
New Mutations in the 5′ Region of the Notch Gene Affect Drosophila melanogaster Oogenesis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
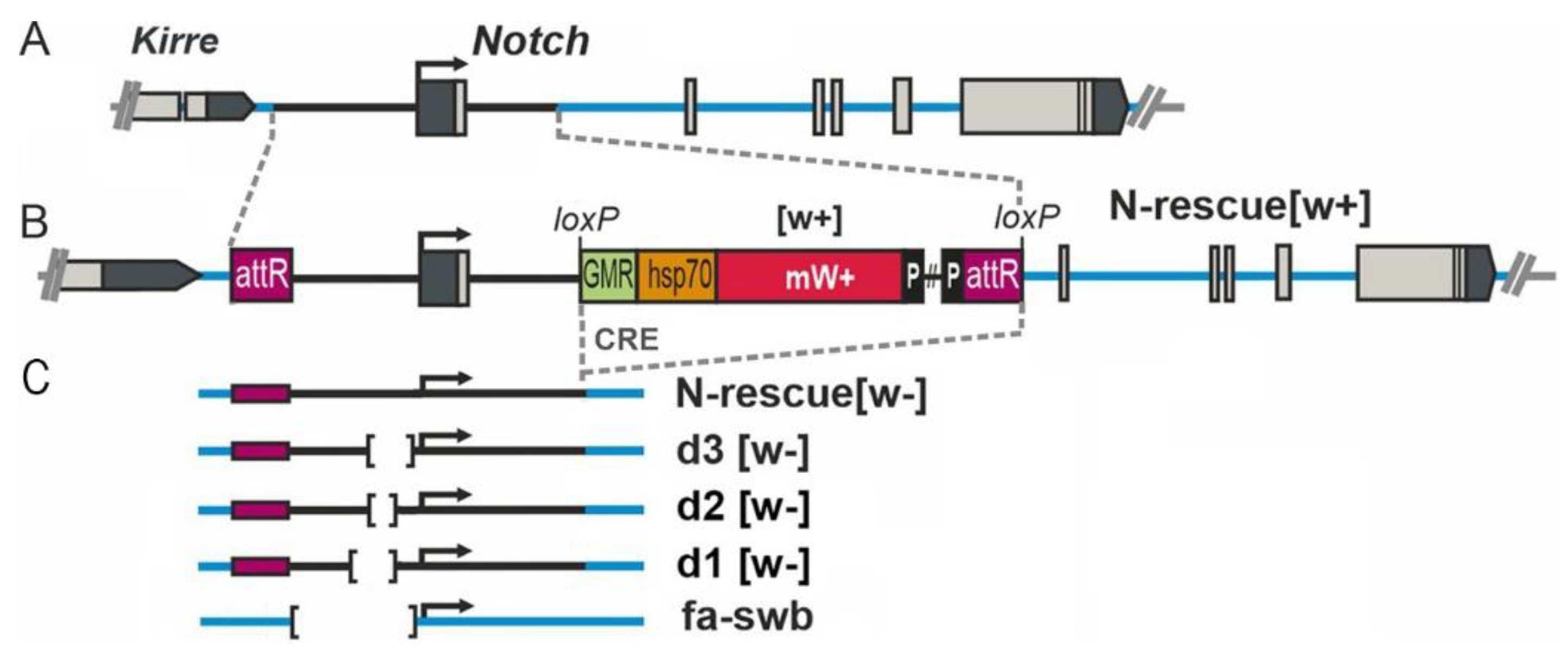
2.1. Drosophila Strains
2.2. Immunofluorescent Staining
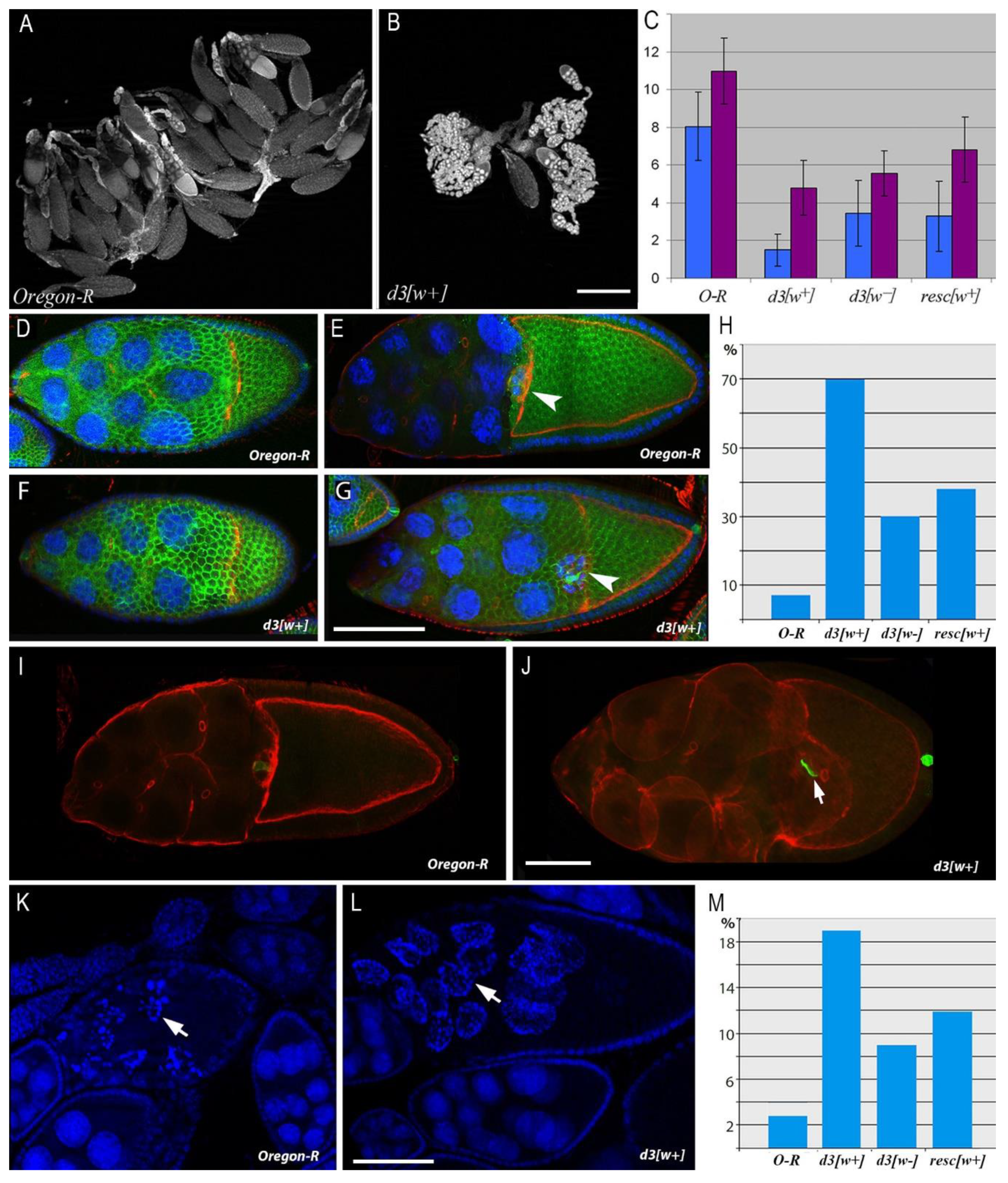
2.3. Ovarian Morphology Analysis and Egg Counting
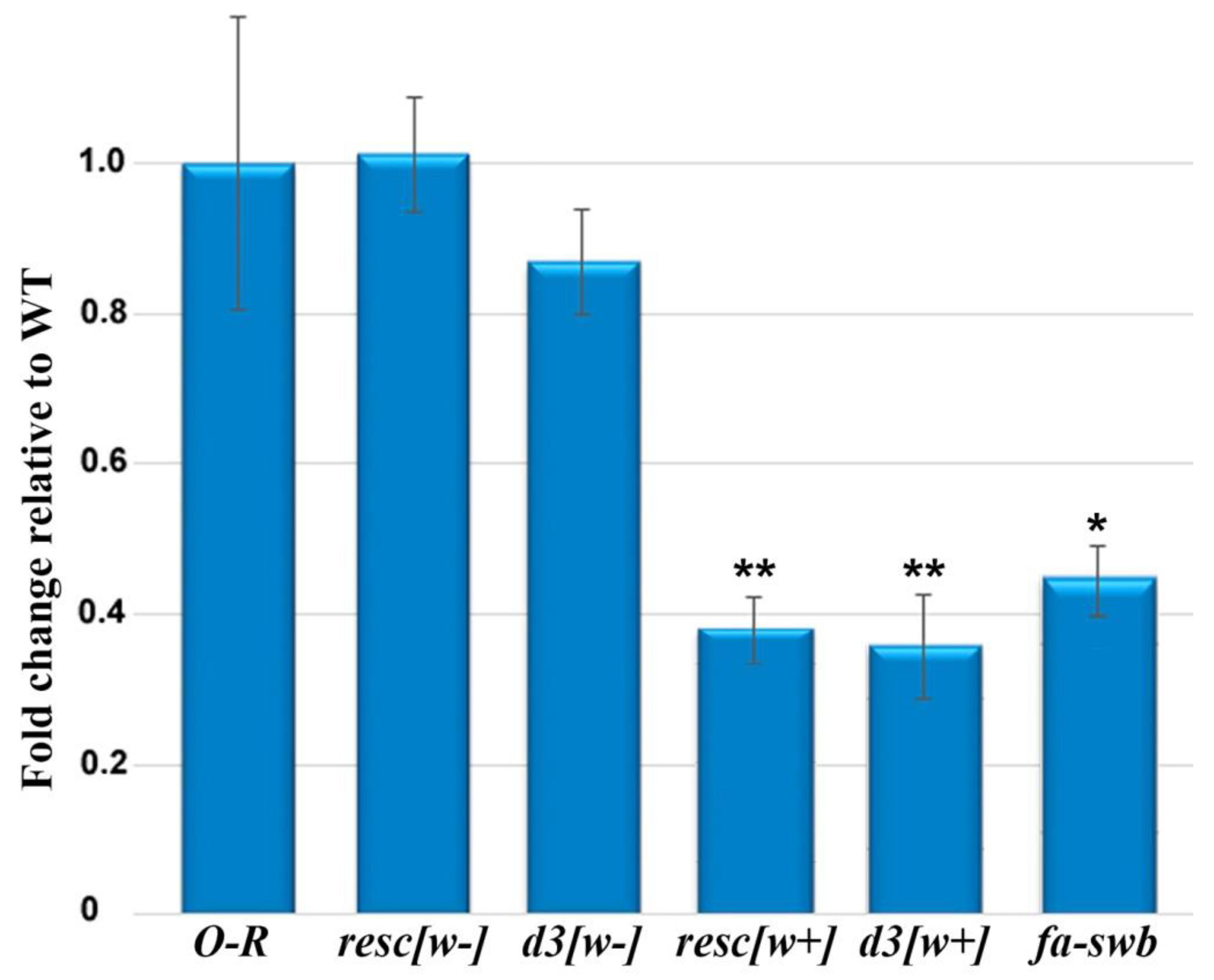
2.4. Reverse-Transcription Quantitative PCR (RT-qPCR)
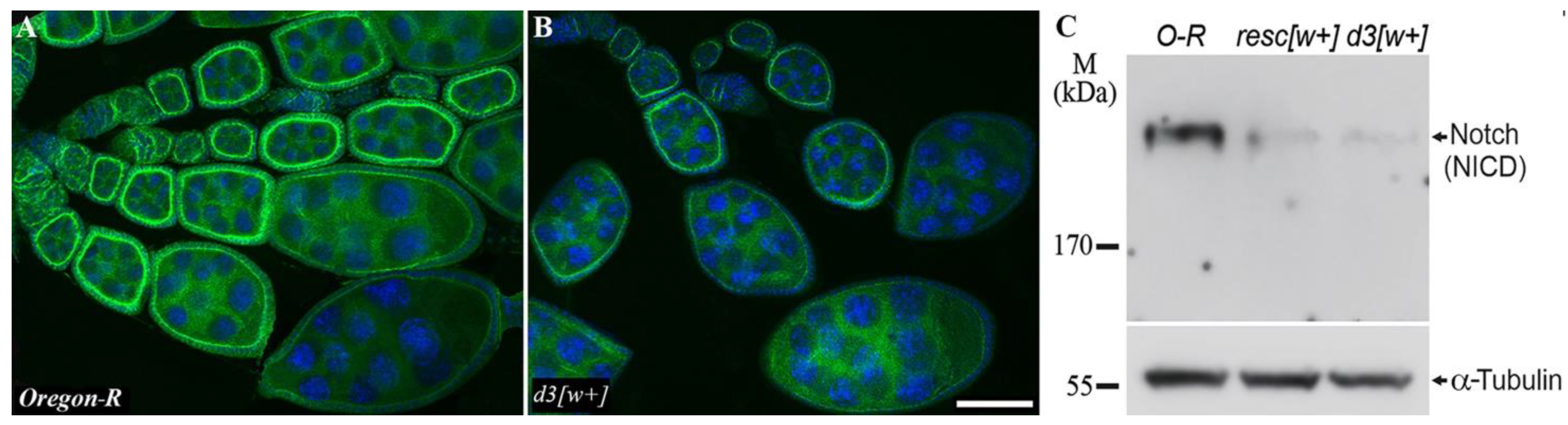
2.5. Western Blot
3. Results
3.1. Oogenesis Disturbances in Mutants with an Altered 5′ Regulatory Region of the Notch Gene
3.2. The Notch Expression Level in the Ovaries of the Mutant Females
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Fortini, M.E.; Bilder, D. Endocytic regulation of Notch signaling. Curr. Opin. Genet. Dev. 2009, 19, 323–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopan, R.; Ilagan, M.X. The canonical Notch signaling pathway: Unfolding the activation mechanism. Cell 2009, 137, 216–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tien, A.C.; Rajan, A.; Bellen, H.J. A Notch updated. J. Cell Biol. 2009, 184, 621–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artavanis-Tsakonas, S.; Rand, M.D.; Lake, R.J. Notch signaling: Cell fate control and signal integration in development. Science 1999, 284, 770–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portin, P. General outlines of the molecular genetics of the Notch signalling pathway in Drosophila melanogaster: A review. Hereditas 2002, 136, 89–96. [Google Scholar] [CrossRef]
- Grammont, M.; Irvine, K.D. fringe and Notch specify polar cell fate during Drosophila oogenesis. Development 2001, 128, 2243–2253. [Google Scholar] [CrossRef]
- Larkin, M.K.; Holder, K.; Yost, C.; Giniger, E.; Ruohola-Baker, H. Expression of constitutively active Notch arrests follicle cells at a precursor stage during Drosophila oogenesis and disrupts the anterior-posterior axis of the oocyte. Development 1996, 122, 3639–3650. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Caron, L.A.; Fehon, R.G.; Artavanis-Tsakonas, S. The involvement of the Notch locus in Drosophila oogenesis. Development 1992, 115, 913–922. [Google Scholar] [CrossRef]
- Dobens, L.; Jaeger, A.; Peterson, J.S.; Raftery, L.A. Bunched sets a boundary for Notch signaling to pattern anterior eggshell structures during Drosophila oogenesis. Dev. Biol. 2005, 287, 425–437. [Google Scholar] [CrossRef] [PubMed]
- López-Schier, H.; St.Johnston, D. Delta signaling from the germ line controls the proliferation and differentiation of the somatic follicle cells during Drosophila oogenesis. Genes Dev. 2001, 15, 1393–1405. [Google Scholar]
- Deng, W.M.; Althauser, C.; Ruohola-Baker, H. Notch-Delta signaling induces a transition from mitotic cell cycle to endocycle in Drosophila follicle cells. Development 2001, 128, 4737–4746. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Adam, J.C.; Montell, D. Spatially localized Kuzbanian required for specific activation of Notch during border cell migration. Dev. Biol. 2007, 301, 532–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schober, M.; Rebay, I.; Perrimon, N. Function of the ETS transcription factor Yan in border cell migration. Development 2005, 132, 3493–3504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Gridley, T. Notch Signaling during Oogenesis in Drosophila melanogaster. Genet. Res. Int. 2012, 2012, 648207. [Google Scholar] [CrossRef] [PubMed]
- Volkova, E.I.; Andreyenkova, N.G.; Andreyenkov, O.V.; Zhimulev, I.F.; Demakov, S.A. Structural and functional dissection of the 5′ region of the Notch gene in Drosophila melanogaster. Genes 2019, 10, 1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertceva, J.A.; Dorogova, N.V.; Bolobolova, E.U.; Nerusheva, O.O.; Fedorova, S.A.; Omelyanchuk, L.V. The role of Drosophila hyperplastic discs gene in spermatogenesis. Cell Biol. Int. 2010, 34, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Guild, G.M.; Connelly, P.S.; Shaw, M.K.; Tilney, L.G. Actin filament cables in Drosophila nurse cells are composed of modules that slide passively past one another during dumping. J. Cell Biol. 1997, 138, 783–797. [Google Scholar] [CrossRef] [Green Version]
- Maksimov, D.A.; Koryakov, D.E.; Belyakin, S.N. Developmental variation of the SUUR protein binding correlates with gene regulation and specific chromatin types in D. melanogaster. Chromosoma 2014, 123, 253–264. [Google Scholar] [CrossRef]
- Ramos, R.G.; Grimwade, B.G.; Wharton, K.A.; Scottgale, T.N.; Artavanis-Tsakonas, S. Physical and functional definition of the Drosophila Notch locus by P element transformation. Genetics 1989, 123, 337–348. [Google Scholar] [CrossRef]
- Assa-Kunik, E.; Torres, I.; Schejter, E.; Johnston, D.; Shilo, B. Drosophila follicle cells are patterned by multiple levels of Notch signaling and antagonism between the Notch and JAK/STAT pathways. Development 2007, 134, 1161–1169. [Google Scholar] [CrossRef] [Green Version]
- Markopoulou, K.; Welshons, W.J.; Artavanis-Tsakonas, S. Phenotypic and molecular analysis of the facets, a group of intronic mutations at the Notch locus of Drosophila melanogaster which affect postembryonic development. Genetics 1989, 122, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Qian, J. EnhancerAtlas 2.0: An updated resource with enhancer annotation in 586 tissue/cell types across nine species. Nucleic Acids Res. 2020, 48, D58–D64. [Google Scholar] [CrossRef] [Green Version]
- Goode, S.; Melnick, M.; Chou, T.B.; Perrimon, N. The neurogenic genes egghead and brainiac define a novel signaling pathway essential for epithelial morphogenesis during Drosophila oogenesis. Development 1996, 122, 3863–3879. [Google Scholar] [CrossRef] [PubMed]
- Ruohola, H.; Bremer, K.A.; Baker, D.; Swedlow, J.R.; Jan, L.Y.; Jan, Y.N. Role of neurogenic genes in establishment of follicle cell fate and oocyte polarity during oogenesis in Drosophila. Cell 1991, 66, 433–449. [Google Scholar] [CrossRef]
- Starble, R.; Pokrywka, N.J. The retromer subunit Vps26 mediates Notch signaling during Drosophila oogenesis. Mech. Dev. 2018, 149, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Torres, I.L.; López-Schier, H.; St Johnston, D. A Notch/Delta-dependent relay mechanism establishes anterior-posterior polarity in Drosophila. Dev. Cell 2003, 5, 547–558. [Google Scholar] [CrossRef] [Green Version]
- Grammont, M. Adherens junction remodeling by the Notch pathway in Drosophila melanogaster oogenesis. J. Cell Biol. 2007, 177, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Arzate-Mejía, R.G.; Cerecedo-Castillo, A.J.; Guerrero, G.; Furlan-Magaril, M.; Recillas-Targa, F. In situ dissection of domain boundaries affect genome topology and gene transcription in Drosophila. Nat. Commun 2020, 11, 894. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volkova, E.I.; Dorogova, N.V.; Andreyenkov, O.V.; Tikhomirov, S.A.; Demakov, S.A. New Mutations in the 5′ Region of the Notch Gene Affect Drosophila melanogaster Oogenesis. J. Dev. Biol. 2022, 10, 32. https://doi.org/10.3390/jdb10030032
Volkova EI, Dorogova NV, Andreyenkov OV, Tikhomirov SA, Demakov SA. New Mutations in the 5′ Region of the Notch Gene Affect Drosophila melanogaster Oogenesis. Journal of Developmental Biology. 2022; 10(3):32. https://doi.org/10.3390/jdb10030032
Chicago/Turabian StyleVolkova, Elena I., Natalya V. Dorogova, Oleg V. Andreyenkov, Saveliy A. Tikhomirov, and Sergey A. Demakov. 2022. "New Mutations in the 5′ Region of the Notch Gene Affect Drosophila melanogaster Oogenesis" Journal of Developmental Biology 10, no. 3: 32. https://doi.org/10.3390/jdb10030032
APA StyleVolkova, E. I., Dorogova, N. V., Andreyenkov, O. V., Tikhomirov, S. A., & Demakov, S. A. (2022). New Mutations in the 5′ Region of the Notch Gene Affect Drosophila melanogaster Oogenesis. Journal of Developmental Biology, 10(3), 32. https://doi.org/10.3390/jdb10030032