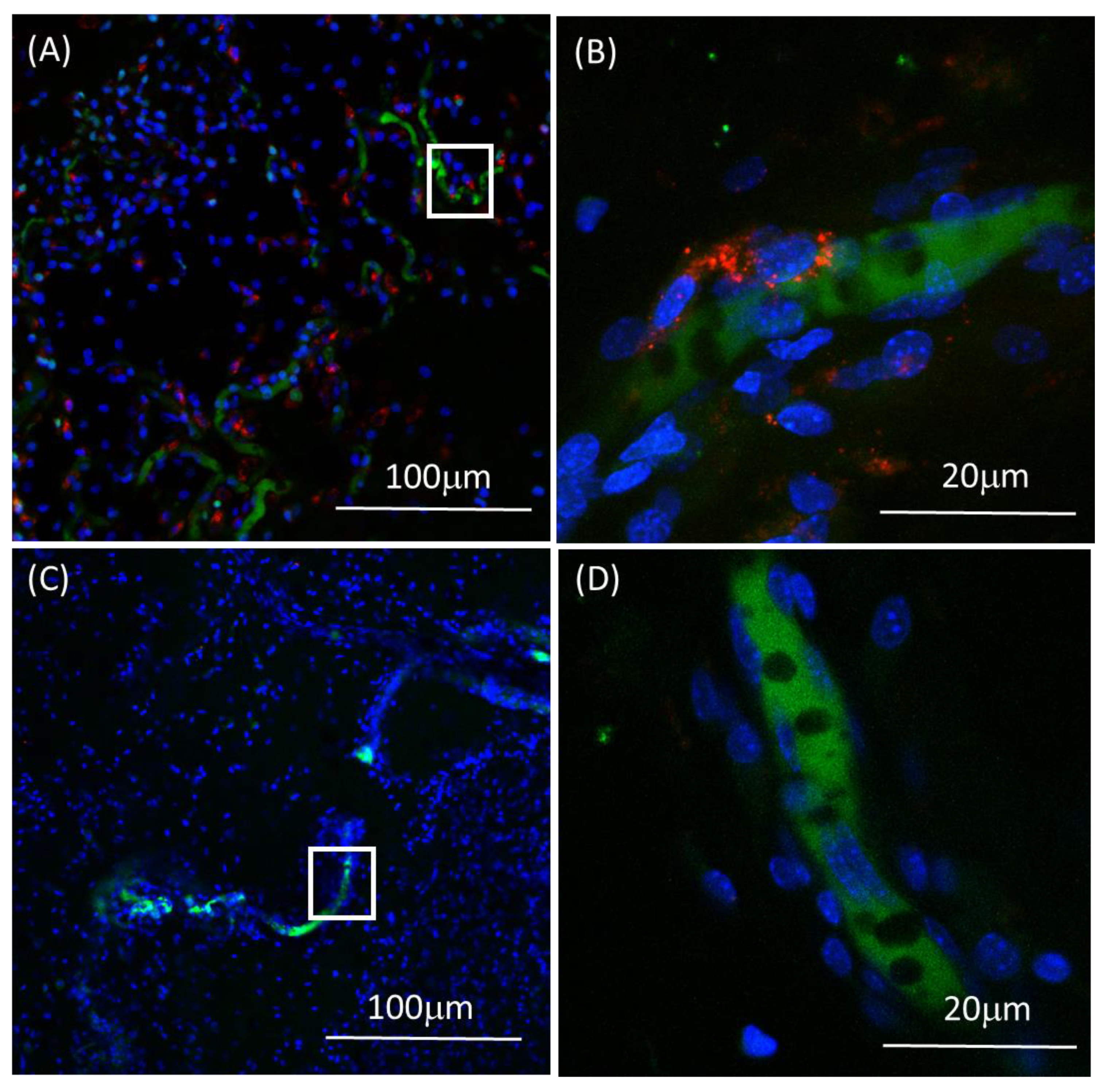
2.9. Immunohistochemistry
2.10. Statistical Analysis
3. Results
3.1. Donor Demographics
3.2. EPCs Expressed a High Percentage of CD31; CD34; VEGFR-2 and CXCR4
3.3. Diversity in EPCs In-Vitro Functions amongst Donors
3.4. Diversity in EPCs Genotype amongst Donors
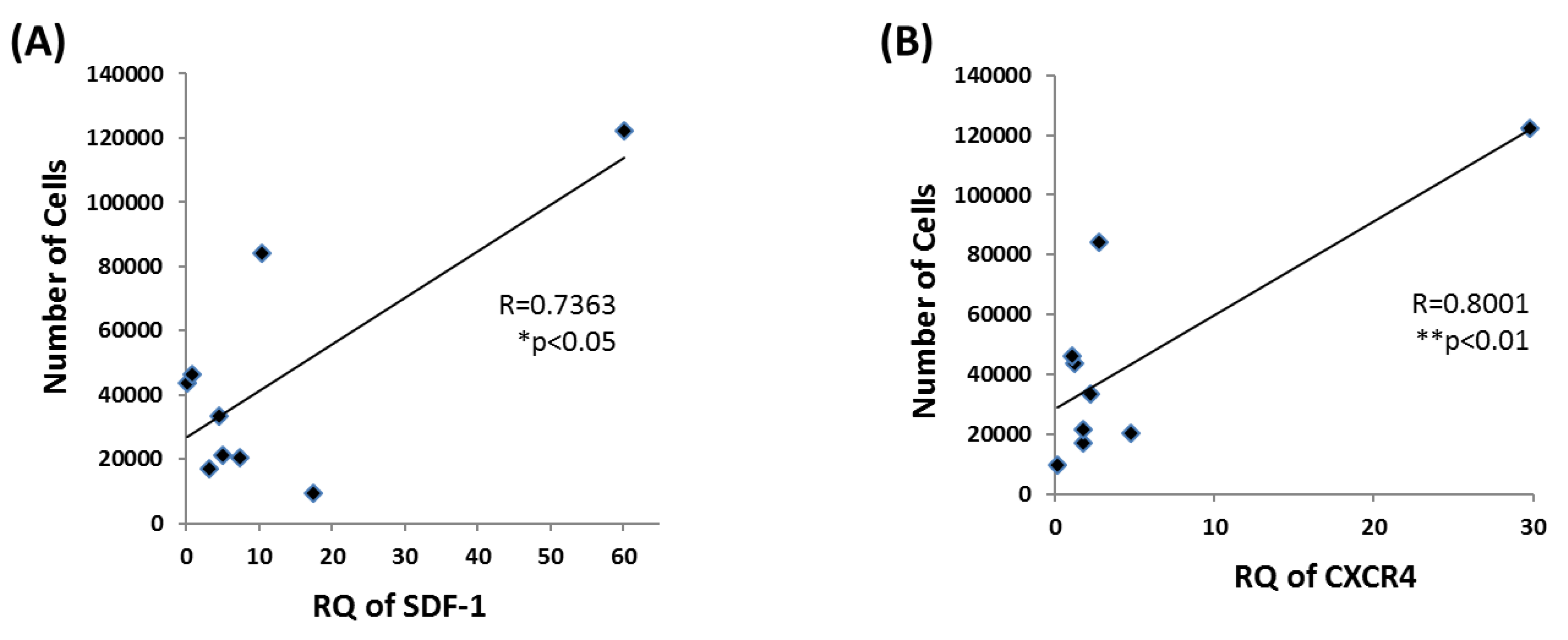
3.5. EPC Proliferation Correlates with SDF-1 and CXCR4 mRNA Levels
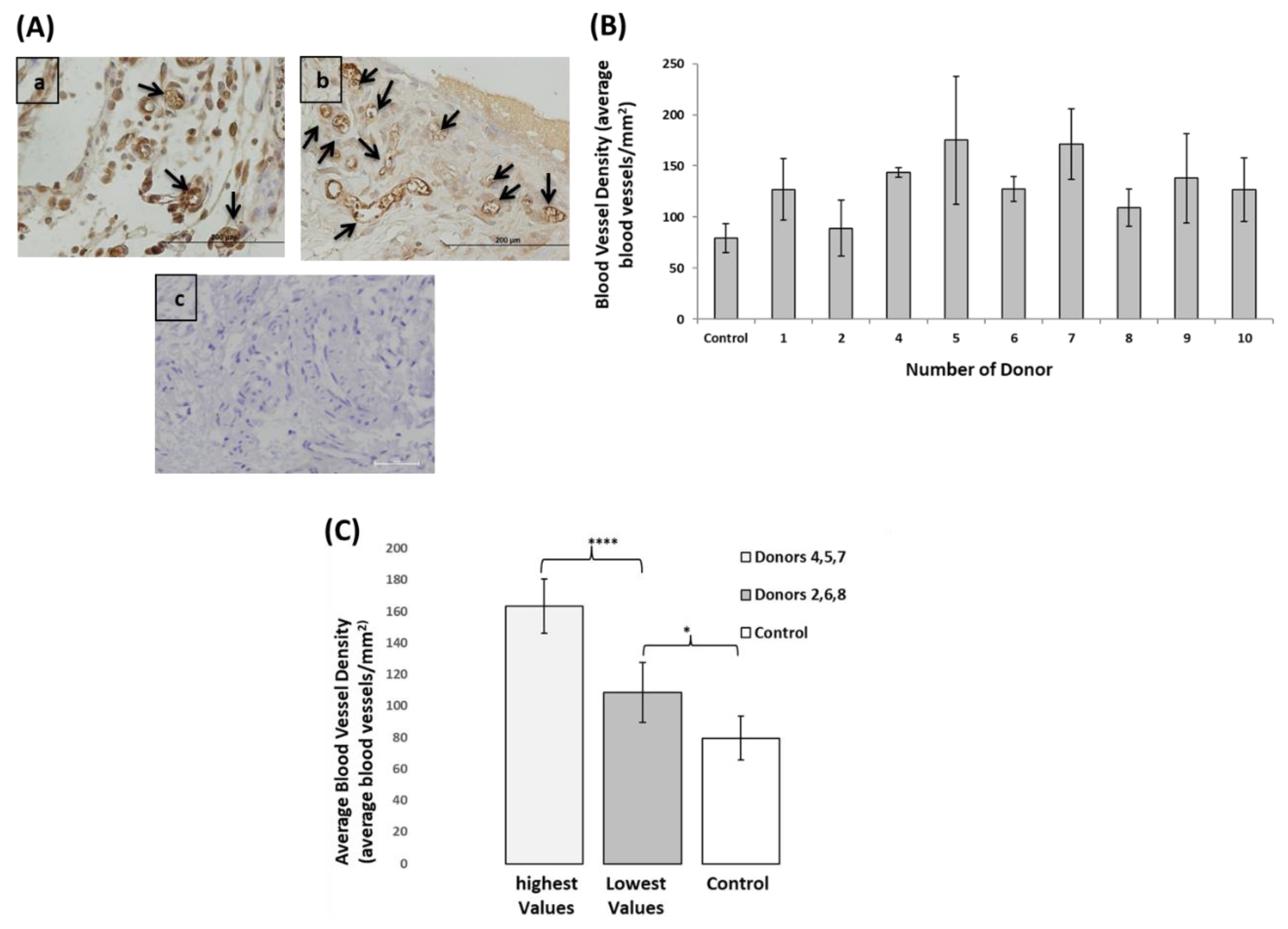
3.6. EPCs Angiogenic Capacity Varied between the Donors, However, even the Lowest Performing Donors Showed Higher Angiogenic Capacity Compared to Control
3.7. Prediction of In-Vivo Blood Vessel Formation Using a Regression Model Equation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Strauer, B.E.; Brehm, M.; Zeus, T.; Köstering, M.; Hernandez, A.; Sorg, R.V.; Kögler, G.; Wernet, P.; Köstering, M.; Sorg, R.V.; et al. Repair of Infarcted Myocardium by Autologous Intracoronary Mononuclear Bone Marrow Cell Transplantation in Humans. Circulation 2002, 106, 1913–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leistner, D.M.; Fischer-Rasokat, U.; Honold, J.; Seeger, F.H.; Schächinger, V.; Lehmann, R.; Martin, H.; Burck, I.; Urbich, C.; Dimmeler, S.; et al. Transplantation of progenitor cells and regeneration enhancement in acute myocardial infarction (TOPCARE-AMI): Final 5-year results suggest long-term safety and efficacy. Clin. Res. Cardiol. 2011, 100, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Stamm, C.; Westphal, B.; Kleine, H.-D.; Petzsch, M.; Kittner, C.; Klinge, H.; Schümichen, C.A.; Nienaber, C.; Freund, M.; Steinhoff, G. Autologous bone-marrow stem-cell transplantation for myocardial regeneration. Lancet 2003, 361, 45–46. [Google Scholar] [CrossRef]
- Wollert, K.C.; Meyer, G.P.; Lotz, J.; Ringes Lichtenberg, S.; Lippolt, P.; Breidenbach, C.; Fichtner, S.; Korte, T.; Hornig, B.; Messinger, D.; et al. Intracoronary autolo-gous bone-marrow cell transfer after myocardial infarction: The BOOST random-ised controlled clinical trial. Lancet 2004, 364, 141–148. [Google Scholar] [CrossRef]
- Kang, H.-J.; Kim, H.-S.; Zhang, S.-Y.; Park, K.-W.; Cho, H.-J.; Koo, B.-K.; Kim, Y.-J.; Lee, D.S.; Sohn, D.-W.; Han, K.-S.; et al. Effects of intracoronary infusion of peripheral blood stem-cells mobilised with granulocyte-colony stimulating factor on left ventricular systolic function and restenosis after coronary stenting in myocardi-al infarction: The MAGIC cell randomised clinical trial. Lancet 2004, 363, 751–756. [Google Scholar] [PubMed]
- Fang, J.; Guo, Y.; Tan, S.; Li, Z.; Xie, H.; Chen, P.; Wang, K.; He, Z.; He, P.; Ke, Y.; et al. Autologous Endothelial Progenitor Cells Transplantation for Acute Ischemic Stroke: A 4-Year Follow-Up Study. STEM CELLS Transl. Med. 2018, 8, 14–21. [Google Scholar] [CrossRef]
- Lopes, L.; Setia, O.; Aurshina, A.; Liu, S.; Hu, H.; Isaji, T.; Liu, H.; Wang, T.; Ono, S.; Guo, X.; et al. Stem cell therapy for diabetic foot ulcers: A review of preclinical and clinical research. Stem Cell Res. Ther. 2018, 9, 188. [Google Scholar] [CrossRef]
- Xie, B.; Luo, H.; Zhang, Y.; Wang, Q.; Zhou, C.; Xu, D. Autologous Stem Cell Thera-py in Critical Limb Ischemia: A Meta-Analysis of Randomized Controlled Trials. Stem Cells Int. 2018, 2018. [Google Scholar] [CrossRef]
- D’Avola, D.; Fernandez-Ruiz, V.; Carmona-Torre, F.; Méndez, M.; Pèrez-Calvo, J.; Prosper, F.; Andreu, E.J.; Herrero, J.I.; Iñarrairaegui, M.; Fuertes, C.; et al. Phase 1–2 pilot clinical trial in patients with decompensated liver cirrhosis treated with bone marrow–derived endothelial progenitor cells. Transl. Res. 2017, 188, 80–91. [Google Scholar] [CrossRef]
- Rozen, N.; Bick, T.; Bajayo, A.; Shamian, B.; Schrift-Tzadok, M.; Gabet, Y.; Yayon, A.; Bab, I.; Soudry, M.; Lewinson, D. Transplanted blood-derived endothe-lial progenitor cells (EPC) enhance bridging of sheep tibia critical size defects. Bone 2009, 45, 918–924. [Google Scholar] [CrossRef]
- Loomans, C.J.M.; de Koning, E.J.P.; Staal, F.J.T.; Rookmaaker, M.B.; Verseyden, C.; de Boer, H.C.; Verhaar, M.C.; Braam, B.; Rabelink, T.J.; van Zonneveld, A.-J. En-dothelial progenitor cell dysfunction: A novel concept in the pathogenesis of vascular complications of type 1 diabetes. Diabetes 2004, 53, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Tepper, O.M.; Galiano, R.D.; Capla, J.M.; Kalka, C.; Gagne, P.J.; Jacobowitz, G.R.; Levine, J.P.; Gurtner, G.C. Human endothelial progenitor cells from type II diabet-ics exhibit impaired proliferation, adhesion, and incorporation into vascular structures. Circulation 2002, 106, 2781–2786. [Google Scholar] [CrossRef] [PubMed]
- Lyden, D.; Hattori, K.; Dias, S.; Costa, C.; Blaikie, P.; Butros, L.; Chadburn, A.; Heissig, B.; Marks, W.; Witte, L.; et al. Impaired recruitment of bone-marrow-derived endothelial and hematopoietic precursor cells blocks tumor angiogene-sis and growth. Nat. Med. 2001, 7, 1194–1201. [Google Scholar] [CrossRef] [PubMed]
- Shaked, Y.; Ciarrocchi, A.; Franco, M.; Lee, C.R.; Man, S.; Cheung, A.M.; Hicklin, D.J.; Chaplin, D.; Foster, F.S.; Benezra, R.; et al. Therapy-induced acute recruit-ment of circulating endothelial progenitor cells to tumors. Science 2006, 313, 1785–1787. [Google Scholar] [CrossRef]
- Stoll, B.R.; Migliorini, C.; Kadambi, A.; Munn, L.L.; Jain, R.K. A mathematical model of the contribution of endothelial progenitor cells to angiogenesis in tu-mors: Implications for antiangiogenic therapy. Blood 2003, 102, 2555–2561. [Google Scholar] [CrossRef]
- Kondo, T.; Hayashi, M.; Takeshita, K.; Numaguchi, Y.; Kobayashi, K.; Iino, S.; Inden, Y.; Murohara, T. Smoking cessation rapidly increases circulating progeni-tor cells in peripheral blood in chronic smokers. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1442–1447. [Google Scholar] [CrossRef]
- Mobarrez, F.; Antoniewicz, L.; Bosson, J.A.; Kuhl, J.; Pisetsky, D.S.; Lundbäck, M. The Effects of Smoking on Levels of Endothelial Progenitor Cells and Micro-particles in the Blood of Healthy Volunteers. PLoS ONE 2014, 9, e90314. [Google Scholar] [CrossRef]
- Lui, M.M.-S.; Tse, H.-F.; Mak, J.C.-W.; Lam, J.C.-M.; Lam, D.C.-L.; Tan, K.C.B.; Ip, M.S.-M. Altered profile of circulating endothelial progenitor cells in obstructive sleep apnea. Sleep Breath Schlaf Atm. 2013, 17, 937–942. [Google Scholar] [CrossRef]
- Duijvestein, M.; Vos, A.C.W.; Roelofs, H.; Wildenberg, M.E.; Wendrich, B.B.; Verspaget, H.W.; Kooy-Winkelaar, E.M.C.; Koning, F.; Zwaginga, J.J.; Fidder, H.H.; et al. Autologous bone marrow-derived mesenchymal stromal cell treatment for refractory luminal Crohn’s disease: Results of a phase I study. Gut 2010, 59, 1662–1669. [Google Scholar] [CrossRef]
- Zigdon-Giladi, H.; Elimelech, R.; Michaeli-Geller, G.; Rudich, U.; Machtei, E.E. Safety profile and long-term engraftment of human CD31 + blood progenitors in bone tissue engineering. Cytotherapy 2017, 19, 895–908. [Google Scholar] [CrossRef]
- Huyck, L.; Ampe, C.; Van Troys, M. The XTT Cell Proliferation Assay Applied to Cell Layers Embedded in Three-Dimensional Matrix. Assay Drug Dev. Technol. 2012, 10, 382–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, G.E.; Chen, E. Life stress and diminished expression of genes encoding glucocorticoid receptor and beta2-adrenergic receptor in children with asthma. Proc. Natl. Acad. Sci. USA 2006, 103, 5496–5501. [Google Scholar] [CrossRef] [PubMed]
- Okubo, Y.; Siddle, K.; Firth, H.; O’Rahilly, S.; Wilson, L.C.; Willatt, L.; Fukushima, T.; Takahashi, S.-I.; Petry, C.J.; Saukkonen, T.; et al. Cell Proliferation Activities on Skin Fibroblasts from a Short Child with Absence of One Copy of the Type 1 In-sulin-Like Growth Factor Receptor (IGF1R) Gene and a Tall Child with Three Copies of the IGF1R Gene. J. Clin. Endocrinol. Metab. 2003, 88, 5981–5988. [Google Scholar] [CrossRef] [PubMed]
- Kalka, C.; Masuda, H.; Takahashi, T.; Kalka-Moll, W.M.; Silver, M.; Kearney, M.; Li, T.; Isner, J.M.; Asahara, T. Transplantation of ex vivo expanded endothelial progenitor cells for therapeutic neovascularization. Proc. Natl. Acad. Sci. 2000, 97, 3422–3427. [Google Scholar] [CrossRef]
- Asahara, T.; Kawamoto, A. Endothelial progenitor cells for postnatal vasculo-genesis. Am. J. Physiol. Cell Physiol. 2004, 287, C572–C579. [Google Scholar] [CrossRef]
- Peripheral Blood-Derived Endothelial Progenitor Cells Enhance Vertical Bone Formation-Zigdon-Giladi-2015-Clinical Implant Dentistry and Related Research-Wiley Online Library. Available online: https://onlinelibrary-wiley-com.ezlibrary.technion.ac.il/doi/full/10.1111/cid.12078 (accessed on 15 March 2019).
- Machtei, E.E.; Kim, D.M.; Karimbux, N.; Zigdon-Giladi, H. The use of endothelial progenitor cells combined with barrier membrane for the reconstruction of pe-ri-implant osseous defects: An animal experimental study. J. Clin. Periodontol. 2016, 43, 289–297. [Google Scholar] [CrossRef]
- Kaur, I.; Rawal, P.; Rohilla, S.; Bhat, M.H.; Sharma, P.; Siddiqui, H.; Kaur, S. En-dothelial progenitor cells from aged subjects display decreased expression of sirtuin 1, angiogenic functions, and increased senescence. Cell Biol. Int. 2018, 42, 1212–1220. [Google Scholar] [CrossRef]
- Dauwe, D.; Pelacho, B.; Wibowo, A.; Walravens, A.-S.; Verdonck, K.; Gillijns, H.; Caluwe, E.; Pokreisz, P.; van Gastel, N.; Carmeliet, G.; et al. Neovascularization Po-tential of Blood Outgrowth Endothelial Cells From Patients With Stable Ischemic Heart Failure Is Preserved. J. Am. Heart Assoc. 2016, 5, e002288. [Google Scholar] [CrossRef]
- Huizer, K.; Mustafa, D.A.M.; Spelt, J.C.; Kros, J.M.; Sacchetti, A. Improving the characterization of endothelial progenitor cell subsets by an optimized FACS protocol. PLoS ONE 2017, 12, e0184895. [Google Scholar] [CrossRef]
- Asahara, T.; Murohara, T.; Sullivan, A.; Silver, M.; van der Zee, R.; Li, T.; Witzenbichler, B.; Schatteman, G.; Isner, J.M. Isolation of Putative Progenitor En-dothelial Cells for Angiogenesis. Science 1997, 275, 964–966. [Google Scholar] [CrossRef]
- Dowling, M.-A.; Smith, M.; Melero-Martin, J.M.; Al-Rubeai, M.; Melero-Martin, J.M.; Al-Rubeai, M. Optimal in-vitro expansion of chondroprogenitor cells in monolayer culture. Biotechnol Bioeng. 2006, 93, 519–533. [Google Scholar]
- Yoder, M.C.; Mead, L.E.; Prater, D.; Krier, T.R.; Mroueh, K.N.; Li, F.; Krasich, R.; Temm, C.J.; Prchal, J.T.; Ingram, D.A. Redefining endothelial progenitor cells via clonal analysis and hematopoietic stem/progenitor cell principals. Blood 2007, 109, 1801–1809. [Google Scholar] [CrossRef] [PubMed]
- Yoon, C.-H.; Hur, J.; Park, K.-W.; Kim, J.-H.; Lee, C.-S.; Oh, I.-Y.; Kim, T.-Y.; Cho, H.-J.; Kang, H.-J.; Chae, I.-H.; et al. Synergistic Neovascularization by Mixed Trans-plantation of Early Endothelial Progenitor Cells and Late Outgrowth Endothelial Cells. Circulation 2005, 112, 1618–1627. [Google Scholar] [CrossRef] [PubMed]
- Janicki, P.; Boeuf, S.; Steck, E.; Egermann, M.; Kasten, P.; Richter, W. Prediction of in vivo bone forming potency of bone marrow-derived human mesenchymal stem cells. Eur. Cell. Mater. 2011, 21, 488–577. [Google Scholar] [CrossRef] [PubMed]
- Deskins, D.L.; Bastakoty, D.; Saraswati, S.; Shinar, A.; Holt, G.E.; Young, P.P. Human Mesenchymal Stromal Cells: Identifying Assays to Predict Potency for Therapeutic Selection. STEM CELLS Transl. Med. 2013, 2, 151–158. [Google Scholar] [CrossRef]
- Jaffe, E.A.; Nachman, R.L.; Becker, C.G.; Minick, C.R. Culture of Human Endo-thelial Cells Derived from Umbilical Veins.Identification by morphologic and im-munologic criteria. J. Clin. Invest. 1973, 52, 2745–2756. [Google Scholar] [CrossRef]
- Xu, C.; Rosler, E.; Jiang, J.; Lebkowski, J.S.; Gold, J.D.; O’Sullivan, C.; Mok, M.; Bronstein, A.; Carpenter, M.K.; Delavan-Boorsma, K.; et al. Basic Fibroblast Growth Factor Supports Undifferentiated Human Embryonic Stem Cell Growth Without Conditioned Medium. STEM CELLS 2005, 23, 315–323. [Google Scholar] [CrossRef]
- Liang, T.; Zhu, L.; Gao, W.; Gong, M.; Ren, J.; Yao, H.; Wang, K.; Shi, D. Coculture of endothelial progenitor cells and mesenchymal stem cells enhanced their pro-liferation and angiogenesis through PDGF and Notch signaling. FEBS Open Bio 2017, 7, 1722–1736. [Google Scholar] [CrossRef]
- Loibl, M.; Binder, A.; Herrmann, M.; Duttenhoefer, F.; Richards, R.G.; Nerlich, M.; Alini, M.; Verrier, S. Direct Cell-Cell Contact between Mesenchymal Stem Cells and Endothelial Progenitor Cells Induces a Pericyte-Like Phenotype In Vitro. Available online: https://www.hindawi.com/journals/bmri/2014/395781/ (accessed on 12 July 2019).
- Burlacu, A.; Grigorescu, G.; Rosca, A.-M.; Preda, M.B.; Simionescu, M. Factors Secreted by Mesenchymal Stem Cells and Endothelial Progenitor Cells Have Complementary Effects on Angiogenesis In Vitro. Stem Cells Dev. 2012, 22, 643–653. [Google Scholar] [CrossRef]
- Mendicino, M.; Bailey, A.M.; Wonnacott, K.; Puri, R.K.; Bauer, S.R. MSC-Based Product Characterization for Clinical Trials: An FDA Perspective. Cell Stem Cell 2014, 14, 141–145. [Google Scholar] [CrossRef] [Green Version]
- Chinnadurai, R.; Rajan, D.; Qayed, M.; Arafat, D.; Garcia, M.; Liu, Y.; Kuga-thasan, S.; Anderson, L.J.; Gibson, G.; Galipeau, J. Potency Analysis of Mesenchy-mal Stromal Cells Using a Combinatorial Assay Matrix Approach. Cell Rep. 2018, 22, 2504–2517. [Google Scholar] [CrossRef] [PubMed]
- Lataillade, J.-J.; Clay, D.; Bourin, P.; Hérodin, F.; Dupuy, C.; Jasmin, C.; Le Bousse-Kerdilès, M.-C. Stromal cell-derived factor 1 regulates primitive hemato-poiesis by suppressing apoptosis and by promoting G(0)/G(1) transition in CD34(+) cells: Evidence for an autocrine/paracrine mechanism. Blood 2002, 99, 1117–1129. [Google Scholar] [CrossRef] [PubMed]
- Rafii, S.; Heissig, B.; Hattori, K. Efficient mobilization and recruitment of mar-row-derived endothelial and hematopoietic stem cells by adenoviral vectors ex-pressing angiogenic factors. Gene Ther. 2002, 9, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.A.; Hattori, K.; Heissig, B.; Shieh, J.H.; Dias, S.; Crystal, R.G.; Rafii, S. Mobilization of endothelial and hematopoietic stem and progenitor cells by ad-enovector-mediated elevation of serum levels of SDF-1, VEGF, and angiopoietin-1. Ann. N. Y. Acad. Sci. 2001, 938, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Tan, Y.; Cai, S.; Xiong, X.; Wang, L.; Ye, Q.; Yan, X.; Ma, K.; Cai, L. The role of CXCR7 on the adhesion, proliferation and angiogenesis of endothelial progen-itor cells. J. Cell. Mol. Med. 2011, 15, 1299–1309. [Google Scholar] [CrossRef] [PubMed]
- Heiss, C.; Keymel, S.; Niesler, U.; Ziemann, J.; Kelm, M.; Kalka, C. Impaired Pro-genitor Cell Activity in Age-Related Endothelial Dysfunction. J. Am. Coll. Cardiol. 2005, 45, 1441–1448. [Google Scholar] [CrossRef]
- Scheubel, R.J.; Zorn, H.; Silber, R.-E.; Kuss, O.; Morawietz, H.; Holtz, J.; Simm, A. Age-dependent depression in circulating endothelial progenitor cells inpatients undergoing coronary artery bypass grafting. J. Am. Coll. Cardiol. 2003, 42, 2073–2080. [Google Scholar] [CrossRef] [Green Version]
- Henrich, D.; Verboket, R.; Schaible, A.; Kontradowitz, K.; Oppermann, E.; Brune, J.C.; Nau, C.; Meier, S.; Bonig, H.; Marzi, I.; et al. Characterization of Bone Marrow Mononuclear Cells on Biomaterials for Bone Tissue Engineering In Vitro. BioMed Res. Int. 2015, 2015, 1–12. [Google Scholar]
- Levenberg, S.; Huang, N.F.; Lavik, E.; Rogers, A.B.; Itskovitz-Eldor, J.; Langer, R. Differentiation of human embryonic stem cells on three-dimensional polymer scaffolds. Proc. Natl. Acad. Sci. USA 2003, 100, 12741–12746. [Google Scholar] [CrossRef] [Green Version]
- Levenberg, S.; Golub, J.S.; Amit, M.; Itskovitz-Eldor, J.; Langer, R. Endothelial cells derived from human embryonic stem cells. Proc. Natl. Acad. Sci. USA 2002, 99, 4391–4396. [Google Scholar] [CrossRef] [Green Version]
- Horwitz, E.M.; Gordon, P.L.; Koo, W.K.K.; Marx, J.C.; Neel, M.D.; McNall, R.Y.; Muul, L.; Hofmann, T. Isolated allogeneic bone marrow-derived mesenchymal cells engraft and stimulate growth in children with osteogenesis imperfecta: Im-plications for cell therapy of bone. Proc. Natl. Acad. Sci. USA 2002, 99, 8932–8937. [Google Scholar] [CrossRef] [PubMed]
- Ball, L.M.; Bernardo, M.E.; Roelofs, H.; van Tol, M.J.D.; Contoli, B.; Zwaginga, J.J.; Avanzini, M.A.; Conforti, A.; Bertaina, A.; Giorgiani, G.; et al. Multiple infusions of mesenchymal stromal cells induce sustained remission in children with ster-oid-refractory, grade III-IV acute graft-versus-host disease. Br. J. Haematol. 2013, 163, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, F.; Rossant, J.; Yamaguchi, T.P.; Gertsenstein, M.; Wu, X.-F.; Breitman, M.L.; Schuh, A.C. Failure of blood-island formation and vasculogenesis in Flk-1-deficient mice. Nature 1995, 376, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Kanno, S.; Oda, N.; Abe, M.; Terai, Y.; Ito, M.; Shitara, K.; Tabayashi, K.; Shibuya, M.; Sato, Y. Roles of two VEGF receptors, Flt-1 and KDR, in the signal transduc-tion of VEGF effects in human vascular endothelial cells. Oncogene 2000, 19, 2138–2146. [Google Scholar] [CrossRef] [PubMed]
- Millauer, B. High affinity VEGF binding and developmental expression suggest Flk-1 as a major regulator of vasculogenesis and angiogenesis. Cell 1993, 72, 835–846. [Google Scholar] [CrossRef]
- Aguirre, A.; Planell, J.A.; Engel, E. Dynamics of bone marrow-derived endothe-lial progenitor cell/mesenchymal stem cell interaction in co-culture and its im-plications in angiogenesis. Biochem. Biophys. Res. Commun. 2010, 400, 284–291. [Google Scholar] [CrossRef]
- Watt, S.M.; Athanassopoulos, A.; Harris, A.L.; Tsaknakis, G. Human endothelial stem/progenitor cells, angiogenic factors and vascular repair. J. R. Soc. Interface 2010, 7, S731–751. [Google Scholar] [CrossRef]
- Samsonraj, R.M.; Rai, B.; Sathiyanathan, P.; Puan, K.J.; Rötzschke, O.; Hui, J.H.; Raghunath, M.; Stanton, L.W.; Nurcombe, V.; Cool, S.M. Establishing Criteria for Human Mesenchymal Stem Cell Potency. Stem Cells 2015, 33, 1878–1891. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Cloud, C.; Duke, T.; Owczarski, S.; Mehrotra, S.; Adams, D.B.; Morgan, K.; Gilkeson, G.; Wang, H. Mesenchymal Stem Cells from Chronic Pancreatitis Patients Show Comparable Potency Compared to Cells from Healthy Donors. Stem Cells Transl. Med. 2019, 8, 418–429. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Donor | Positive Stained EPCs (%) | |||||
|---|---|---|---|---|---|---|
| CD14 | CD31 | CD34 | CD45 | VEGFR-2/KDR | CXCR4 | |
| 1 | 1.9 | 98.8 | 98.9 | 18.1 | 69.2 | 99.7 |
| 2 | 5.1 | 93.4 | 83.6 | 6.2 | 99.1 | 99.5 |
| 3 | 0 | 99.2 | 93.0 | 3.8 | 96.5 | 98.3 |
| 4 | 1.8 | 100 | 55.4 | 1.9 | 98.7 | 98.0 |
| 5 | 4.8 | 99.6 | 84.7 | 14.3 | 22.8 | 97.3 |
| 6 | 1.9 | 93.8 | 96.2 | 5.9 | 8.9 | 2.8 |
| 7 | 0.6 | 100 | 21.5 | 3.1 | 63.3 | 67.3 |
| 8 | 5.7 | 91.7 | 76.9 | 10.5 | 15.8 | 18.4 |
| 9 | 17.0 | 99.4 | 98.8 | 20.6 | 99.0 | 90.4 |
| 10 | 3.7 | 99.7 | 94.1 | 14.4 | 100 | 96.2 |
| Average ± SD | 4.1 ± 4.7 | 97.7 ± 3.1 | 81.2 ± 23.5 | 10.4 ± 6.5 | 69.5 ± 36.7 | 78.9 ± 35.2 |
| Donor | Migration (B in Equation) | RT-PCR VEGFR-2 (C in Equation) | Calculated Blood Vessels (A in Equation) | In-Vivo Vessel Count |
|---|---|---|---|---|
| 5 | 2.94 | 0.23 | 173.96 | 175.0 |
| 7 | 2.32 | 0.66 | 171.10 | 171.2 |
| 10 | 2.30 | 0.17 | 131.98 | 126.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabbah, N.; Tamari, T.; Elimelech, R.; Doppelt, O.; Rudich, U.; Zigdon-Giladi, H. Predicting Angiogenesis by Endothelial Progenitor Cells Relying on In-Vitro Function Assays and VEGFR-2 Expression Levels. Biomolecules 2019, 9, 717. https://doi.org/10.3390/biom9110717
Sabbah N, Tamari T, Elimelech R, Doppelt O, Rudich U, Zigdon-Giladi H. Predicting Angiogenesis by Endothelial Progenitor Cells Relying on In-Vitro Function Assays and VEGFR-2 Expression Levels. Biomolecules. 2019; 9(11):717. https://doi.org/10.3390/biom9110717
Chicago/Turabian StyleSabbah, Nadin, Tal Tamari, Rina Elimelech, Ofri Doppelt, Utai Rudich, and Hadar Zigdon-Giladi. 2019. "Predicting Angiogenesis by Endothelial Progenitor Cells Relying on In-Vitro Function Assays and VEGFR-2 Expression Levels" Biomolecules 9, no. 11: 717. https://doi.org/10.3390/biom9110717
APA StyleSabbah, N., Tamari, T., Elimelech, R., Doppelt, O., Rudich, U., & Zigdon-Giladi, H. (2019). Predicting Angiogenesis by Endothelial Progenitor Cells Relying on In-Vitro Function Assays and VEGFR-2 Expression Levels. Biomolecules, 9(11), 717. https://doi.org/10.3390/biom9110717